长江口潮间带底栖生物生态及变化趋势
2022-03-31陈思思周红宏夏利花刘鹏霞杨幸幸邓邦平
杨 颖,陈思思,周红宏,夏利花,刘鹏霞,杨幸幸,邓邦平,*
1 国家海洋局东海环境监测中心,上海 201206
2 自然资源部海洋生态监测与修复技术重点实验室,上海 201206
长江口是世界特大型淤泥质三角洲河口,长江口潮滩湿地是我国重要河口湿地,具有丰富的自然资源和重要生态系统服务价值。长江口及其邻近水域滩涂中的潮间带生物,在生物链中扮演着重要的角色,是生态系统中不可或缺的一部分,在物质循环和能量流动中有着不可取代的作用。由于底栖动物栖息于潮间带沉积物中,不易移动且对环境变化比较敏感,对潮间带湿地环境状况有较好的指示作用[1],众多学者对长江口潮间带底栖生物的群落演替[2—3]、季节变化[4]、生物多样性与生态学状况[5—8]等进行研究,空间分布上,覆盖了长江口崇明岛[3, 9—10]、九段沙[4, 11]、北支[12]、长兴岛、横沙岛[13]和南汇边滩[14]等典型河口潮间带。已有的研究成果从不同角度对长江口潮间带底栖生物生态及群落演变情况进行了探讨,但针对长时序群落结构演变状况的研究尚不多见。近30年以来,长江口潮间带湿地经历了大规模围垦[15—16]、渔业开发[15]、过度放牧[15,17]、陆源污染[15,18]以及互花米草入侵[19—22]的严重影响,对潮间带底栖生物生态系统造成一定破坏。本文基于在长江口海域潮间带固定断面多年连续底栖生物监测数据,评价了近30年来在各种人类活动叠加影响下的潮间带底栖生物生态现状及变化趋势,以期为长江大保护国家战略及长江保护法实施提供支持。
1 材料与方法
1.1 数据来源
本文中现状评价采用东海监测中心2019年8月在长江口生态监控区海域的3个区域潮间带(崇明东滩、南汇边滩和嵊山岛潮间带,分别简称为CM、NH、SS)布设的9条潮间带断面底栖生物调查数据,断面位置如图1。每条潮间带分别在高、中、低潮区采集潮间带生物样品(定量和定性,表1),滩涂定量样品采集0.25 m×0.25 m×0.30 m的样方4个,野外淘洗装瓶,用福尔马林固定后贴上标签,注明采样时间地点;岩石岸取2个样方合为1个样品。定性样品将该站附近出现的生物种类收集齐全。潮间带生物种类组成、栖息密度、生物量监测方法采用计数法[23]。崇明东滩和南汇边滩总体为淤泥质潮间带,其中崇明东滩中断面高潮区为人工护岸,类同于岩石基质;嵊山岛均为基岩质潮间带。

图1 长江口海域潮间带底栖生物采样断面示意图

表1 各断面及采样点编号与基质
文中近30年潮间带底栖生物趋势变化评价数据均来源于东海监测中心历年在该区域的业务化调查与监测数据,样品采集与监测方法同上,监测季节均为夏季。
1.2 多样性评价
采用种类多样性指数H′ (Shannon-Weaver index)、种类丰富度指数d(Margalef′s index)、均匀度指数J′ (Pielou index)和优势度指数Y对海洋生物多样性进行评价[23],计算公式如式(1)—(4)。
H′=-∑Pilog2Pi
(1)
d=(S-1)/log2N
(2)
J′=H′/log2S
(3)
Yi=ni/N×fi
(4)
式中,S为种类数;N为所有种的个体总数;Pi为样品中第i种个体数占总个体数的比例。fi为第i种在各样品中的出现频率;ni为样品中第i种生物个体数。取Yi≥0.02的种类为优势种类[24]。
1.3 群落结构分析
采用等级聚类(Cluster)、非度量多维标度(MDS)等多变量分析方法分析底栖生物群落的空间格局[25];多变量分析统计结果应用PRIMER 5.0[26]软件统计生成。
2 结果与分析
2.1 种类组成
长江口潮间带共鉴定出底栖生物9门57种(表2),其中软体动物24种,节肢动物17种,环节动物10种,脊索动物2种,扁形动物、棘皮动物、刺胞动物和纽形动物各1种;软体动物占比最高,为42.1%。潮间带定量样品共鉴定出46种,各断面种类数范围为7—31种,平均单条断面种类数为8种;定性样品共鉴定39种,各断面种类数范围5—19种。

表2 大型底栖动物种名录(2019)
崇明东滩3条潮间带断面共鉴定出底栖生物18种。其中节肢动物、软体动物各7种,分别占38.9%;环节动物2种,占11.0%;脊索动物、腔肠动物各1种,分别占5.6%。优势种共有7种,为绯拟沼螺Assiminealatericea、谭氏泥蟹Llyoplaxdeschampsi、紫游螺Neritinaviolacea、无齿相手蟹Sesarmadehaani、弹涂鱼Periophthalmuscantonensis、河蚬Cobiculaflaminca、尖锥拟蟹守螺Cerithidealargillierti。
南汇边滩3条断面共鉴定出底栖生物14种。其中软体动物5种,占35.7%;环节动物、节肢动物各3种,分别占21.4%;脊索动物2种,占14.3%;纽形动物1种,占7.2%。优势种共6种,为中国绿螂Glauconomechinensis、谭氏泥蟹、丝异须虫Heteromastusfiliformis、绯拟沼螺、尖锥拟蟹守螺、光滑狭口螺Stenothyraglabar,其中,软体动物4种、节肢动物和环节动物各1种。南汇边滩和崇明东滩均为泥相潮间带,谭氏泥蟹、绯拟沼螺和尖锥拟蟹守螺为共有优势种。
嵊山岛潮间带为岩相潮间带,3条断面共鉴定出底栖生物8门38种,其中软体动物15种,占总种类数的39.5%,节肢动物12种,环节动物7种,扁形动物、棘皮动物、刺胞动物和纽形动物各1种。优势种共2种,分别为日本笠藤壶和条纹隔贻贝。
2.2 生物密度和生物量
崇明东滩、南汇边滩、嵊山岛3个监测区域潮间带底栖生物密度和生物量如表3所示。生物密度和生物量均值均为嵊山岛潮间带最高,崇明东滩生物密度低于南汇边滩,但生物量高于南汇边滩。
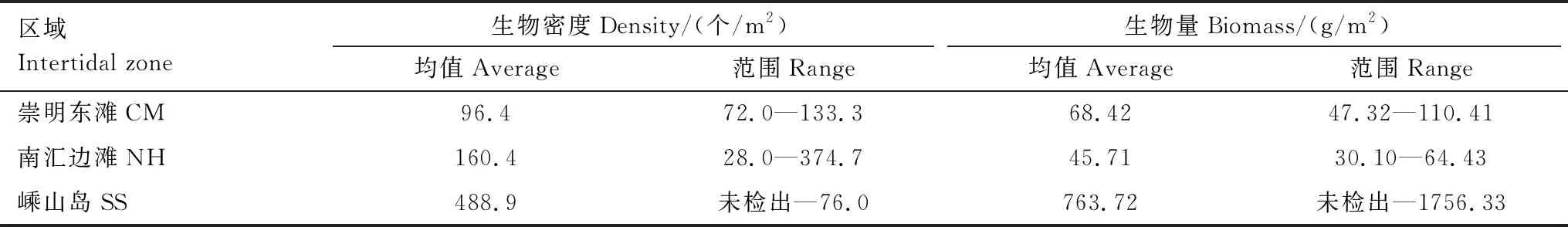
表3 长江口各区域潮间带底栖生物密度和生物量统计表
崇明东滩和南汇边滩均为淤泥质潮滩,但生物密度和生物量的分布具有较大不同(图2)。崇明东滩潮间带生物密度和生物量分布均为高潮区>低潮区>中潮区,南汇边滩生物密度为中潮区>低潮区>高潮区,生物量则为高潮区>中潮区>低潮区。南汇边滩高潮区生物量远高于中、低潮区的原因是南汇南和南汇中断面高潮区监测到较高生物量的无齿相手蟹,其生物量占到南汇边滩3条断面高潮区总生物量的60.7%。
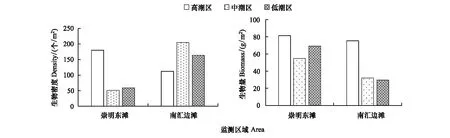
图2 崇明东滩和南汇边滩各潮区底栖生物平均生物密度和生物量分布
嵊山岛岩相潮间带生物密度为低潮区最高,生物量则为中潮区最高(图3)。嵊山岛潮间带3条断面底栖生物分布差异巨大,大玉湾断面(CJD8)未采集到定量生物样品;后陀湾断面(CJD7)定量监测到底栖生物8种,平均生物量1756.01 g/m2;鳗咀头断面(CJD9) 定量监测到底栖生物22种,平均生物量535.05 g/m2。中潮区生物量最高是由于后陀湾中潮区监测到大量日本笠藤壶,其生物量占后陀湾断面生物量的50%,占到两条断面总生物量的37.6%。

图3 嵊山岛各潮区底栖生物平均生物密度和生物量分布
2.3 生物多样性
一般来说,种类多样性较高的生物群落,其生态环境较好,生物群落有较好的稳定性、较强的抗变能力和修复能力。以站位生物为单元统计H′、J′、d3项指数,获得各断面各项指数均值见图4。从图中可以看出,崇明和南汇共6条泥相断面总体上种类组成多样性不高,可能与长江口环境复杂多变有关。其中,CJD1断面的H′、J′最大,d也大,该断面生物多样性最高,CJD6和CJD5断面也较高,CJD2断面多样性低;嵊山CJD9基岩断面的3项指数均显著大于CJD7断面,该断面生物多样性远高于CJD7断面。3个潮区多样性比较分析,高潮区多样性指数、均匀度指数和丰富度指数分别为1.19、0.62、0.56,中潮区分别为1.57、0.71、0.96,低潮区分别为1.67、0.70和0.93,总体上低潮区生物多样性优于中潮区和高潮区。

图4 长江口潮间带各断面底栖生物多样性指数统计表
对崇明东滩和南汇边滩6条泥相潮间带断面各潮区多样性指数进行统计(图5),低潮区多样性指数和丰富度指数最高,中潮区均匀度指数最高,高潮区的多样性指数和丰富度指数均最低。相对而言,低潮区生物多样性总体优于中潮区,中潮区略好于高潮区。
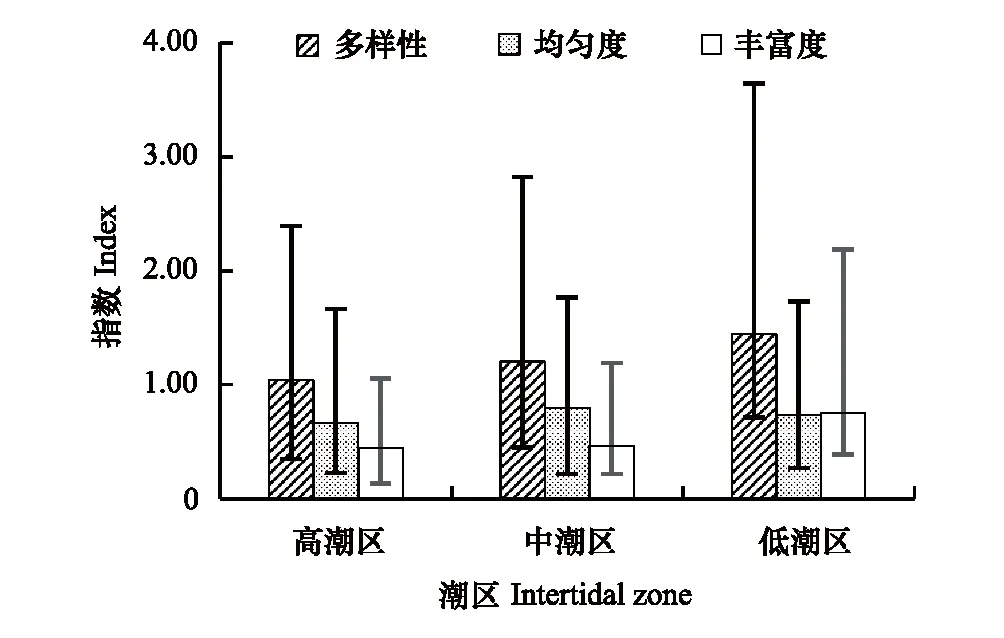
图5 崇明东滩与南汇边滩各潮区多样性指数比较
对嵊山岛后陀湾和鳗咀头岩相断面各潮区多样性指数进行统计(图6),中潮区的多样性指数、均匀度和丰富度均为最高,低潮区次之,高潮区最差,总体生物多样性为中潮区优于低潮区,优于高潮区。
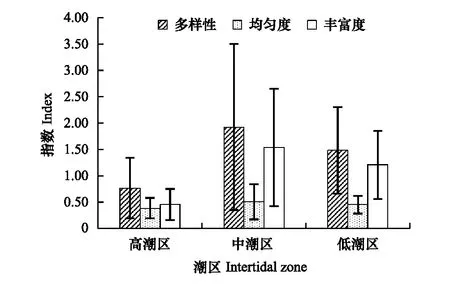
图6 嵊山岛各潮区多样性指数比较
2.4 群落结构分析
以断面底栖生物种类和密度绘制Bray-Curtis 相似性系数聚类树状图(图7),依据分析结果长江口潮间带明显分为两个大群组,第1 组为崇明东滩、南汇边滩共6条断面,为泥相断面;第二组为嵊山后陀湾和嵊山鳗咀头断面,为岩相断面,证明不同栖息环境是影响生物群落分布的主要影响因素之一。以各断面高、中、低潮区采样点绘制Bray-Curtis 相似性系数聚类树状图(图8),以相似度大于50%来划分,发现高潮区站位成群或与个别中潮区站位成群,低潮区站位成群或与个别中潮区站位成群。嵊山潮间带可以划分为3个小群组,分别为(1)后陀湾高、中潮区和鳗咀头高潮区,(2)鳗咀头中、低潮区,(3)后陀湾低潮区。崇明东滩和南汇边滩潮间带可以划分为8个小群组,分别为(1)南汇中断面中、低潮区和南汇北断面中潮区,(2)南汇北断面低潮区和崇明中断面中潮区,(3)崇明南断面低潮区和南汇南断面低潮区,(4)南汇南断面高、中潮区,崇明南断面高、中潮区,(5)崇明北断面、南汇北断面和南汇中断面高潮区,(6)崇明北断面中潮区、崇明中断面高潮区;(7)崇明北断面低潮区和(8)崇明中断面低潮区虽在一个群组,但相似度低于40%。可见高、中、低潮区的分布有较大差异,表明潮汐也是影响潮间带大型底栖生物群落结构的主要因子。
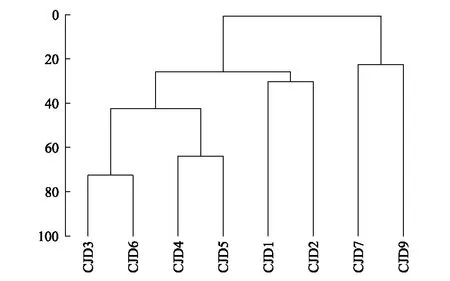
图7 各断面相似性系数聚类树状图

图8 各站点相似性系数聚类树状图
采用Bray-Curtis相似系数转换距离系数绘制非度量多维标度(MDS)图,以断面为单位MDS图展示的群落结构分布与相应的Bray-Curtis 相似性系数聚类树状图结论一致(图9),分为泥相潮间带和岩相潮间带两个清晰的群组,Stress 值0.01,CJD3和CJD6相似性最高,几乎重合。以各断面高、中、低潮区采样点为单位绘制的MDS图(图10),Stress 值0.05,可以较清晰的体现高、中、低潮区的差异,与图8相似性聚类分析结果可相互验证。
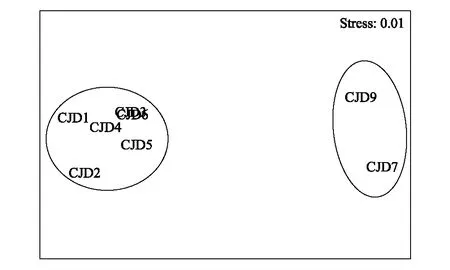
图9 各断面非度量多维标度图
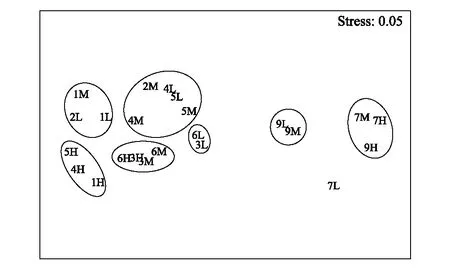
图10 各站点MDS图
2.5 多年变化趋势分析
对崇明东滩(CJD1—CJD3)、南汇南断面(CJD4)、嵊山后陀湾断面(CJD7)的近30年监测资料进行统计分析,评价潮间带底栖生物变化趋势。
2.5.1崇明东滩
1990—2019年8月,崇明东滩潮间带底栖生物种类数变化范围11—48种(图11),平均23种。种类数按峰谷分布可分成3个时段,1990年监测到48种;2000—2009的10年间种类变化范围11—33种,平均23种,总体趋势震荡上升;2010—2019年的10年间种类数变化范围16—29种,平均21种,总体趋势震荡下降。

图11 崇明东滩潮间带底栖生物种类数变化
崇明东滩潮间带底栖生物密度与生物量波动较大(图12),近30年生物密度范围为31—12352个/m2, 2009年在崇明东滩北断面低潮区采集到大量的中间拟滨螺(49600个/m2)和光滑狭口螺(54400个/m2),导致2009年生物密度极大值,其他年份密度均值为741个/m2。生物量范围为5.03—235.04 g/m2,各年份平均生物量为94 g/m2。对各年份数据添加指数型趋势线,发现近30年来生物量与生物密度总体均呈现下降的趋势,生物量从2003年最高点震荡下降至2014年最低点,2014年后有所反弹,生物密度变化趋势基本一致。
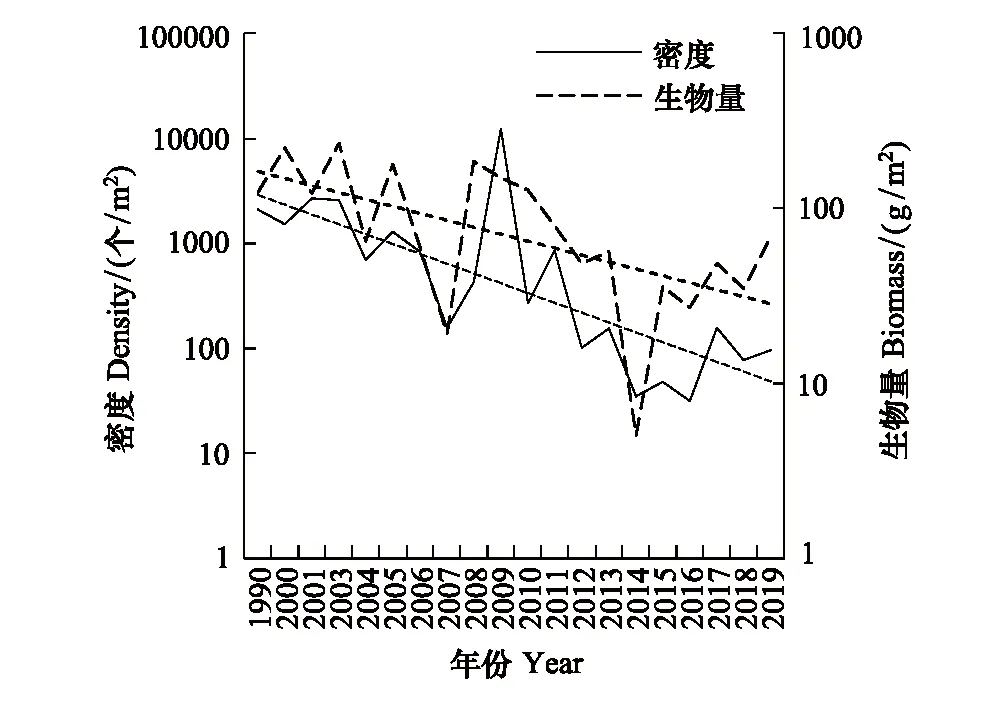
图12 崇明东滩潮间带底栖生物密度和生物量变化情况
2.5.2南汇南断面
1993—2019年,南汇南断面(CJD4)潮间带底栖生物种类数变化范围3—24种(图13),平均10种,2016年8月种类数最低,2009年8月份最高。1993—2009年略显上升趋势,2009年后略显下降趋势。总体上南汇南断面潮间带底栖生物种类数不丰富。
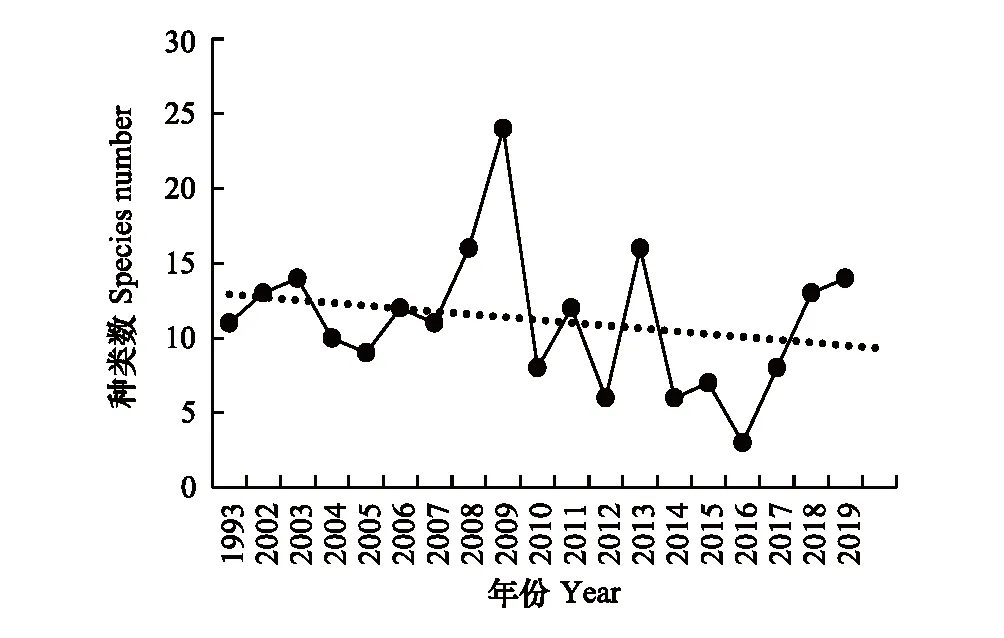
图13 南汇南断面潮间带底栖生物种类数变化
1993—2019年,南汇南断面生物密度与生物量波动较大(图14)。生物密度范围为7—4628个/m2,各年份平均密度为449个/m2。生物量范围为1.9—2526 g/m2,各年份平均生物量为210 g/m2。2009年南汇南断面高潮区采集到大量泥螺,导致该年份生物量均值的极大值。生物量与生物密度总体无明显变化规律,均呈现大幅震荡,2011年以后生物密度明显低于1993—2011年。生物量分别于2002年2009年有两个峰值,2007年为最低值,其余年份在一定范围内波动。2011年以后生物量和生物密度变化趋势基本一致。
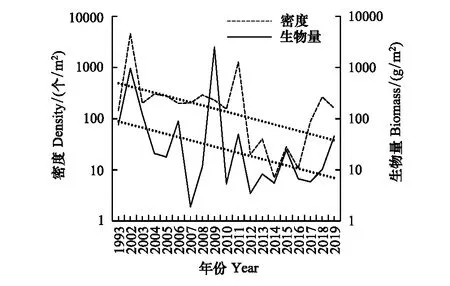
图14 南汇南断面潮间带底栖生物密度和生物量变化
2.5.3嵊山后陀湾断面
1990—2019年,嵊山后陀湾断面(CJD7)潮间带底栖生物种类数变化范围15—84种(图15),各年份平均33种,2015年8月鉴定的种类数最低,2009年8月份最高。各年份种类数波动较大,但总体上呈现下降的趋势。

图15 嵊山后陀湾潮间带底栖生物种类数变化
嵊山后陀湾断面生物密度与生物量变化趋势总体一致(图16),变化趋势基本呈反“S”型,1990—2001年生物密度和生物量逐渐上升,2001—2014年缓慢下降,2014年以后有所回升。生物密度范围为124—22720个/m2,各年份平均密度为2530个/m2。生物量范围为183—11818 g/m2,各年份平均生物量为3230 g/m2。
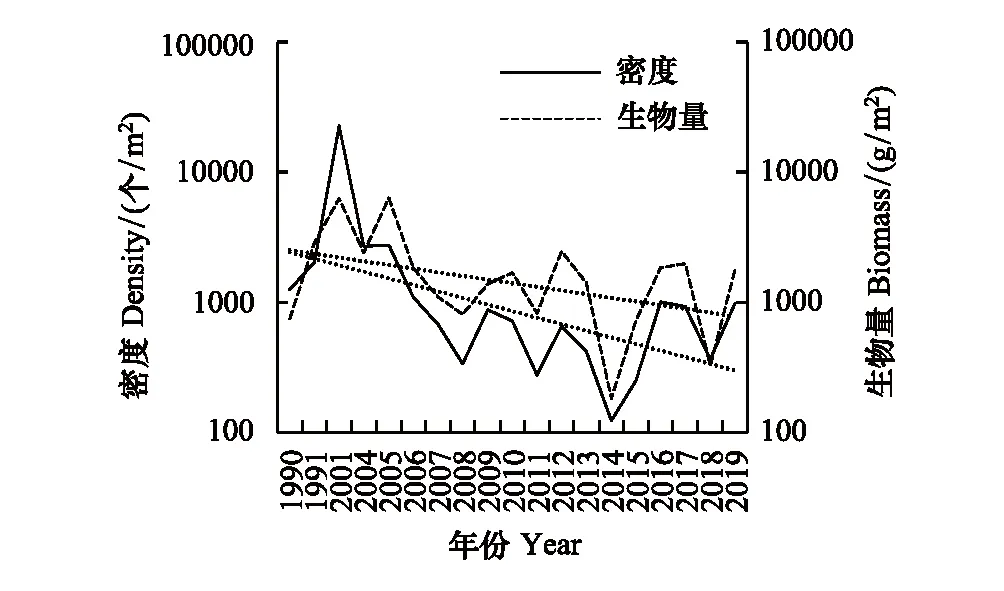
图16 嵊山后陀湾潮间带底栖生物密度和生物量变化趋势
3 讨论
3.1 种类组成特征
崇明东滩和南汇边滩分别位于长江口南支主泓南北两侧,受潮汐作用影响,盐度变化明显。各断面的位置分布上,崇明东滩北断面靠近北支水域,南断面靠近南支水域,北、中、南断面盐度依次降低,南汇边滩北、中、南断面则盐度依次升高。盐度是影响底栖生物分布的重要非生物因子[27],栖息于河口泥相潮间带的生物表现出河口低盐种、半咸水种和淡水种共存的特点。河蚬、光滑狭口螺等是生态幅较宽的淡水种,主要生于活河湖水域,也可生活在河口低盐潮间带区域,在崇明南断面淡水区域和南汇南断面低盐区域均有分布;绯拟沼螺、谭氏泥蟹和无齿相手蟹、丝异须虫等典型河口咸淡水种类在大部分断面均有分布;尖锥拟蟹守螺、紫游螺等咸水种类主要分布在崇明北和南汇南断面等盐度较高区域,南汇南断面还分布有少量广盐性日本刺沙蚕。
嵊山岛常年盐度较为稳定,生物类群多由生命周期长的固着海洋生物组成,如日本笠藤壶和条纹隔贻贝等。后陀湾和鳗咀头断面处于外海开敞性岩岸,风浪较大,生物的垂直分布明显表现出外海高盐特征,且分带现象明显。高潮区分布有矮拟帽贝、条纹隔贻贝、嫁虫戚、日本笠藤壶、小结节滨螺、龟足、粗腿厚纹蟹、红条毛肤石鳖、紫贻贝等。中潮区生物种类最多,后陀湾断面中潮区有明显的日本笠藤壶生物带,扁模裂虫、短石蛏、单齿螺、条纹隔贻贝、海葵、海蟑螂、疣荔枝螺、粗腿厚纹蟹、紫贻贝、红条毛肤石鳖、近江牡蛎、黑凹螺、寄居蟹、棘刺牡蛎、甲虫螺、覆瓦小蛇螺、黑凹螺、尖额麦杆虫、钩虾、腔齿海底水虱等在中潮区也有分布。低潮区主要种类有条纹隔贻贝、日本笠藤壶、尖额麦杆虫、钩虾、软背鳞虫、疣荔枝螺、海葵、腹侧凹盖鳃水虱等。
3.2 潮位与底质类型对潮间带底栖生物分布的影响
不同潮区大型底栖动物的分布与其本身的生物学特性有关,泥相潮间带高潮区以相手蟹属Sesarma和拟沼螺属Assiminea等种类为主,是对缺水环境适应能力较强的半陆生种类,这些个体平均生物量大,致使高潮区生物量较高;低潮区的种类对缺水适应能力较弱,主要有多毛类和双壳类,丰度和生物量均较低。崇明东滩高潮区生物量和生物密度远高于中、低潮区,高潮区优势种为绯拟沼螺、谭氏泥蟹、紫游螺、无齿相手蟹等。南汇边滩高潮区生物量亦远高于中、低潮区,因高潮区监测到的大量无齿相手蟹,其生物量占到南汇边滩3条断面高潮区总生物量的60.7%。崇明东滩中断面高潮区由于修建人工建筑,出现了鳞笠藤壶等岩生种类。
岩相潮间带与淤泥质明显不同,嵊山岛潮间带中、低潮区生物密度比较接近;生物量则高、低潮区比较接近,由于后陀湾中潮区生长大量日本笠藤壶,导致中潮区生物量远高于高、低潮区。嵊山海域位于波浪作用强烈的敞开性海域,中潮区的优势种为耐受波浪冲积的日本笠藤壶,分布特征与嵊泗岛潮间带相似[28]。嵊山岛潮间带与邻近的6个海岛(嵊泗、衢山、岱山、六横、舟山、朱家尖)相比(表4),高、中、低潮区的底栖生物生物量均远高于6个海岛潮间带的相应平均值;除高潮区外,中、低潮区生物密度也远高于6个海岛潮间带的相应平均值[28],生物量与生物密度的分布规律也不同,可能由于嵊山岛位于海域最东侧,远离大陆,受人类影响最小。
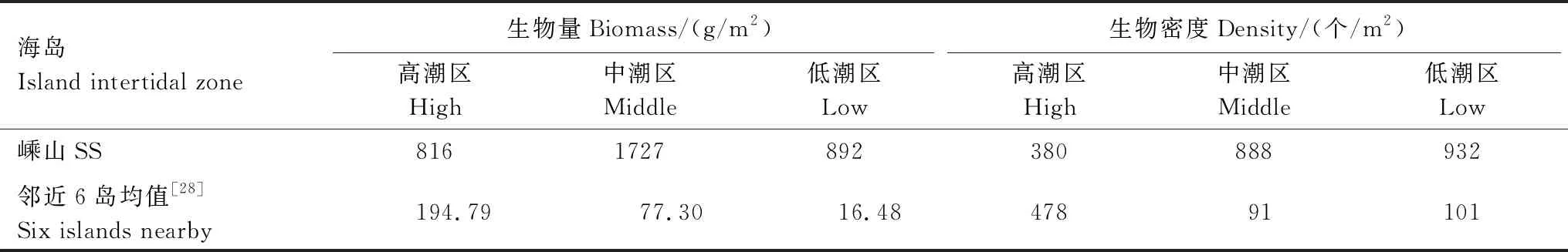
表4 嵊山岛潮间带底栖生物密度、生物量与邻近6岛的比较
3.3 潮间带底栖生物优势种变化
长江口3个调查区域潮间带底栖生物优势种类组成变动较大。2014—2019年,崇明东滩、南汇边滩和嵊山岛出现的优势种类分别为15、9和12种,均未发现同一区域历年均出现的优势种。崇明东滩优势种出现频率不低于50%的仅1种,为谭氏泥蟹(83.3%),谭氏泥蟹和绯拟沼螺逐渐成长为群落的主要优势种类;南汇边滩优势种出现频率不低于50%的仅2种,为光滑狭口螺(83.3%)和谭氏泥蟹(50.0%),光滑狭口螺在群落种优势地位呈现较大的波动,2015和2018年为最主要的优势种类,但2019调查的主要优势种类为中国绿螂和谭氏泥蟹;嵊山岛势种出现频率不低于50%的亦为2种,为紫贻贝(83.3%)和鳞笠藤壶(50.0%),紫贻贝在群落中的优势度呈现降低趋势,日本笠藤壶逐渐演替为主要的优势种类。调查资料表明,2004—2006崇明东滩潮间带优势度前3的优势种为硬爪始根钩虾、拟沼螺和谭氏泥蟹[5];2011年崇明东滩调查优势种主要为光滑狭口螺、谭氏泥蟹、焦河篮蛤和角形环棱螺Bellamyaangularia,优势度较高,均超过0.1,其中角形环棱螺在其他年份未发现;2015年崇明东滩优势度前3的优势种为中华拟蟹守螺、谭氏泥蟹和绯拟沼螺;2019年该区域优势度前3为绯拟沼螺、谭氏泥蟹和紫游螺,崇明东滩和南汇边滩潮间带出现频率最高的的优势种仅1种为谭氏泥蟹。滩涂生物群落的演替,也直接反映了长江口多元的人类活动干扰等因素引起的滩涂生境的变化。
3.4 干扰对群落结构历史演变的影响
本文对长江口3个区域的潮间带近30年底栖生物状况演变趋势进行统计,发现崇明东滩、南汇南断面和嵊山后陀湾断面的生物密度和生物量均呈下降的趋势,其中崇明东滩和嵊山后陀湾属于持续下降,南汇南断面为剧烈震动下降。生境变化和人为干扰是造成底栖动物群落密度和生物量锐减、生物多样性降低的主要原因[27]。Hobbie[29]梳理的人类活动对河口造成的影响中对河口潮间带底栖生物造成直接影响的有4点,包括生境破坏、流域水沙变化、渔业资源过度开发及外来物种物种入侵等。这些影响因素在长江口均有明显表现,长江口区域潮滩湿地生境和生物多样性变化主要受以下人类活动的影响:
(1)潮滩圈围与冲淤变化。1989—2007 年,长江口总共匡围了448.24 km2边滩湿地,主要分布在崇明东滩、南汇边滩和横沙东滩等区域,造成长江口潮间带湿地生境损失明显[16]。近30年来,长江口崇明东滩湿地、横沙东滩湿地和南汇边滩湿地由于促淤围垦,不断向东扩张。但2002—2004年间由河口浅滩由淤积转为侵蚀38.10 km2,边滩湿地总体转为侵蚀趋势明显[16]。受长江流域来沙减少、长江口深水航道建设和促淤造地等工程设施影响,湿地资源的增生能力受限。多年来长江口潮滩部分区域处于不稳定状态,对潮间带底栖生物多样性应有一定影响。
(2)资源过度开发。与历史资料相比,崇明东滩一些经济物种如彩虹明樱蛤、泥螺等已不再是优势种且丰度很低。1980年代资源量较大的石蟥目前几近绝迹,1990年代中后期还比较丰富的泥螺和缢蛏目前的资源量也急剧下降[8]。其资源量的下降可能与环境变迁有关外,人为过度捕捞更可能是资源量下降的原因之一[14]。1990s嵊山后陀湾、大玉湾和鳗咀头断面的生物密度、生物量并无明显差异[30],2004年大玉湾断面生物密度和生物量分别降至1990s的50%和80%左右,2015年又进一步快速下降到6%和3%,2016年以后大玉湾断面定量样品均未检出。大玉湾位于嵊山岛西部与枸杞岛的夹角内部,风浪最小,断面附近有嵊泗岛上唯一的沙滩,是岛上旅游资源较丰富的区域。潮间带底栖生物资源量减少应与开发活动和人类采捕有一定关联。
(3)外来物种入侵。由于引种和自然扩散导致互花米草泛滥,崇明东滩和南汇边滩均有大量互花米草分布[19,20]。在长江口崇明岛和九段沙湿地,互花米草入侵后形成的单优群落拥有发达的地下结构,这会改变土壤的物理和化学性质,这些变化会驱动盐沼中底栖无脊椎动物群落特征的改变[22]。互花米草入侵海三棱藨草群落后,大型底栖无脊椎动物群落的物种组成没有显著的不同,但是大型底栖无脊椎动物的多样性有显著降低[21-22]。
(4)环境持续恶化。近30年来,长江口海域持续加重的污染和富营养化状态也可能对边滩湿地生态系统造成一定的影响[18],水体富营养化会影响底栖动物的群落结构[31—32],底栖动物的各种特征参数都与有机质污染源在时间和空间上存在明显关系[33]。
4 小结
长江口3个监测区域潮间带共鉴定出底栖生物9门57种,底栖生物密度均值为248.6 个/m2,生物量均值为292.6 g/m2。多样性指数(H′)均值为1.98,均匀度(J′)均值为0.73,丰富度(d)均值为0.91。栖息环境(基岩海岸、淤泥海岸)和潮汐均是影响底栖生物群落结构的重要因素。
根据近30年的监测结果,崇明东滩潮间带底栖生物种类数变化总体平稳,生物量与生物密度均呈现下降的趋势;南汇南断面潮间带底栖生物种类数变化波动较大,生物量与生物密度震荡下降,2011年以后生物密度明显低于1993—2011年;嵊山后陀湾潮间带底栖生物种类数总体呈下降趋势,生物密度与生物量变化趋势基本呈“S”型,1990—2004年生物密度和生物量逐渐上升,2004—2014年缓慢下降,2014年以后有所回升。
