过氧化物还原酶4蛋白对小鼠卵巢颗粒细胞凋亡的影响及机制
2022-03-28邹小飞梁秀茹严正杰崔毓桂刘嘉茵孟艳
邹小飞,梁秀茹,严正杰,崔毓桂,刘嘉茵,孟艳
颗粒细胞通过缝隙连接与卵母细胞进行信号、能量及物质的交换,是卵巢中重要的细胞成分。颗粒细胞凋亡是卵泡闭锁的原因之一[1],异常的卵泡闭锁增加会影响卵巢功能。凋亡是受基因控制的主动死亡过程,分为内源性途径(线粒体途径)、外源性途径(死亡受体途径)和内质网应激诱导的细胞凋亡途径[2]。卵巢氧化微环境的建立和氧化还原稳态的丧失是颗粒细胞凋亡的主要原因[3]。在病理状态下活性氧(reactive oxygen species,ROS)水平增加,抗氧化酶的表达及活性下降,导致ROS在颗粒细胞内进一步蓄积,进而扰乱内质网中正常的氧化还原状态,可引起内质网应激[4]。内质网首先通过诱发未折叠蛋白反应(unfold protein response,UPR)恢复内质网稳态。持续的内质网应激和过量的ROS以正反馈的方式相互作用并相互加强,最终ROS诱导的内质网应激决定细胞的状态或命运,如自噬、凋亡或衰老等[5]。过氧化物还原酶4(peroxiredoxin 4,Prdx4)在哺乳动物细胞中广泛表达,其有2个半胱氨酸残基,通过清除过氧化氢(H2O2)抑制氧化应激,与年龄相关性疾病(如癌症、糖尿病和炎症性疾病等)相关[6-8]。为深入探索Prdx4在颗粒细胞中的作用,本研究采用Prdx4小干扰RNA(small interfering RNA,siRNA)下调小鼠颗粒细胞Prdx4蛋白的表达,探讨下调Prdx4表达水平对颗粒细胞凋亡的影响及其相关机制。
1 材料与方法
1.1 试剂十二烷基磺酸钠-聚丙烯酰胺凝胶电泳(SDSpolyacrylamide gel electrophoresis,SDS-PAGE)凝胶配制试剂盒购自南京源之信生物技术有限公司,蛋白裂解液及5×上样缓冲液、BCA蛋白浓度测定试剂盒购自上海碧云天生物技术有限公司,DMEM/F12购自美国Gibco公司,蛋白酶抑制剂购自美国赛默飞公司,特超敏增强型化学发光显影液购自苏州宇恒生物科技有限公司,含细胞核荧光染料4,6-联脒-2-苯基吲哚(DAPI)的封片液购自德国SBA公司,Prdx4单克隆抗体、B细胞淋巴瘤2关联X蛋白(Bcl-2 associated X protein,Bax)单克隆抗体、CCAAT/增强子结合蛋白同源蛋白(C/EBP homologous protein,CHOP) 单克隆抗体和转录激活因子4(activating transcription factor 4,ATF4)多克隆抗体购自美国Abcam公司,蛋白质二硫键异构酶(protein disulfide isomerase,PDI)单克隆抗体和免疫球蛋白结合蛋白(binding immunoglobulin protein,BIP)单克隆抗体购自美国Cell Signaling Technology公司,ATF6单克隆抗体购自美国Santa Cruz Biotechnology公司,Bcl-2多克隆抗体和裂解的半胱氨酸天冬氨酸蛋白酶3(cleaved caspase 3)多克隆抗体购自美国Proteintech公司。
1.2 实验动物实验小鼠均为SPF级雌性C57BL/6J小鼠,购于浙江维通利华实验动物技术有限公司,饲养于南京医科大学实验动物房中。
1.3 细胞培养取8周龄小鼠,腹腔注射孕马血清8 IU/只,36 h后颈椎脱臼处死,无菌条件下取出卵巢,体式显微镜下用注射器针头刺破卵泡,释放出颗粒细胞,均匀接种入6孔板。另取部分颗粒细胞放置到有盖玻片的培养皿,置于5%CO2、37 ℃细胞培养箱中培养。人卵巢颗粒细胞(KGN)来自于生殖医学国家重点实验室。
1.4 转染siRNA待细胞融合率达到50%~60%,予siRNA转染细胞,以空白组作为对照,12 h后换液,24 h后提取细胞。部分细胞加入戊二醛固定后送南京医科大学电镜中心观察亚细胞结构,部分细胞经蛋白裂解液裂解后取上清检测相关蛋白的表达。
1.5 免疫荧光将细胞爬片置于6孔板中,当KGN细胞在爬片上融合率达70%~80%时,予4%多聚甲醛(paraformaldehyde solution,PFA)固定15 min,0.5%TritonX-100通透20 min,磷酸盐缓冲液(PBS)洗3次,山羊血清室温下封闭30 min后孵育一抗Prdx4(1∶100)及PDI(1∶100)于4 ℃过夜。室温孵育荧光标记的二抗2 h,PBS清洗3次后DAPI室温避光孵育1 min,荧光显微镜下观察。
1.6 细胞凋亡检测siRNA转染细胞24 h后,使用不含乙二胺四乙酸(EDTA)的胰酶消化细胞至细胞漂浮(约5 min),PBS反复清洗后,于4 ℃、2 000 r/min离心3 min,按照试剂盒说明,用流式细胞仪检测细胞凋亡情况。
1.7 蛋白质印迹法(Western blotting)检测各蛋白表达水平收集2组处理后的细胞,BCA法检测蛋白浓度后,取总蛋白样品各40 μg进行Western blotting。将在凝胶上分离的蛋白转移至聚偏氟乙烯(PVDF)膜上(250 mA,80 min),5%脱脂奶粉室温下封闭1 h,加入Prdx4、BIP、ATF4、ATF6、CHOP、Bax、Bcl-2及cleaved caspase 3抗体于4 ℃过夜。次日用TBST洗膜3次后再加入抗辣根过氧化物酶(HRP)标记的羊抗兔IgG(1∶5 000)室温下孵育2 h。TBST洗膜3次后采用化学发光法检测蛋白条带的灰度值,用目的蛋白与内参甘油醛-3-磷酸脱氢酶(GAPDH)及纽蛋白(Vinculin)条带的灰度值之比表示检测蛋白的相对表达量。
1.8 统计学方法所有实验均独立重复3次以上,采用GraphPad Prism7.04统计软件进行数据分析。定量资料用均数±标准差(±s)表示,组间比较采用t检验。P<0.05为差异有统计学意义。
2 结果
2.1 Prdx4在KGN细胞中的亚细胞定位由于PDI特异性定位于内质网,利用免疫荧光共定位,在荧光显微镜下观察绿色荧光(Prdx4)与红色荧光(PDI)位置重合,明确Prdx4主要定位于KGN细胞内质网中。见图1。

图1 Prdx4 在KGN细胞中的亚细胞定位(×600)
2.2 下调Prdx4蛋白表达后小鼠颗粒细胞凋亡的改变Prdx4-siRNA处理组凋亡率较空白对照组增加[(23.900±4.431)%vs.(13.233±1.557)%,n=3,t=3.934,P=0.017],见图2A。Western blotting结果提示,Prdx4-siRNA处理组促凋亡因子Bax及cleaved caspase 3蛋白水平表达较空白对照组高(均P<0.01),抗凋亡因子Bcl-2表达水平较空白对照组下降(P<0.05)。见图2B、表1。
表1 2组小鼠颗粒细胞凋亡相关蛋白相对表达量比较 (±s)

表1 2组小鼠颗粒细胞凋亡相关蛋白相对表达量比较 (±s)
组别 n cleaved caspase 3 Bax Bcl-2空白对照组 3 0.590±0.081 0.866±0.027 1.025±0.051 Prdx4-siRNA处理组 3 1.282±0.154 1.204±0.049 0.787±0.110 t 6.909 10.450 3.409 P 0.002 0.001 0.027
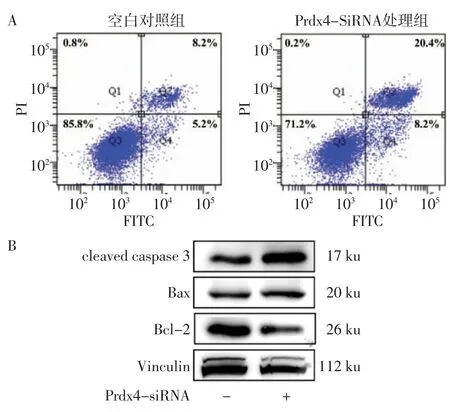
图2 小鼠颗粒细胞下调Prdx4蛋白表达后凋亡情况
2.3 下调Prdx4蛋白表达后小鼠颗粒细胞内质网超微结构改变通过透射电镜对2组小鼠颗粒细胞进行超微结构的观察,发现与空白对照组相比,Prdx4-siRNA处理组内质网肿胀、扩张和增生。见图3。
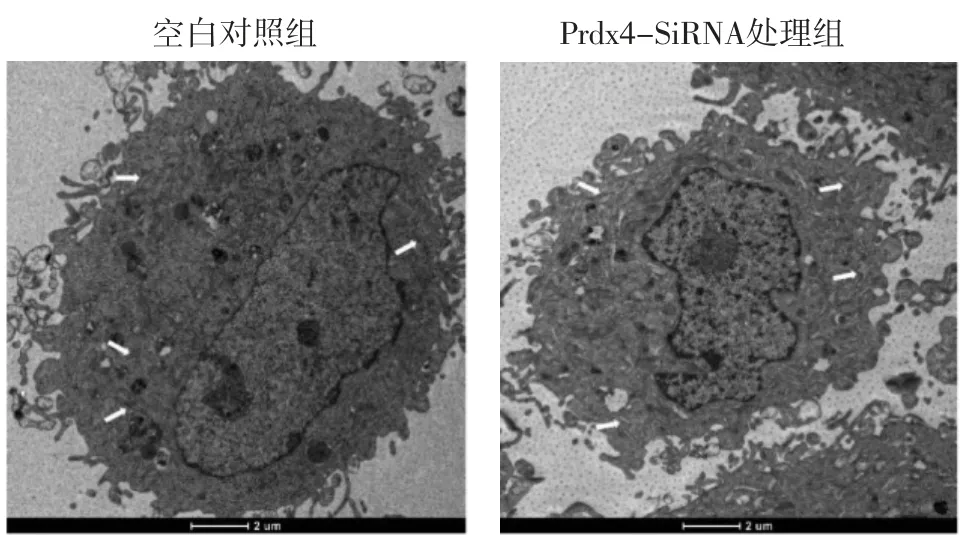
图3 小鼠颗粒细胞下调Prdx4蛋白表达后内质网超微结构的改变(×3 000)
2.4 下调Prdx4蛋白表达后小鼠颗粒细胞内质网应激相关蛋白表达情况采用Western blotting检测2组内质网应激信号通路相关标志物的表达水平,发现Prdx4-siRNA处理组Prdx4蛋白表达水平较空白对照组下降(P<0.01),BIP、ATF4和CHOP表达水平均较空白对照组升高(均P<0.05),ATF6表达水平差异无统计学意义(P>0.05)。见图4、表2。

图4 小鼠颗粒细胞下调Prdx4蛋白表达后内质网应激相关蛋白表达情况
表2 2组小鼠颗粒细胞内质网应激相关蛋白相对表达量比较 (±s)

表2 2组小鼠颗粒细胞内质网应激相关蛋白相对表达量比较 (±s)
组别 n Prdx4 BIP ATF6 ATF4 CHOP空白对照组 3 1.057±0.126 0.563±0.169 0.943±0.106 0.373±0.139 0.567±0.145 Prdx4-siRNA处理组 3 0.469±0.080 0.967±0.162 0.671±0.486 0.959±0.149 1.013±0.162 t 6.859 2.982 0.948 4.975 3.549 P 0.002 0.041 0.397 0.008 0.024
3 讨论
内质网是真核细胞中重要的细胞器,在蛋白质合成、加工修饰、氧化折叠、组装和新生肽链的运输中起着重要作用[9]。内质网氧化还原稳态是其发挥功能的基础。一些生理和病理条件,包括氧化应激、炎症、钙稳态丧失和病原体感染等都会引起内质网应激[10]。在这种情况下,错误折叠和未折叠的蛋白质在内质网中积累,首先通过启动UPR增加蛋白质氧化折叠能力并减少蛋白质氧化折叠负荷来缓解内质网应激[11]。UPR是在内质网的3个信号传感器肌醇酶1α(inositol requiring enzyme 1α,IRE1α)、蛋白激酶R样内质网激酶(protein kinase R-like endoplasmic reticulum kinase,PERK)和ATF6的刺激下启动的。在生理条件下,这3个跨膜蛋白都与内质网分子伴侣蛋白BIP结合,处于非活性状态[12]。当内质网应激发生时,它们与BIP解离并被激活。细胞通过启动UPR抑制mRNA转录,增强内质网的折叠能力,并激活内质网相关降解途径(endoplasmic reticulum associated degradation,ERAD)以恢复内质网稳态平衡[13]。在慢性或过度的内质网应激下,内质网的正常功能无法恢复,最终导致细胞功能障碍和细胞凋亡[14]。
本研究通过免疫荧光共定位明确Prdx4定位于颗粒细胞内质网,并论证了下调Prdx4蛋白表达对小鼠颗粒细胞凋亡的影响及相关机制。据报道,Prdx4协同内质网氧化还原酶1(endoplasmic reticulum oxidoreductin 1,ERO1)可促进蛋白质二硫键形成[15]。ERO1是内质网中主要的二硫键合成酶,在氧化折叠过程中产生H2O2。而Prdx4在介导二硫键形成的同时可消除ERO1产生的H2O2[16],从而使蛋白质氧化折叠更高效,以维持内质网氧化还原稳态。本研究发现Prdx4蛋白表达水平下调引起颗粒细胞凋亡水平增加,说明Prdx4在调节颗粒细胞凋亡中起着重要作用。
在内质网应激状态下,高度分泌的细胞如胰腺β细胞、浆细胞的内质网显著扩张以代偿大量的蛋白质合成及运输。而在持续的内质网应激刺激下,内质网结构和功能进行性损伤,细胞功能不可逆地破坏,最后导致细胞凋亡。本研究中透射电镜显示下调Prdx4蛋白表达的小鼠颗粒细胞内质网显著扩张。随后,检测内质网应激标志物BIP、UPR通路中的ATF6、UPR信号通路下游的关键转录因子ATF4以及CHOP的表达,结果表明下调Prdx4蛋白表达后的小鼠颗粒细胞BIP、ATF4和CHOP显著升高。有研究表明,当持续的内质网应激无法缓解时,PERK磷酸化真核翻译启动因子2α(eukaryotic translation initiation factor 2α,eIF2α),从而抑制蛋白质翻译但促进ATF4的翻译[17]。ATF4可促进凋亡转录因子CHOP转录,CHOP通过线粒体依赖性途径、死亡受体途径及诱导ROS生成等其他方式促进细胞凋亡,在内质网应激介导的细胞凋亡中起着重要作用[2]。有研究发现上调CHOP表达可以诱导胰岛β细胞凋亡[18],而下调CHOP表达可以抑制内质网应激诱导的心脏成纤维细胞凋亡[19]。CHOP通过下调抗凋亡蛋白、上调促凋亡蛋白促进细胞凋亡,促凋亡蛋白又可以诱导细胞色素c的释放并启动caspase级联反应[20],加速凋亡。本研究证实内质网应激条件下,抗凋亡蛋白Bcl-2表达降低,促凋亡蛋白Bax和cleaved caspase 3表达水平升高,进一步验证了Prdx4表达下降引起持续的内质网应激反应,最终导致颗粒细胞凋亡。
在卵泡发育、卵母细胞成熟过程中经历着复杂的细胞、基因及遗传变化,需要颗粒细胞分泌大量细胞因子和生长因子。这些细胞因子及生长因子需要在内质网中合成、折叠及成熟,并运输到适当位置。因而维持颗粒细胞内质网稳态对其分泌功能尤为重要。有研究表明,Prdx4缺乏导致B细胞向浆细胞分化过程中错误折叠的IgM蓄积[21]。Prdx4与胰岛素原正确的氧化折叠有关[7],并且Prdx4失活会加剧淀粉样蛋白β寡聚体诱导的神经元细胞的内质网应激[22],进一步明确了Prdx4在维持蛋白质稳态中的作用。同时在哺乳动物细胞中存在其他促进蛋白质氧化折叠及抗氧化的基因,如谷胱甘肽过氧化物酶7/8(glutathione peroxidase 7/8,Gpx7/8)及维生素K环氧化物还原酶(vitamin K epoxide reductase,VKOR)等[23]。笔者推测Prdx4保护颗粒细胞的机制主要是通过抑制内质网应激导致的颗粒细胞凋亡,且此作用无法通过颗粒细胞中的ERO1和其他抗氧化酶代偿。
综上所述,本研究发现下调Prdx4蛋白表达可引起颗粒细胞内质网应激,从而促进颗粒细胞凋亡,表明Prdx4作为内质网抗氧化剂,可通过维持蛋白质稳态,下调内质网应激水平,从而抑制颗粒细胞凋亡。Prdx4维持内质网蛋白质稳态的具体机制还需要进一步深入研究。
