藤茶及二氢杨梅素的生物学功能研究进展
2022-03-07罗非君丁锦屏
罗非君,丁锦屏
(1.中南林业科技大学 食品科学与工程学院,湖南 长沙 410004;2.中南林业科技大学 稻谷及副产物深加工国家工程实验室,湖南 长沙 410004)
藤茶是指葡萄科(Vitaceae)蛇葡萄属(Ampelopsis michx.)显齿蛇葡萄(Ampelopsis rossedenttata)植物上生长的嫩茎叶经加工而成的茶制品,俗称为甜茶藤、红五爪金龙、长寿藤、大齿折葡萄等,主要产于我国湖南、湖北、广东、广西、江西等山区较多的省份。分析藤茶的主要活性成分发现,其中含量最高的为黄酮类物质,其中最主要的成分为二氢杨梅素(dihydromyricetin,DMY)。现阶段国内外学者主要致力于研究藤茶及DMY生理功能及其分子机理,为特医食品及保健食品的开发以及临床应用提供理论基础[1-3]。文献[4-6]综述了藤茶及其主要活性成分DMY的研究进展,这些文献主要从藤茶成分的功效、藤茶应用以及DMY的提取工艺等方面展开,但对其生理功能及其分子机理的研究尚处于起步阶段。作者对藤茶和DMY的抗癌、抗氧化、抗炎和抑菌作用、降血压和血糖、降血脂和保护心血管作用等生物学功能及其分子机理的研究进行了系统综述。
1 抗癌作用
癌症在中国已逐渐成为各类疾病的致死率之首,中国平均每天有7 700多名患者死于癌症,即每分钟就有5人死于癌症。虽然分子靶向药物和癌症免疫治疗等新的治疗手段不断出现,但治疗费用昂贵,所以采用膳食调理来预防和抗癌的方式更易被人们接受。至今已经有许多人通过实验证明了藤茶及其主要活性成分具有广泛抗癌及抗肿瘤的作用。藤茶及DMY能抑制乳腺癌细胞增殖和诱导癌细胞凋亡,周等[7]发现DMY可以抑制体外培养的乳腺癌细胞的增殖和侵袭转移,同时促进癌细胞凋亡。DMY可通过调控解整合素金属蛋白酶17(recombinant a disintegrin and metalloprotease 17,ADAM17)、蛋白质酪氨酸激酶(Janus kinase 2,JAK2) 和信号传导及转录激活因子3(signal transducers and activators of transcription,STAT 3)的表达来有效抑制乳腺癌细胞MCF-7增殖并促进其凋亡[8]。还有研究发现,DMY能诱导人乳腺癌细胞MCF-7中抗癌基因十号染色体上缺失的张力蛋白同源物磷酸酯酶基因(phosphatase and tensin homolog deleted on chromosome ten,PTEN)启动子的去甲基化,提高PTEN的表达来部分抑制癌细胞增殖和迁移;DMY通过B淋巴细胞瘤-2/哺乳动物雷帕霉素靶蛋白(B-cell lymphoma-2/the mammalian target of rapamycin,Bcl-2/mTOR)信号途径来抑制乳腺癌细胞增殖、迁移和侵袭[9-10]。藤茶及DMY能降低卵巢癌治疗的耐药性,研究显示,DMY呈剂量依赖性方式下调凋亡抑制因子(inhibitor of apoptosis proteins,IAP)家族中的生存素蛋白表达,诱导卵巢癌细胞凋亡并激活相关信号途径[11],减少肿瘤细胞的耐药性,因此DMY可用于作肿瘤治疗过程的辅助药物。亚硝酸盐是一种致癌物,研究证明藤茶提取物及DMY能抑制PC-3前列腺癌细胞增殖、降低致癌物质亚硝酸盐的生成、减少其代谢产物N-亚硝胺的形成[12]。还有研究表明[13],藤茶提取物可以通过抑制磷脂酰肌醇3-激酶/蛋白激酶B(phosphatidylinositol 3-kinase/protein kinase B,PI3K/Akt)细胞信号通路来下调Bcl-2蛋白表达,从而促进人前列腺癌细胞LNCaP的细胞凋亡。藤茶总黄酮也能通过激活PI3K/Akt/p53的凋亡通路,从而抑制Bcl-2表达和提高Bcl-2相关X蛋白(Bcl-2-associated X protein,Bax)的表达,促进肝癌细胞凋亡[14]。奥沙利铂(Oxaplatin,OXA)是临床上一种结直肠癌(colorectal cancer,CRC)常用治疗药物,WANG等[15]发现DMY可以增强对OXA的药物敏感性,DMY与OXA存在着协同效应,可以增强OXA的抗癌作用。DMY可能通过上调Caveolin-1的表达抑制人胃癌腹腔移植瘤的生长[16]。DMY对非小细胞肺 癌 (non-small cell lung cancer,NSCLC)细 胞(A549和H1975)具有选择性细胞毒作用,但对正常肺成纤维细胞WI-38不具有细胞毒作用[17]。绒毛膜癌是一种恶性滋养细胞肿瘤,研究发现DMY以时间和剂量依赖性方式显著抑制人绒毛膜癌细胞JAr细胞活性,增加了Bax蛋白表达水平,同时抑制Bcl-2和含半胱氨酸的天冬氨酸蛋白水解酶3(cysteinyl aspartate specific proteinase 3,pro-Caspase-3)的蛋白质表达水平,该研究提示DMY是通过线粒体依赖的方式介导的人绒毛膜癌JAr细胞凋亡[18]。DMY除了调控Bax/Bcl-2表达失衡量和促使癌细胞凋亡外,还可能通过调控P13K/Akt信号通路来抑制人绒毛膜癌JAr细胞基质金属蛋白酶MMP-2基因表达,阻滞该细胞的侵袭和转移[19]。迄今为止,很多研究表明藤茶及其提取物在抑制致癌物的形成、促癌细胞凋亡、抑制癌细胞的转移、减少癌细胞的耐药性和增加抗癌药物敏感性等方面均起作用,但其作用的分子靶点尚处于起步阶段,见表1。在今后的研究中,需要通过基因芯片等技术系统地确定藤茶及其提取物调控的基因和相关的信号通路,阐明其抗癌作用的分子机理。藤茶及其提取物作为一种有效的天然抗癌活性成分,无毒性作用和副作用,但DMY临床试验疗效证据尚不确定,需要进一步研究。当前需要开展临床方面的实验研究,评估在临床应用的有效性,是否跟细胞实验存在差异,是否以相同的分子机理抑制癌症;在功能性食品开发方面,需要进行动物实验,进一步确定防癌、抗癌的功效,也需要通过流行病学的研究,确定长期饮用藤茶与癌症发病率之间的关系等。
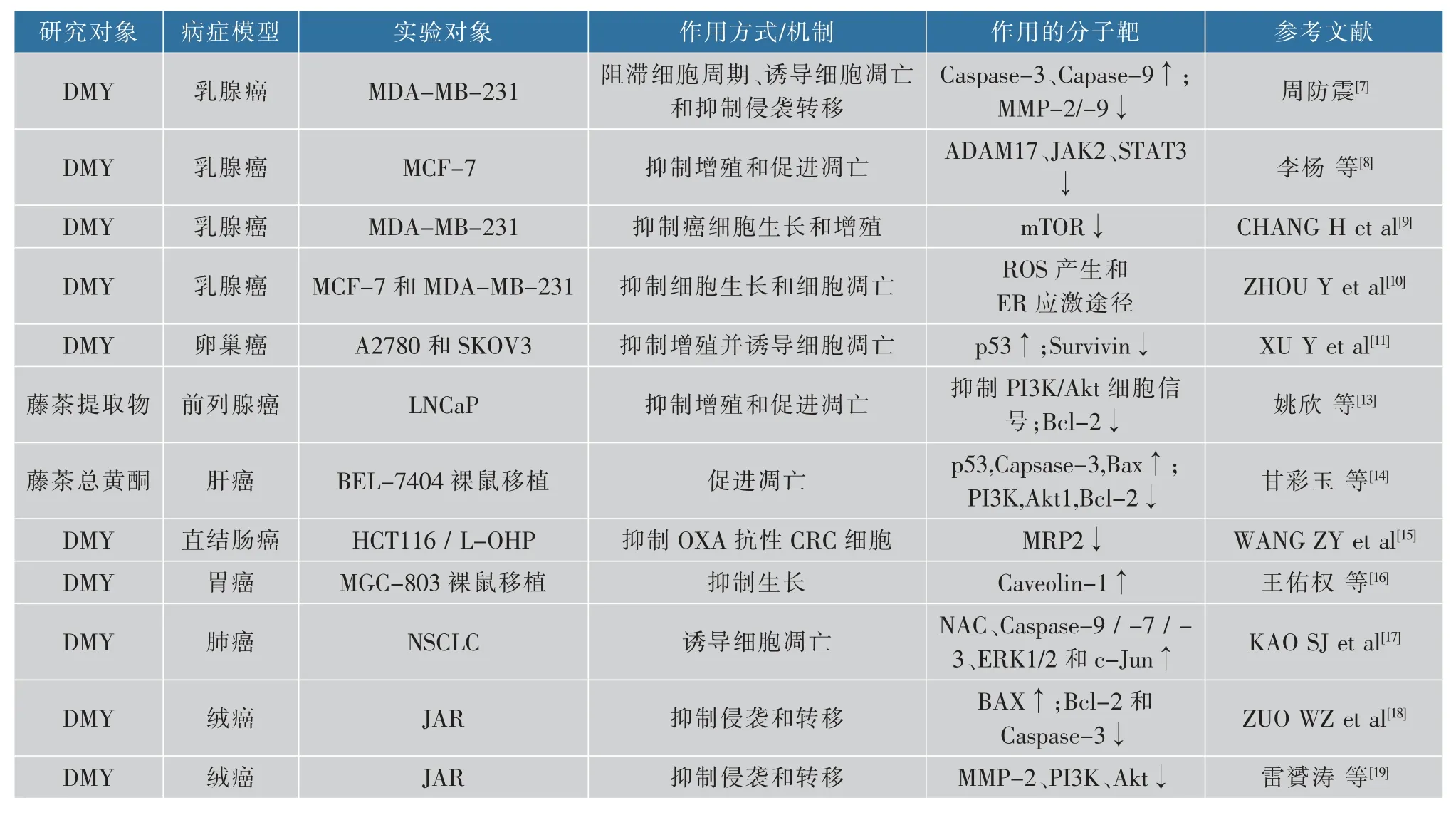
表1 藤茶及其主要活性成分抗癌作用机理研究文献Table 1 References in mechanism of anti-cancer effects of vine tea and its main extracts
2 抗氧化作用
抗氧化是抗氧化自由基的简称,机体因为呼吸作用(氧化反应)、外界污染、放射线照射等因素能在体内产生大量的自由基,而癌症、衰老或其他疾病大都与过量自由基的产生有关联。有研究发现藤茶及其主要活性成分均具有抗氧化作用[20-22]。陈丽等[23]研究表明,藤茶提取物可能是通过调节肠道微生物和自由基平衡来阻止小鼠体内脂质的过氧化从而缓解小鼠的腹泻并提高超氧化物歧化酶(superoxide dismutase,SOD)、谷胱甘肽过氧化物酶(glutathione peroxidase,GSH-Px)和过氧化氢酶(catalase,CAT)的活性。罗淑萍等[24]研究了藤茶总黄酮对大鼠脑缺血再灌损伤的保护作用,与对照组相比,藤茶总黄酮组可以使心脏梗死面积减少,且可以提高大脑超氧化物歧化酶(superoxide dismutase,SOD)活性。丙二醛(malondialdehyde,MDA)是机体氧化损伤的重要指标,藤茶总黄酮能显著抑制大鼠的MDA,说明藤茶总黄酮有较好的抗氧化功能。欧贤红等[25]的研究发现,藤茶的主要提取物清除·OH的效果依次是杨梅素>DMY>藤茶总黄酮。徐新等[26]发现藤茶提取物呈一定的剂量依赖性对DPPH自由基、羟基自由基以及超氧阴离子自由基有显著的清除或抑制作用。李雪晖[27]分析了DMY对O2-·、·OH、ROO·的清除效果,在一定质量浓度范围内,DMY的清除能力和质量浓度成正比。清除程度最高分别为90%、64%以及88%,由此看出,DMY本身具有比较强的抗氧化能力。有研究者将DMY添加在曲奇中[28],添加后虽然会对其颜色和挥发性有一些影响,但是不会对质地有影响。感官评估表明,添加DMY对曲奇没有明显影响,反而可以减弱脂质和蛋白质氧化的作用,并降低了曲奇中羧甲基赖氨酸和荧光高级糖基化终产物的含量,因此DMY可作为一种潜在的功能性烘焙添加剂。目前关于藤茶及其提取物抗氧化作用的研究很少涉及其作用机理,以后的研究需要更多地注重其分子机制。
3 抑菌与抗炎作用
抑菌作用主要是指抑制或杀灭细胞,抗炎作用包括抗感染性炎症和抗非感染性炎症,抗菌药能够通过抑制或杀灭引发炎症的细菌,从而消除感染性炎症,达到治愈疾病的目的。已有研究证明藤茶及其主要活性成分都有一定的抑菌效果[29]。耐甲氧西林金葡菌(Methicillin-resistant Staphylococcus aureus,MRSA)是医院及社区感染的主要病原菌之一,其具有广谱耐药性,对β-内酰胺类和头孢类抗生素均耐药,对氨基糖苷类、大环内酯类、四环素类、氟喹诺酮类、磺胺类、利福平均有不同程度的耐药性。颜欣[30]通过研究Toll样受体2(Toll-like receptor 2,TLR 2)信号通路探究DMY对MRSA菌血症的影响。结果表明,DMY可在体外抑杀MRSA;在体内有可能是调节TLR2信号通路的免疫细胞因子的表达和调控机体中的白细胞数量,从而增强机体免疫应答功能,并且增强了机体对细菌的清除及抑杀能力,通过这些方式DMY在体内防治MRSA菌血症的感染。DMY与苯甲酸钠的抑菌作用对比研究发现,DMY对大肠杆菌、副伤寒沙门氏菌、白色念珠菌3种菌的抑菌和杀菌效果更好,而对金黄色葡萄球菌、绿脓杆菌、枯草芽孢杆菌3种菌的抑菌和杀菌效果与苯甲酸钠相当[31]。现有研究表明,藤茶对炎症有一定的缓解作用[32-33]。在脂多糖(lipopolysaccharide,LPS)诱导的小鼠炎症模型和RAW264.7巨噬细胞炎症模型中,Hou[34]发现使用高浓度的DMY能够明显减少小鼠血清中肿瘤坏死因子-α(tumor necrosis factorα,TNF-α)、白细胞介素6(interleukin-6,IL-6)和白细胞介素-1β这些促炎因子的表达,并且使小鼠血清抗炎因子白细胞介素10的表达水平提高,表明DMY可以通过减少炎症介质的释放和相关酶的蛋白质表达来抗炎,且可能是通过抑制p38和JNK的磷酸化以及NF-κB的活化来实现的。Wang等[35]研究表明,DMY能抑制RAW264.7细胞IKKα、IKKβ的磷酸化,阻止转录因子NF-κB进入细胞核,抑制前体炎症因子一氧化氮合酶(induced nitrogen monoxide synthase,iNOS)表达,进而减少NO的产生;同时,DMY也能抑制炎症因子环氧合酶-2(cyclooxygenase-2,COX-2)表达。在角叉菜胶诱导的大鼠爪水肿炎症模型中,DMY能通过抑制炎症因子的表达,明显改善大鼠爪的水肿。DMY对溃疡性结肠炎(ulcerative colitis,UC)也有一定的缓解和保护作用,DMY可以调整Treg/Th17细胞状态,改善肠道微环境,并且可以减少局部结肠组织中MMP-9的表达以及ECM的降解[36]。除此以外,藤茶可以通过抑制IRAK1/TRAF6/NFκB介导的炎症信号通路改善UC[37]。加热煎煮减压浓缩得到的0.5 g/mL的藤茶水提取物对氨水急性咽炎大鼠模型进行灌胃保护,实验结果表明,藤茶提取物可通过降低TNF-α、IL-1β、IL-6、IFN-γ的表达和提高IL-10的表达量来缓解炎症[38]。用完全弗氏佐剂诱导建立类风湿性关节炎(rheumatoid arthritis,RA)模型,发现DMY能通过激活Nrf 2途径来抑制促炎性细胞因子的表达来改善RA症状[39]。DMY也可以通过抑制NF-κB信号传导来缓解胶原诱导关节炎(collagen-induced arthritis,CIA)大鼠的炎症并减轻IL-1β诱导的成纤维细胞样滑膜细胞(FLS)活性[40]。微胶质介导的神经炎症被认为是阿尔茨海默病(Alzheimer’s disease,AD)、帕金森病(Parkinson’s disease,PD)等神经退行性疾病的病因之一。有研究表明,藤茶的抗炎作用主要是通过激活NF-κB和JAK2/STAT3信号通路来减轻小胶质细胞中LPS刺激的炎症反应[41]。急性胰腺炎(acute pancreatitis,AP)是胃肠道常见的炎症性疾病,DMY可以同时抑制雨蛙肽(caerlin)诱导的TRAF3-p38信号通路的激活作用和AP反应[42]。在藤茶及其提取物抗炎研究领域,目前可以确定其可能抑制炎症因子的表达,但其如何干预炎症相关的信号通路和转录因子尚需要进一步研究。慢性炎症与肥胖、癌症的发生等相关,推测其可能对炎症引起的肥胖、癌症或其他疾病有一定的缓解和保护作用,见表2。
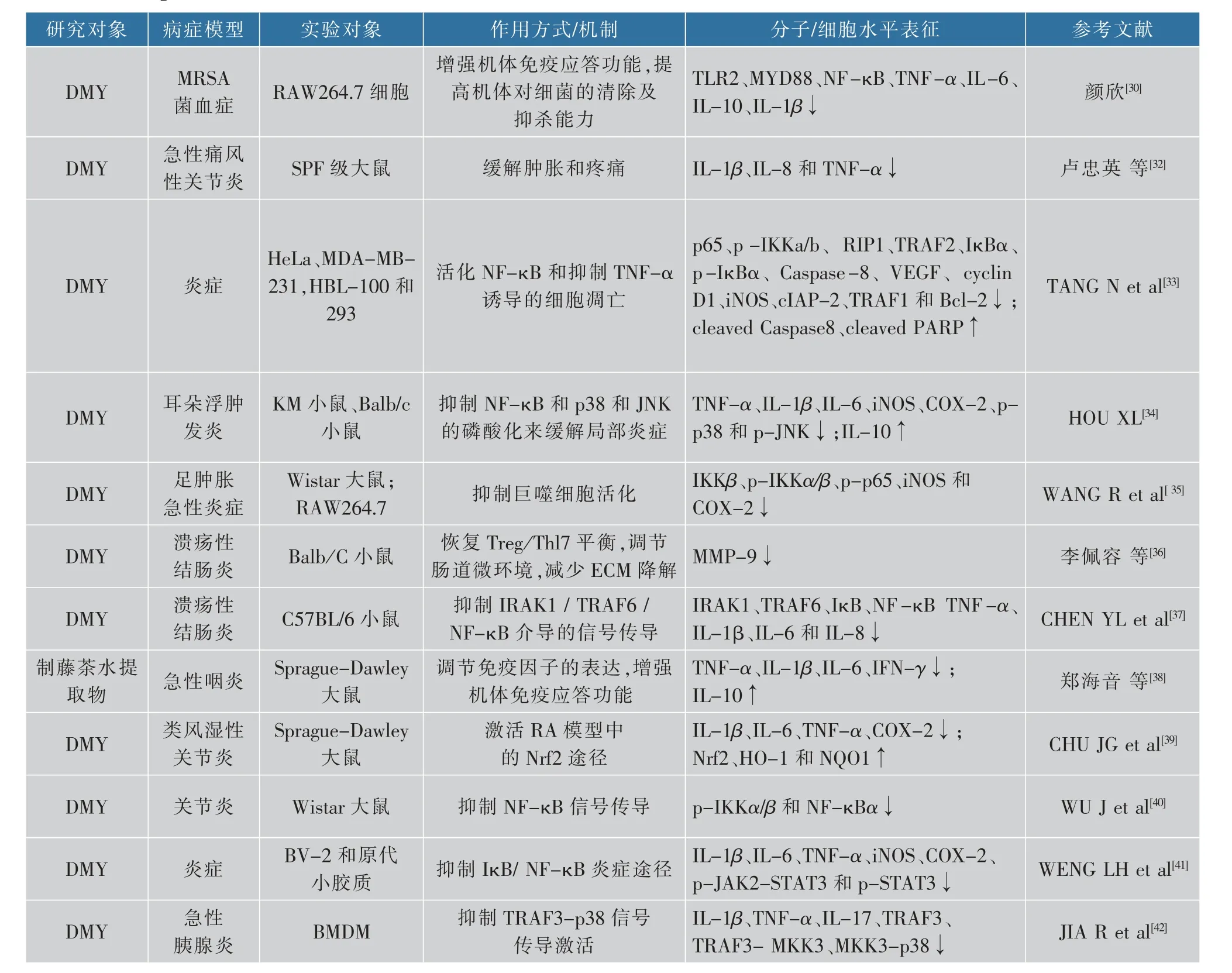
表2 藤茶及其主要活性成分抑菌及抗炎作用机理研究文献Table 2 References in mechanism of anti-bacteria and anti-inflammatory effects of vine tea and its main bioactive components
4 降血糖与降脂作用
高血糖和高血脂是两种完全不同的疾病。当胆固醇和甘油三醇大于正常值会患高脂血症,而当血糖大于5.6 mmol/L时便是高血糖,大于7.0 mmol/L时会患糖尿病。高血糖往往会伴随血脂代谢紊乱,而血脂过高时能引起心脑血管相关疾病。现有研究表明,藤茶及其主要活性成分具有降血脂与血糖的作用[43-44]。 其中吴瑛等[45-49]通过建立2型糖尿病(type 2 diabetes mellitus,T2DM)大鼠模型来证明藤茶提取物及DMY可有效缓解糖尿病大鼠的症状。为了探讨藤茶黄酮对生物生成细胞内源性胆固醇和对高脂血症大鼠的降血脂作用效果,ZUO等[50]使用乙醇提取得到藤茶黄酮(TFAG),采用两性霉素B-细胞模型方法来测定藤茶黄酮对细胞内胆固醇合成的影响。研究表明,藤茶黄酮可以显著控制细胞内的胆固醇合成,并减少大鼠血清以及心、肝组织中的总胆固醇和甘油三酯的含量,调控肝脏中的SOD、CAT、GSH和MDA的含量,表明藤茶黄酮有较好的抗氧化和降血脂的效果,并且可以有效抑制肝脏脂肪的变性。此外,DMY还可以通过改善葡萄糖相关代谢,同时降低喂食高脂饮食(high fat diet,HFD)的大鼠的脂质水平,来治疗T2DM胰岛素抵抗作用和肝脏脂肪变性[51]。刘蕾等[52]的研究发现,DMY通过抑制MEK/ERK信号通路,降低PPARγ273位点丝氨酸磷酸化,促进脂肪细胞糖摄取和脂联素分泌,提高细胞胰岛素敏感性。而Ran等[53]通过70名受试者的临床试验证明,补充藤茶1个月可明显改善成人T2DM患者的血糖水平和肾功能指标。DMY能够减弱线粒体合成酶(FIFO-ATP)的活性,使得AMP:ATP的比值增大,进一步激活AMPK通路,由于SIRT3、PGC-1α表达上调,使得骨骼肌细胞的自噬作用增强,骨骼肌胰岛素敏感性提高,改善骨骼肌胰岛素抵抗作用[54]。脂肪肝是因为肝细胞内脂肪堆积过多而导致的一种常见的病变,脂肪肝发病率一直不断增加,现已经成为仅落后于病毒性肝病的第二大肝病,但这种肝病是可逆转的。研究也发现,DMY干预可部分恢复非酒精性脂肪肝病(non-alcoholic fatty liver disease,NAFLD)小鼠肝脏生物钟相关基因表达节律紊乱,进而改善肝脏脂代谢[55]。近期有研究表明[56],藤茶提取物可以降低血清胆固醇和甘油三酸酯,减少肝脂质的积累,通过激活AMPKα磷酸化,提高PPARα、细胞色素P450等的表达量;并通过降低SREBP1和FAS的表达,激活HO-1、NQO1和Nrf2的表达;同时还降低TBARS的表达来防止氧化应激反应的发生。此外,藤茶还能增加肠道粘连蛋白ZO-1的表达,防止细菌入血;下调肠道菌Firmicutes/Bacteroidetes的比率,改善肠道微生物的组成。以上结果表明,藤茶可以通过平衡脂肪酸的氧化和脂肪生成,抑制肝脏的氧化应激反应并调节肠道微生物组成来预防NAFLD。DMY可以上调AMPK的磷酸化,增加AMPK的活性,同时增加SIRT1的表达,从而减少HepG2细胞脂肪蓄积,还可以下调HepG2中SREBP-1c、FAS和ACC这些脂质合成相关基因,减少细胞内脂质合成,缓解肝细胞脂肪变性症状[57]。DMY也能缓解十八烯酸诱导的肝脏脂肪变性,并使得总甘油三酯和总胆固醇含量下降;同时DMY还能极显著减少脂肪代谢关键调控因子PPARγ的表达量、增加AMPK在细胞中的磷酸化作用[58]。此外,DMY可以减少肝脏脂肪的蓄积可能与DMY可以调节肝脏线粒体融合与分裂有关[59],还有研究证明DMY可以对巨噬细胞泡沫细胞转化中脂质积累起阻碍作用[60]。目前已有的研究确定了藤茶及其提取物具有降脂的功效,分子机理方面也不断取得进展,已有的研究确定其作用的多个分子靶,但其对相关信号通路的研究尚处于起步阶段,其降脂的分子机理尚不够完善。除此之外,越来越多的研究发现,降脂与机体肠道微生物菌群的改变相关。藤茶及其提取物是否能调节肠道微生物菌群,是否能通过调控肠道微生物菌群参与降脂作用还没有相关实验证据,见表3。
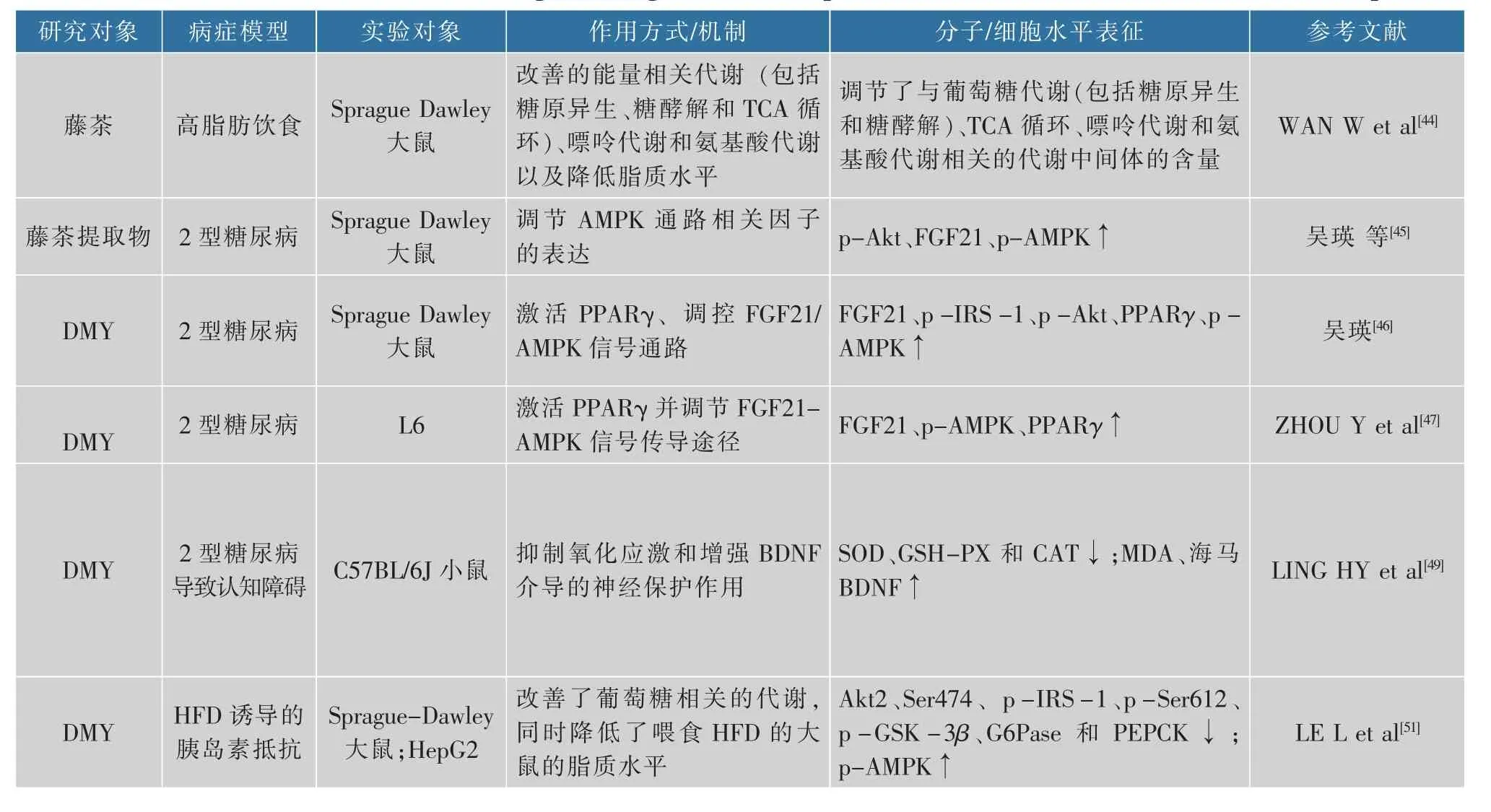
表3 藤茶及其主要活性成分降血糖血脂作用机理研究文献Table 3 References in mechanism of lowering blood sugar and blood lipid effects of vine tea and its main bioactive components
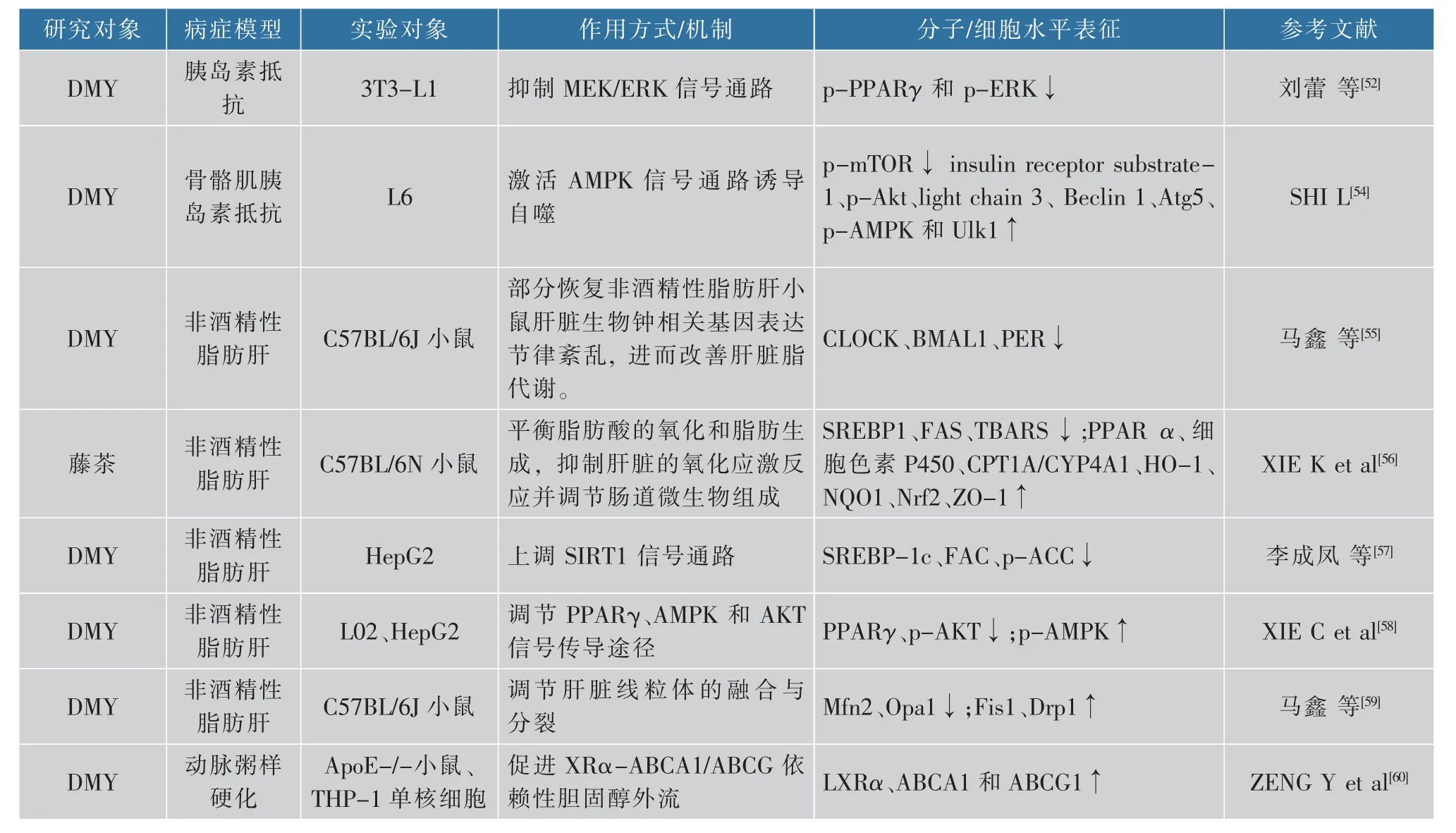
续表
5 降压与对心脑血管病的防治作用
随着时代的发展,人们生活水平不断提高,在饮食方面人们越来越倾向于摄入脂肪含量高的食物,这也导致很多人血压高于正常值。虽然高血压是一种十分常见的慢性病,却与心脑血管病有着密切的联系,人们应该关注自身血压的变化,出现异常时应及时采取措施进行防治。现有研究表明,藤茶及其提取物可以预防和改善动脉粥样硬化,对心血管有一定的保护作用[61-62]。秦力等[63]研究发现,DMY通过减少人脐带静脉内皮细胞(human umbilical vein endothelial cells,HUVECs)细胞线粒体酶复合物(FIFO-ATP)合成酶活力来减少ATP的含量并使其AMPK活性上升,对HUVECs细胞起到保护作用。DMY也可以缓解过氧化氢诱导的内皮细胞损伤或凋亡,抑制动脉粥样硬化的起始环节——内皮细胞的损伤来防治或减慢动脉粥样硬化的病变过程[64]。DMY能增进高脂喂养ApoE-/-小鼠体内胆固醇逆向转运(reverse cholesterol transport,RCT),有助于DMY抗动脉粥样硬化和调节血脂,为DMY在心血管疾病方面的应用提供了新的理论基础[65]。miR-21介导的DDAH1/DMA/NO信号通路在TNF-α诱导的内皮功能障碍中起重要作用,DMY可以通过阻碍该通路的激活与表达来缓解TNF-α诱导的内皮功能障碍[66]。已有研究表明,棕榈酸可以诱导血管内皮细胞发生焦亡作用,而DMY可阻滞棕榈酸诱导的血管内皮细胞NLRP3活化及IL-1β分泌,说明DMY可以预防和缓解由棕榈酸引起的血管内皮细胞焦亡[67]。DMY通过过度激发的MaxiK和KATP改善实验性败血症中大鼠主动脉的收缩性受损[68],DMY还可以明显抑制结扎诱导的颈动脉新内膜形成。实验结果表明,DMY增加孤立核受体TR3在平滑肌细胞(smooth muscle cells,SMC)和颈动脉中的表达,并且证明DMY促进SMC分化,通过调节TR3的表达抑制SMC增殖和迁移[69]。DMY可以增强大鼠的抗氧化能力、抑制体内和体外的细胞凋亡,在I/R(ischemia-reperfusion,I/R)损伤中有保护心脏的潜力[70],其可能对急性心肌梗死(acute myocardial infarction,AMI)治疗的临床研究提供新的方向。另外还有研究表明,DMY可以减弱LPS诱导的心肌细胞损伤,该作用机制可能与DMY抑制TLR-4活化和随后调节NF-κB的相关下游信号传导途径有关[71]。藤茶及其提取物对心血管疾病的保护作用,也与其抗氧化和抗炎功能密切相关,见表4。
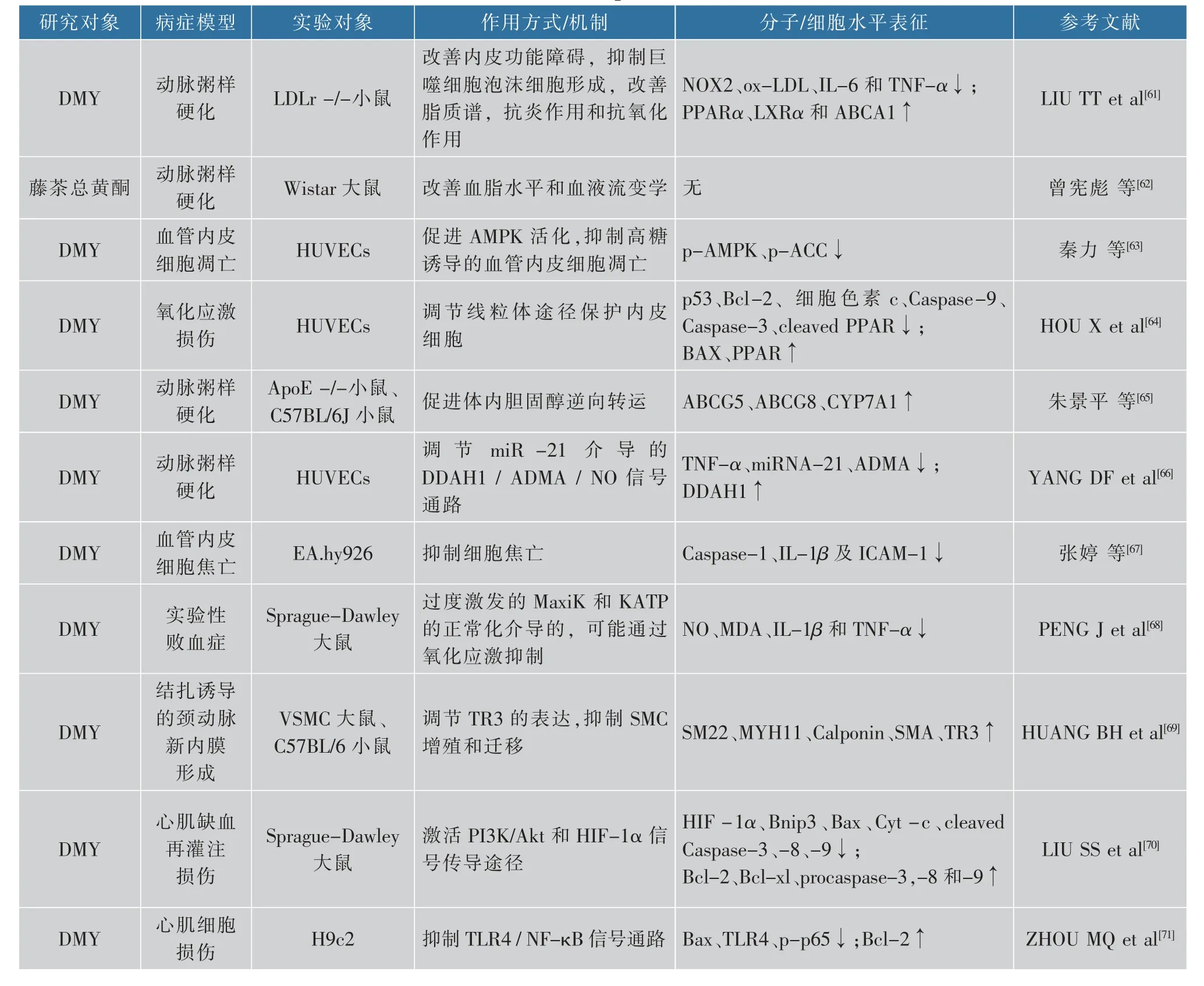
表4 藤茶及其主要活性成分降压与对心脑血管病的防治作用机理研究文献Table 4 References in mechanism of antihypertensive and prevention and treatment of cardiovascular and cerebrovascular diseases effects of vine tea and its main bioactive components
6 其他作用
藤茶及其提取物的生理功能十分多样,在研究中还发现,DMY能够显著改变肠道微生物群的丰富性和多样性,并调节肠道微生物群的组成[72]。另外,将藤茶提取物混入鸡的饲料中可以增强鸡的消化和吸收能力,促进鸡的生长[73]。有研究表明,DMY还能通过Keap-1/Nrf2信号通路来调节p62表达和肝细胞的自噬,对酒精性肝病导致的损伤具有保护作用[74]。LIU等[75]研究发现,DMY可以通过预防氧化应激、细胞凋亡和炎症来改善铅(Pb)诱导的认知功能障碍。FENG等[76]研究表明,DMY可以降低APP/PS1小鼠的海马和皮层中的活化小胶质的数量来显著改善记忆和认知缺陷,并可以通过抑制NLRP3炎症组分基底微胶质介导的神经炎症来阻滞AD的病理进展。此外,还有研究表明,DMY可以通过上调AMPK/SIRT1通路抑制炎症反应和海马细胞凋亡来改善认知功能,在AD中发挥保护作用[77]。在KOU等[78]的研究中发现,脑老化模型大鼠中显示miR-34a表达明显上升,而在DMY处理后miR-34a表达得到显著抑制。DMY还可以抑制D-gal诱导的细胞凋亡并挽救海马组织中神经元自噬受损,说明DMY可以预防和治疗脑老化。也有研究表明,DMY可以调节Akt/GSK-3β信号通路,表明DMY是DA神经元的有效神经保护剂,所以猜测DMY可能是一种潜在的PD的治疗药物[79]。DMY还可以减少骨质流失,并通过RANKL信号传导的多个途径调节破骨细胞的形成和活性。因此,DMY可能成为治疗破骨细胞相关疾病(例如类风湿性关节炎和骨质疏松症)的药物成分[80]。也有研究表明,SIRT3缺乏与退行性骨关节病(osteoarthritis,OA))发展有关,而DMY可通过在软骨细胞中的SIRT3依赖性方式维持线粒体稳态进而发挥抗变性作用[81]。DMY可以增加—OH处理后的间充质干细胞的活力,分析表明DMY可能是细胞移植治疗的有益剂[82]。WANG等[83]还发现,DMY可以抑制丝裂原活化蛋白激酶(mitogenactivated protein kinase,MAPK)信号传导,可能对急性肺损伤(acute lung injury,ALI)具有保护作用。顺铂是抗肿瘤最有效的治疗药物之一,但是顺铂化疗会出现肾毒性,从而限制了顺铂的临床应用。研究表明,DMY可以改善顺铂引起的肾功能损害和结构损伤。此外,DMY显著减弱顺铂诱导的氧化应激,并且在小鼠肾组织中保护免受顺铂诱导的炎症和凋亡细胞死亡[84]。DMY还可以通过逆转PGC-1α/TFAM和PGC-1α/mfn2信号通路介导的线粒体功能障碍缓解肌肉萎缩[85]。也有研究表明,DMY可以通过抑制MMP-2的表达来抑制人视网膜色素上皮细胞(RPE)迁移,说明DMY可能在治疗人类增殖性玻璃体视网膜病变中(proliferative vitreoretinopathy,PVR)有潜在治疗价值[86]。HUANG等[87]证明DMY可以抑制黑素生成,该研究表明,DMY还有可能发展成防止色素沉着的护肤品。肝纤维化是一种慢性肝损害,是慢性肝病发展到肝硬化的过程,目前仍没有确定有效的药物,而张文涛等[88]的研究表明,藤茶总黄酮可以调节转化生长因子-β(transforming growth factor-β,TGF-β)、Smad蛋 白(drosophila mother against decapentaplegic protein,Smad)2、Smad 4、Smad 7、人α平滑肌肌动蛋白(α smooth muscle actin,α-SMA)的表达缓解肝的纤维化病理进程,说明藤茶总黄酮可以通过调控TGFβ1/Smad信号通路来抗肝纤维化,见表5。
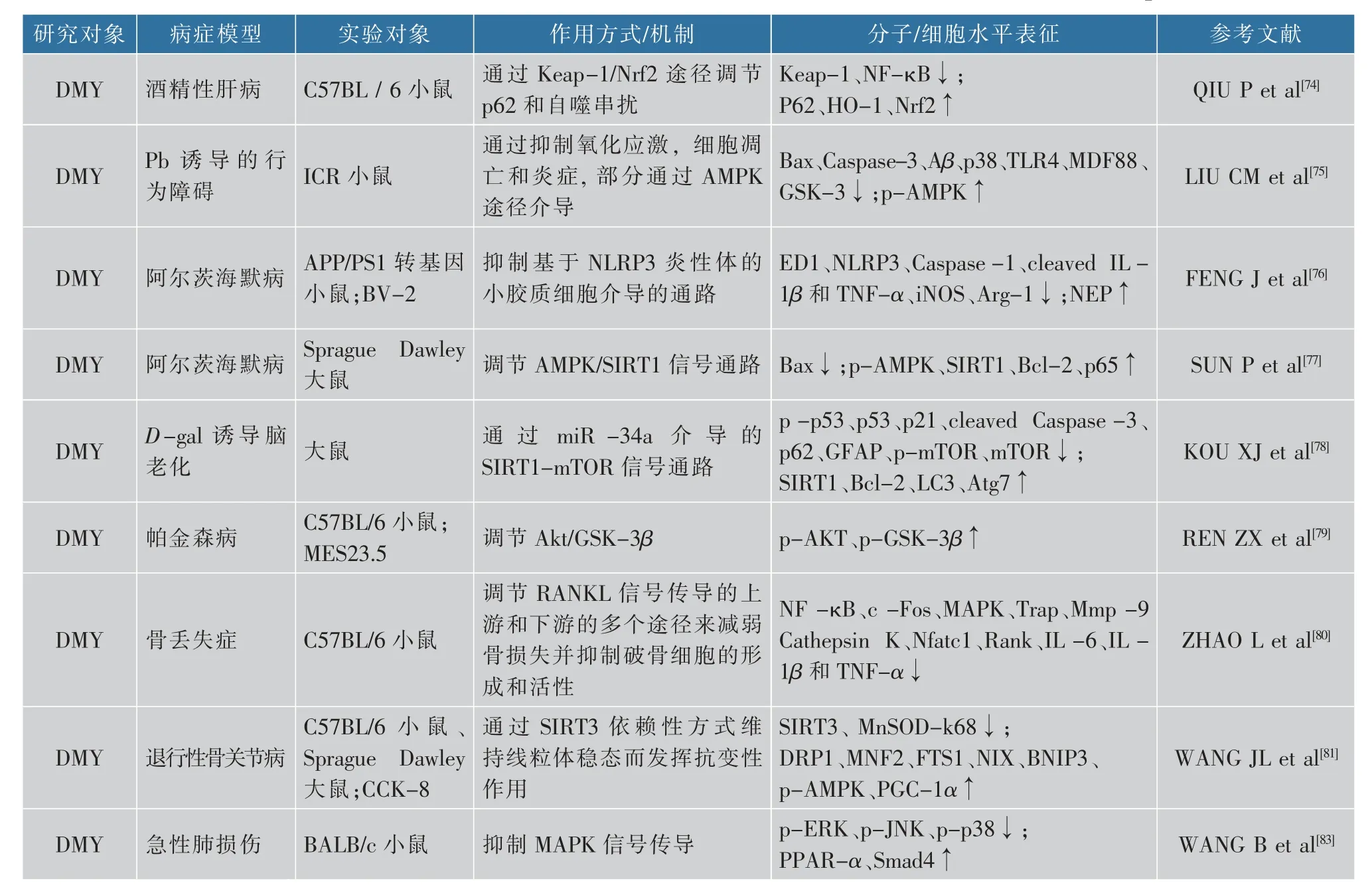
表5 藤茶及其主要活性成分其他作用的机理研究文献Table 5 References in the mechanism of other effects of vine tea and its main bioactive components
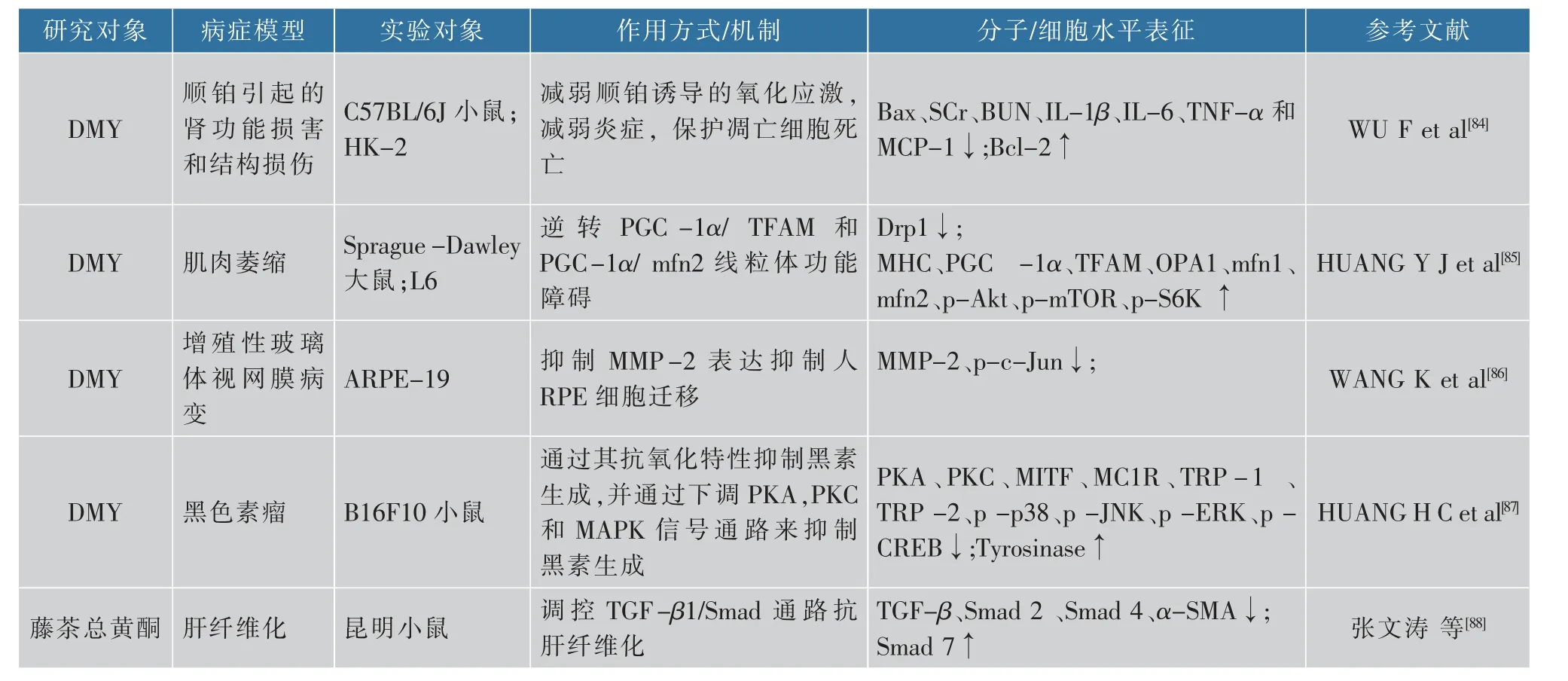
续表
7 展望
综上所述,藤茶及其主要活性成分的抗癌作用、抗氧化作用、抗炎和抑菌作用、降血压和血糖、降血脂和保护心血管等功效都已被证实。此外,藤茶及其主要活性成分在阿尔兹海默症、脑老化、肌肉萎缩等疾病进展中也有一定的保护和缓解作用。这些研究还表明,藤茶及其提取物除了在功能性食品应用上有发展前景之外,还可能在食品添加剂、护肤品等方面有应用的可能性。
总的来说,藤茶及其主要活性成分的研究在分子机理方面有了一定的进展,许多生理功能的相关通路被证实,藤茶及其主要活性成分的研究也取得了一些突破性进展。有文献表明[66,78],藤茶及其主要活性成分可能靶向microRNA调控信号通路来发挥其生理作用。未来的研究方向应该深入研究信号通路之间的联系、重要靶蛋白和靶基因如何调控相关通路以及microRNA如何影响其发挥相应的生理功能等[89-90]。除此以外,还应该探索一些新的调控方式,例如:藤茶活性成分是否能通过调控长链非编码RNA(long non-coding RNA,LncRNA)、肠道微生物菌群来发挥生物学功效。
近年来,关于DMY的药理代谢研究也有了初步的进展。有研究发现[91],大鼠口服DMY后在机体许多组织中都能检测到,且在肠组织中含量最高;同时,机体内DMY含量出现快速降低,其代谢途径可能与脱羟基化、甲基化、葡萄糖醛酸化和硫酸化等相关,DMY代谢物可以通过尿液排泄或直接进入胆道排泄。当前这一领域的研究需要解决的关键问题包括:DMY如何进入细胞?DMY进入细胞后与什么靶蛋白质结合?DMY与靶蛋白质结合后如何调控相关的信号通路?DMY的分子改构是否能增加其成药性和药效性?
目前藤茶活性物质相关的分子机理研究还不够系统、全面,已有的研究尚缺少转录组学、蛋白质组学和代谢组学的研究。另外,目前尚缺少临床试验方面的相关研究。
基于藤茶及其提取物无毒性且具有多种生物学效应,其在功能性食品开发方面有应用价值。
