我国家禽新城疫病毒流行基因型分析
2022-04-11赵怡倩赵跃祺吴慧芳杨增岐刘海金
赵怡倩,赵跃祺,吴慧芳,杨增岐,刘海金
(西北农林科技大学动物医学院,陕西杨凌 712100)
新城疫(Newcastle disease,ND)是由副黏病毒科的新城疫病毒(Newcastle disease virus,NDV)强毒株引起的一种禽类烈性传染病,世界动物卫生组织(OIE)将其列为必须通报的动物疫病,我国将其列为一类动物疫病,给养禽业带来巨大经济损失[1]。NDV只有一种血清型,存在多种基因型,疫苗接种是防控ND的最有效方法,ND疫苗主要是由20世纪50年代分离的NDV弱毒株制成,包括VG/GA、Ulster/67、La Sota、B1、Clone 30和Mukteswar等,这些疫苗株属于Class Ⅱ类基因Ⅰ、Ⅱ或Ⅲ型[2]。现有的ND疫苗能保护鸡不被异源基因型NDV强毒株感染发病,但不能有效地防止病毒的复制和排泄[3-4]。家禽对NDV均易感。NDV有宿主嗜性,不同禽类中流行的NDV基因型不同。疫苗株和流行株基因型的不匹配是导致ND在免疫家禽中暴发的原因之一。迫切需要基于不同家禽的流行基因型来研发基因匹配的ND疫苗,从而提高疫苗防控ND的效果。
1 NDV基因型
NDV为单股负链RNA病毒,其基因组的长度为15186 bp、15192 bp或15198 bp,结构为3′Leader-NP-P-M-F-HN-L-Trailer′5,编码6个结构蛋白和2个非结构蛋白[1]。膜融合蛋白(fusion,F)是决定病毒毒力的主要因子。根据F基因,可把NDV分为Class Ⅰ类和Class Ⅱ类。Class Ⅰ类的NDV只有1个基因型,与Class Ⅱ类的NDV同源性约60%。Class Ⅱ类的NDV包含21个基因型,基因型的遗传距离相差约10%[5]。NDV在我国流行时间较长,本文从NCBI Nucleotide数据库中筛选了分离自我国的2 352株NDV,并对所有毒株的分离时间、宿主和基因分型进行统计分析[6](图1~图7)。我国家禽中存在复杂的NDV基因型,有Class Ⅰ类和Class Ⅱ类基因Ⅰ-Ⅲ、Ⅵ-Ⅸ、Ⅻ、ⅩⅢ、ⅩⅩ和ⅩⅩⅠ型。虽然家禽对Class Ⅰ类和Class Ⅱ类多种基因型均易感,但不同宿主中流行的NDV基因型不同。在鸡群中,Class Ⅱ类基因Ⅶ型(59.35%)是造成ND暴发的主要流行基因型,其中Ⅶ.1.1亚型(97.43%)占绝对优势(图2);水禽是NDV弱毒株的主要宿主,能够引起水禽严重发病的NDV强毒株主要为Class Ⅱ类基因Ⅶ型(28.36%),96.10%为Ⅶ.1.1亚型(图3);鸽ND主要由Class Ⅱ类基因Ⅵ型引起(78.04%)(图4)。
2 Class Ⅰ类NDV
20世纪70年代Class Ⅰ类NDV就已经在北美和欧亚大陆的家禽和野生鸟类中存在[7]。2002年我国在家鸭中首次发现了Class Ⅰ类NDV[8]。根据基因型分析,33.33%的我国水禽分离株为Class Ⅰ类弱毒株(图3),而且50.74%(205/404)Class Ⅰ类NDV从水禽和野生鸟类中分离出来。家鸭是Class Ⅰ类NDV的重要宿主,在病毒的保存和进化中发挥重要作用。Class Ⅰ类NDV对家禽无致病性。鸭源Class Ⅰ类NDV与常用NDV的La Sota、V4等疫苗株存在显著的抗原差异[9]。鸭源Class Ⅰ类NDV弱毒株在鸡的气囊内连续传代后毒性增强,在鸡组织器官内能够快速复制和扩散[10]。在我国,鸡经常与携带NDV的健康水禽接触甚至混养,给NDV弱毒株突变为强毒株创造了条件,可能导致Class Ⅰ类NDV引起的ND疫情。因此,应根据我国鸭源Class Ⅰ类NDV的特点,开发用于鸭类的疫苗,对家鸭提供保护的同时减少病毒的复制和排泄,从而避免鸭源Class Ⅰ NDV传播给鸡,造成毒力返强引起的ND疫情,以保护鸡群免受Class Ⅰ类NDV的潜在威胁。

图1 NDV毒株分离时间统计图
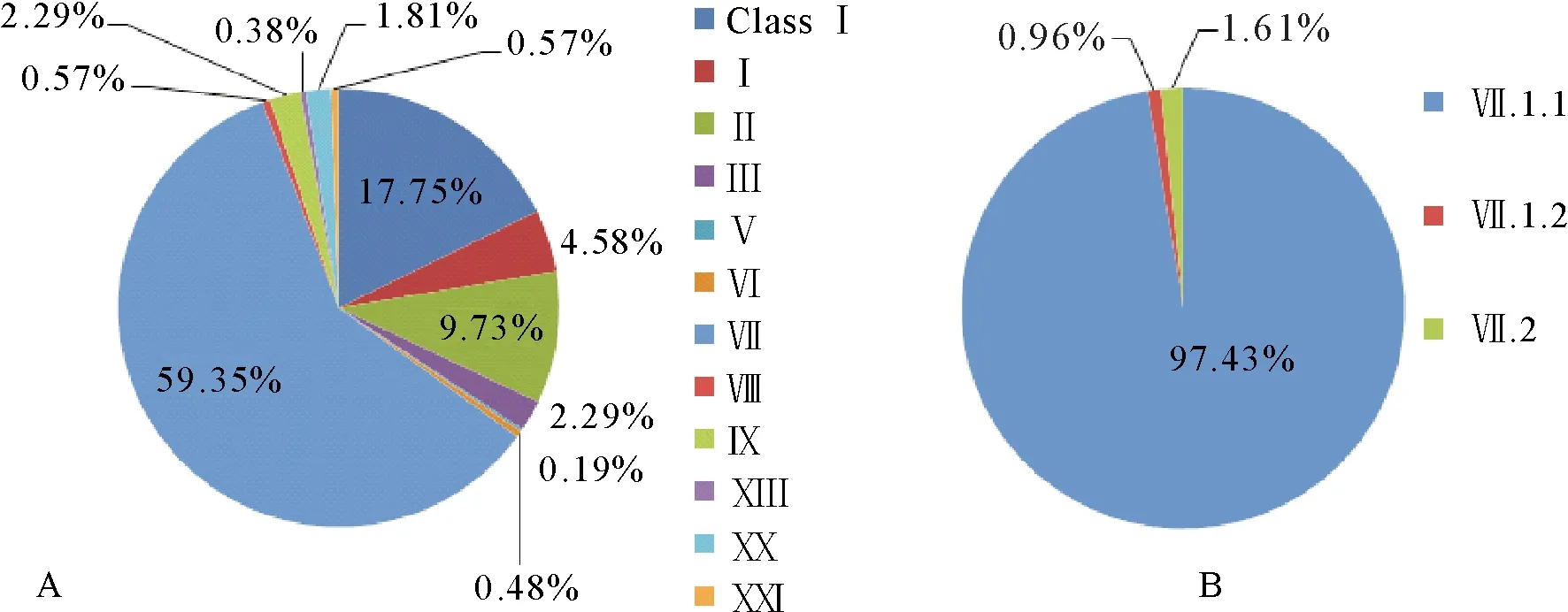
图2 鸡源NDV基因型统计图

图3 水禽源NDV基因型统计图
3 Class Ⅱ类NDV
3.1 基因Ⅰ型和Ⅱ型NDV
Class Ⅱ类基因Ⅰ型和Ⅱ型NDV多为弱毒株,广泛存在于北美、南美、非洲、亚洲和欧洲的家禽、水禽和野生鸟类中。商品化的V4 和Ulster/chicken/Ireland/1967疫苗株属于基因Ⅰ型,而La Sota属于基因Ⅱ型[7]。我国14.31%的鸡源NDV(图2)和29.46%的水禽源(图3)NDV属于这两种基因型。Ⅰ型分为4个基因亚型,我国2004年首次分离到基因Ⅰ型NDV[11],以1.2亚型居多。Ⅱ型NDV首次报道于北美,包括强毒株和弱毒株。我国分离的基因Ⅱ型NDV都为弱毒株,与疫苗株存在高度同源性,可能是由于使用弱毒活疫苗造成的散毒[12]。
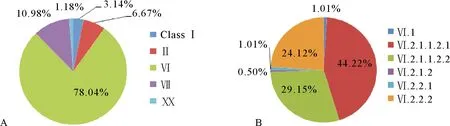
图4 鸽源NDV基因型统计图
3.2 基因Ⅲ型NDV
基因Ⅲ型NDV在20世纪30年代分离于日本,20世纪70年代在巴基斯坦也有发现[7]。1969年我国台湾暴发了由基因Ⅲ型NDV引起的ND[13],之后一直没有基因Ⅲ型强毒的分离报道,而且国内偶尔分离到的基因Ⅲ型毒株也与Mukteswar疫苗株具有很高的同源性(99.7%)[14]。目前,已经禁止使用中等毒力Mukteswar疫苗,防止疫苗株毒力返强带来的风险[3,14]。
3.3 基因Ⅵ型NDV
基因Ⅵ型NDV起源于20世纪60年代末和70年代初的中东地区[7],并传播至世界各地,导致了ND第3次世界大流行。1985年我国首次报道由基因Ⅵ型NDV引起的鸽ND[15],之后在我国鸽中频繁分离到该类型的NDV(图5),特别是华南、华东、西北和东北地区[15-16]。鸽副黏病毒1型(PPMV-1)主要属于Class Ⅱ类基因Ⅵ型NDV(78.04%)(图4)。鸡和水禽等宿主的Ⅵ型占比仅在0.48%~0.55%,而且92.17%基因Ⅵ型病毒分离自鸽(图6)。表明基因Ⅵ型NDV对鸽表现出很强的宿主嗜性。该基因型NDV的F蛋白裂解位点具有强毒特征,致病性试验显示鸡致病性较弱,甚至无致病性,但是对鸽表现出强致病性[15]。基因Ⅵ型NDV分为7个亚型,我国至少存在6种基因亚型,Ⅵ.2.1.1.2.1和Ⅵ.2.1.1.2.2亚型最为流行(图4)。基因Ⅵ型NDV与疫苗株La Sota的F和HN基因核苷酸序列相似相均小于85%,表现出明显的抗原差异[15-16],这些疫苗株不能对鸽提供完全保护。除食用外,赛鸽在基因Ⅵ型NDV的传播中发挥重要作用。为了保护家鸽和赛鸽不被NDV感染,针对鸽的基因Ⅵ型NDV疫苗的研发刻不容缓。

图5 鸽源Class Ⅱ类基因Ⅵ型NDV分离统计
3.4 基因Ⅶ型NDV
基因Ⅶ型NDV具有广泛的宿主范围和地理分布。基因Ⅶ型NDV在20世纪80年代早期就已经在东亚地区存在[17]。1984年和1995年我国台湾暴发了由基因Ⅶ型NDV引起的严重疫情。一株基因Ⅶ.1.1亚型NDV在1984年首次分离到,并在1995年成为优势基因型,在鸡群中迅速传播[13]。2000年首次报道了基因Ⅶ型NDV在中国大陆存在[18]。一直以来人们认为水禽是NDV的天然宿主,即使感染了对鸡有强致病性的毒株,也很少或不表现临床症状。然而,1997年我国首次报道了鹅副黏病毒病,典型症状与鸡相似,具有较高的发病率和死亡率,之后证明是基因Ⅶ型NDV引起的[19]。1995年以来该型NDV一直处于流行趋势,尤其是2000年-2012年。我国近年采取生物防控结合疫苗免疫,基因Ⅶ型NDV分离株有所下降但在家禽中仍存在(图7)。基因Ⅶ型NDV正在引起世界ND第4次大流行,用的疫苗株(基因型Ⅰ和Ⅱ)与基因Ⅶ型毒株的遗传距离为18.3%~26.6%[20]。因此,这些疫苗不能完全保护家禽,不能有效防止病毒复制和排泄。基于我国家禽中分离的NDV强毒株主要基因型为Ⅶ.1.1(图2和3),应加快鸡源和水禽源基因Ⅶ.1.1亚型ND疫苗的研发。

图6 各宿主基因Ⅵ型NDV统计
3.5 基因Ⅷ型NDV
20世纪60年代至90年代在马来西亚、新加坡、中国、土耳其、意大利、阿根廷和南非等地分离到该型NDV[7,13,21]。我国2011年从斗鸡中重新分离到基因Ⅷ型NDV[21]。近些年未见该基因型毒株的分离报道。
3.6 基因Ⅸ型NDV
在家禽、水禽和野生鸟类体内分离到Ⅸ型毒株,并且能够使水禽表现出临床症状[22]。2008年-2011年在我国秦岭和2013年-2015年广东省的健康野鸟中分离到Ⅸ型NDV,说明野鸟可能是该型病毒的天然宿主,且基因Ⅸ型NDV在家禽和野生鸟类中循环存在并维持着较低的进化率[22-23]。
3.7 基因Ⅻ型NDV
首次分离于秘鲁的鸡群,属于Ⅻ.1亚型[7,24]。我国于2010年-2011年首次从鹅群中分离到基因Ⅻ型NDV。2017年和2019年从广东省健康鹅群中再次分离到该类型的毒株。遗传进化树显示,从我国分离到基因Ⅻ型的NDV属于Ⅻ.2亚型,与南美洲的毒株并无流行病学联系[24]。该型毒株属于强毒株,能引起鹅ND暴发,应持续监测鹅群中该型病毒。

图7 我国Class Ⅱ类基因Ⅶ型NDV分离情况统计
4 展望
现有的疫苗虽然降低了ND的发病率和死亡率,但是不能够完全保护,以非典型ND为主的疫情还时有发生。本文综合分析近20多年我国NDV分离株的流行情况,发现水禽(尤其是鸭类)携带大量Class Ⅰ类弱毒株,在Class Ⅰ类NDV的遗传进化中发挥重要作用;Class Ⅱ类基因Ⅵ型NDV对鸽子有很强的宿主嗜性,是引起鸽ND的主要基因型,尤其是Ⅵ.2.1.1.2亚型;基因Ⅶ.1.1亚型NDV是造成我国鸡和水禽(尤其是鹅类)ND发生的优势基因型。未来研发重点应针对鸡和水禽的基因Ⅶ.1.1型疫苗;针对鸽子的基因Ⅵ.2.1.1.2型疫苗的研发;针对水禽Class Ⅰ类ND疫苗研发的储备,防止Class Ⅰ类弱毒株突变为强毒株带来的潜在风险。最大限度提高疫苗免疫效果,减少NDV流行株在免疫家禽体内的复制和排泄,更好的防控ND在我国的暴发。
