离心除菌UHT乳贮藏期内乳蛋白组成的变化
2022-03-02孙玥吴华星王荣春段雪梅李琳赵广生马莺李启明
孙玥,吴华星,王荣春,段雪梅,李琳,赵广生,马莺,李启明
(1.哈尔滨工业大学 化工与化学学院,哈尔滨 150090;2.新希望乳业股份有限公司,成都 610063;3.乳品营养与功能四川省重点实验室,成都 610000;4.杭州新希望双峰乳业有限公司,杭州 310000)
0 引言
超高温瞬时杀菌(ultra-high temperature,UHT)是牛乳在135~150℃下加热3~5 s,然后无菌包装于微量透气的容器中,生产出来的产品称UHT乳[1]。UHT可杀死细菌、体细胞、酶、多数芽孢,但不能杀死全部的耐热芽孢菌。UHT乳在室温下的保质期是6个月[2]。耐热性内源酶和耐热芽孢产生的酶,微生物及体细胞自溶所释放的胞内酶在UHT乳贮藏期间保持活性严重地影响了UHT乳的质量[2]。离心除菌技术(bactofugation)是通过超速离心除去原料乳中所有的体细胞和90%以上的细菌,降低原料乳的微生物和体细胞来改善UHT乳的质量[3]。
在同一乳品厂按照相同的工艺生产常规UHT乳(N-UHT)和离心预除菌UHT乳(C-UHT),研究热处理和贮藏期N-UHT乳和C-UHT乳的蛋白质组成和结构的变化,探讨离心预除菌工艺对改善UHT乳贮藏期质量品质的作用。
1 材料与方法
1.1 材料与设备
牛乳来源于新希望自有牧场,原乳细菌总数小于2×105cfu/mL,体细胞小于5×105个。K9840凯氏定氮仪,济南海能仪器股份有限公司;L8800全自动氨基酸分析仪,日本日立公司;LC-16高效液相色谱仪,岛津企业管理(中国)有限公司。
1.2 方法
在同一乳品厂按照相同的工艺分别生产了两批常 规UHT(N-UHT)乳 和 离 心 除 菌UHT乳(CUHT),每一批的原料乳相同。N-UHT乳杀菌工业参数为135℃/4 s;C-UHT工艺参数为:55~60℃原料乳在7 200 g下离心,回流速率为300~400 kg/h,经检测可去除所有的体细胞和91%的细菌,离心后的原料乳经过135℃/4 s杀菌。UHT乳在常温下贮藏6个月,每间隔1个月取样。参照食品安全国家标准(GB/T5009.5-2016)凯氏定氮法测定样品的总蛋白(TN)质量浓度;参照IDF020-4-2001测定非蛋白氮(NPN)质量浓度。参照IDF20B:1993凯氏定氮法,采用醋酸沉淀样品中的酪蛋白,测定非酪蛋白(NCN)质量浓度。乳中每种蛋白质的质量浓度在测得的NPN和NCN最终乘上系数6.38,根据乳清蛋白氮(WPN)=NCN-NPN;酪蛋白氮(CN-N)=TN-NCN计算氮质量浓度。采用反向高效液相色谱法测定样品中αs1-酪蛋白(αs1-CN)、β-酪蛋白(β-CN)、κ-酪蛋白(κ-CN)、α-乳白蛋白(α-La)、β-乳球蛋白(β-Lg)质量浓度[4]。采用氨基酸分析仪测定乳样品中游离氨基酸组成和质量浓度[5]。
2 结果与讨论
2.1 热处理和贮藏过程中氮分布的变化
乳中蛋白是由真蛋白和非蛋白氮(NPN)构成,真蛋白是由酪蛋白和乳清蛋白组成,NPN主要是由游离氨基酸、短肽、尿素、肌酐、生物胺等含氮化合物组成[6]。NPN是牛乳代谢产生的,占总氮的约5%,热处理、贮藏等环境因素的改变,乳蛋白氧化、分解能够产生一些游离氨基酸和含氮的小分子化合物,体现在NPN的增加和真蛋白的减少。普通的UHT乳(NUHT)和原料乳离心除菌后的UHT乳(C-UHT)在热处理前后(D0)及贮藏期(M1-M6)内酪蛋白氮(CNN)、乳清蛋白氮(WP-N)和非蛋白氮(NPN)之间的比例发生了改变,如图1所示。

图1 两种UHT乳热处理和贮藏过程中氮分布
加热和贮藏过程中发生了乳蛋白的变性和分解,NPN增加,但是总氮(TN)不会变化。与原乳(RM)比较,热处理后N-UHT-D0中WP-N质量分数由28.1%减少到21.5%,NPN由4.2%增加到5.3%,CNN由67.8%增加到73.3%;C-UHT-D0中WP-N质量分数由28.1%减少到22.8%,NPN由4.2%增加到4.6%,CN-N由67.8%增加到72.7%。乳清蛋白是具有4级结构的球状蛋白,热处理破坏其结构,分子展开,游离巯基暴露出来,通过分子间或分子内二硫键相互作用形成凝聚物,进入胶束相[7]。β-Lg和α-La是乳清蛋白的主要成分,变性β-Lg和α-La与胶束表面的κ-CN通过分子间二硫键作用形成乳清蛋白/κ-CN复合物聚集到酪蛋白胶束表面[8]。热诱导的蛋白降解以及美拉德反应和脱酰胺作用的发生,产生了游离氨基酸、肽、以及一些蛋白降解产物,这些产物以非蛋白氮形式存在[9]。因此,WP-N质量分数降低,NPN质量分数增加。
N-UHT乳和C-UHT乳在贮藏过程中CN-N质量分数变化不显著,两者之间差异不显著(P<0.05)。随着贮藏时间的增加WP-N质量分数逐渐降低,NUHT中WP-N由N-UHT-D0的21.5%降低到17.6%(N-UHT-M6),C-UHT的WP-N由CUHT-D0的22.8%降低到17.2%(C-UHT-M6),两者之间的变化差异不显著;NPN随着贮藏时间的增加而显著增加,从N-UHT-D0的5.3%增加到N-UHTM6的8.7%,C-UHT的NPN从C-UHT-D0的4.6%增加到7.9%(C-UHT-M6),如图1所示。结果表明,在贮藏过程中NPN的增加主要与WP-N有关。热诱导的蛋白质氧化及蛋白质之间的相互作用在贮藏过程中持续发生,蛋白水解、蛋白的氧化降解、美拉德反应等是引起贮藏期间蛋白质量浓度改变的主要原因[10]。原料乳中嗜冷菌分泌的耐热性蛋白酶和内源性纤溶蛋白酶在UHT贮藏过程中持续水解蛋白,造成UHT乳苦味和凝胶形成已经被大量研究所证实[2,11]。C-UHT乳由于原料经过预除菌处理,源于原料中嗜冷菌产生的蛋白酶的量低于N-UHT乳,所以其贮藏过程中NPN增加的程度低于N-UHT乳。非酶脱酰胺是将蛋白质的天冬酰胺和谷氨酰胺残基中的酰胺基水解为天冬氨酸和谷氨酸以及氨水,UHT热处理引起的非酶脱酰胺程度较小,但在贮藏过程中能够发生明显的脱酰胺作用[12],并且是随着贮藏时间的延长而增加[13]。UHT杀菌强度引发美拉德反应,贮藏过程中美拉德反应持续进行生成糠氨酸及褐色色素[14],体现在UHT长期贮藏颜色变褐色。蛋白水解产物在乳中最直观的体现是NPN的增加。
2.2 热处理和贮藏过程中乳蛋白在胶束相和乳清相的分布
天然乳中酪蛋白(αs1-CN、αs2-CN、β-CN、κ-CN)是以胶束形式存在于胶体相中,乳清蛋白(α-La、β-Lg、BSA)以可溶性形式存在于乳清相中,热处理导致乳清蛋白变性,变性乳清蛋白与胶束表面的κ-CN结合形成乳清蛋白/κ-CN复合物进入胶束相,复合物的形成降低了酪蛋白胶束的稳定性,部分酪蛋白解聚进入乳清相[15];贮藏过程中这种聚合/解聚作用持续发生[16]。天然(RM)牛乳中,αs-CN、β-CN、κ-CN全部分布在胶束相,如图2(a)所示,α-La、β-Lg、BSA主要分布在乳清相中,如图2(b)所示。热处理后N-UHTD0和C-UHT-D0乳其胶束相中的κ-CN由4.61 g/L分别降低到2.54 g/L和2.31 g/L,αs-CN由17.48 g/L分别降低到16.93 g/L和17.31 g/L,β-CN由12.12 g/L分别降低到9.40 g/L和8.75 g/L,少量的α-La和β-Lg进入胶束相如图2(a)所示;N-UHT和C-UHT乳其乳清相中的α-La由2.48 g/L分别降低到2.07 g/L和2.10 g/L,β-Lg由5.56 g/L分别降低到4.57 g/L和4.45 g/L,较高质量浓度的αs-CN、β-CN、κ-CN进入乳清相,乳清相中蛋白浓度显著增加,如图2(b)所示。随着贮藏时间的增加,αs-CN、β-CN、κ-CN在胶束相的分布量降低,在乳清相的分布量增加,乳清蛋白在乳清相和胶束相的分布量变化不显著;贮藏6个月后,N-UHTM6和C-UHT-M6其胶束相中的κ-CN质量浓度分别降低到2.92 g/L和2.57 g/L,αs-CN质量浓度分别降低到10.77 g/L和11.94 g/L,β-CN质量浓度分别降低到4.59 g/L和5.39 g/L,如图2(a)所示;NUHT-M6和C-UHT-M6其乳清相中的κ-CN质量浓度分别增加到1.10 g/L和1.37 g/L,αs-CN质量浓度分别增加到3.76 g/L和3.65 g/L,β-CN质量浓度分别增加到1.28 g/L和1.54 g/L如图2(b)所示。α-La和β-Lg在乳清相分布量减少,胶束相分布量增加,但是变化幅度显著低于酪蛋白。综合分析,贮藏过程中NUHT乳的变化程度显著高于C-UHT乳。

图2 热处理和贮藏期乳蛋白组分在胶束相和乳清相中分布
酪蛋白胶束中心部分是磷酸钙,内部的酪蛋白被缠绕在一起形成一种微团簇的无定型状态,也就是无规则卷曲状态[17]。αs1-CN和αs2-CN是天然展开的蛋白质,具有扩展的卷曲构象,而β-CN和κ-CN具有熔融球蛋白状态的性质,具有结构致密、水化程度高、侧链柔性的特点,因缺乏固定的三维立体构象,对环境变化反应非常迅速[18]。当乳的温度升高至80℃时,酪蛋白组分均发生解离,其中热诱导的κ-CN从胶束相解聚量最大,αs-CN解聚最低[19]。κ-CN缺少磷酸丝氨酸残基,并且其亲水性C-末端的糖基化阻止了与其他蛋白质的强静电相互作用可能性[20]。κ-CN的N端区域疏水键连接到酪蛋白胶束并覆盖胶束表面,当酪蛋白解离后,覆盖在酪蛋白胶束表面的κ-CN会进一步与变性的乳清蛋白之间发生反应,同时αs-CN和β-CN从酪蛋白胶束上解聚下来的过程受到抑制[21]。κ-CN的解离随着温度的升高上呈线性增加,导致其在胶束相的解离逐渐增多[22]。酪蛋白的解离或者水解以及乳清蛋白与胶束表面酪蛋白结合,引起酪蛋白和乳清蛋白相分配行为的改变,这些变化与蛋白质交联、蛋白质分子共价和非共价相互作用、美拉德反应、蛋白质水解、去磷酸化等化学反应引起的蛋白质疏水性改变有关[16,23]。酪蛋白持续从胶束相解聚进入乳清相,增加了胶束聚集的可能性[24-25]。这些因素造成UHT乳的贮藏不稳定性。因此,UHT乳在贮藏期间均发生了酪蛋白从胶束相解聚进入乳清相,乳清蛋白与酪蛋白聚合从乳清相进入了胶束相,这种变化与贮藏期间蛋白质的和水解和贮藏时间呈正相关。
2.3 热处理和贮藏对UHT乳中游离氨基酸的影响
热处理能使N-UHT中游离氨基酸质量浓度由原乳的31.4 mg/L增加到35.1 mg/L,C-UHT由原乳的31.4 mg/L增加35.7 mg/L,两者差异不显著;贮藏过程中数据波动较大,但总量变化不显著;C-UHT中游离氨基酸质量浓度略低于N-UHT如图3(c)所示。
在贮藏过程中游离氨基酸质量浓度变化不显著(P<0.05),说明贮藏过程没有发生显著的蛋白质水解。贮藏过程中乳蛋白水解主要与乳中耐热性内原酶纤溶蛋白酶和细菌蛋白酶的活力有关,随着贮藏时间的增加,UHT乳中纤溶蛋白酶的活力逐渐增加[26],但是游离氨基酸质量浓度并没有显著增加。因为,纤溶蛋白酶和细菌蛋白酶并不能将乳蛋白直接水解成氨基酸,纤溶蛋白酶主要水解酪蛋白,释放出疏水性肽,导致乳蛋白发生凝聚作用;细菌蛋白酶主要水解κ-CN,类似于凝乳酶的作用,导致乳蛋白形成凝胶[10]。加热导致的游离氨基酸质量浓度增加与热诱导的乳蛋白氧化有关,如美拉德反应的Strecker降解过程形成游离态氨基酸[10]。
木其尔等人研究证明UHT处理使乳中游离氨基酸质量浓度降低了1.9%[27],与本研究得到的结论相反。可能与原乳的差异及分析方法有关,乳中游离氨基酸质量浓度较低,分析结果的相对误差也较大。在18种氨基酸中只有赖氨酸(Lys)随着贮藏时间的增加其质量浓度显著降低如图3(b)所示。贮藏过程中赖氨酸质量浓度的减少与美拉德反应有关。热诱导的美拉德反应使氨基酸等小分子营养物质的损失,随贮藏时间的延长,乳中蛋白质与乳糖会持续缓慢地发生美拉德反应[28]。美拉德反应是还原性羰基糖(主要是乳糖)与蛋白中的赖氨酸残基发生反应,尤其是酪蛋白中的赖氨酸残基,乳糖也与精氨酸,蛋氨酸,色氨酸和组氨酸反应,但以与赖氨酸的反应为主。乳糖与赖氨酸残基的ε-氨基反应形成席夫碱(乳糖基赖氨酸),热处理强度越大,乳糖和赖氨酸ε-氨基的结合越强烈,造成了赖氨酸的损失[29]。
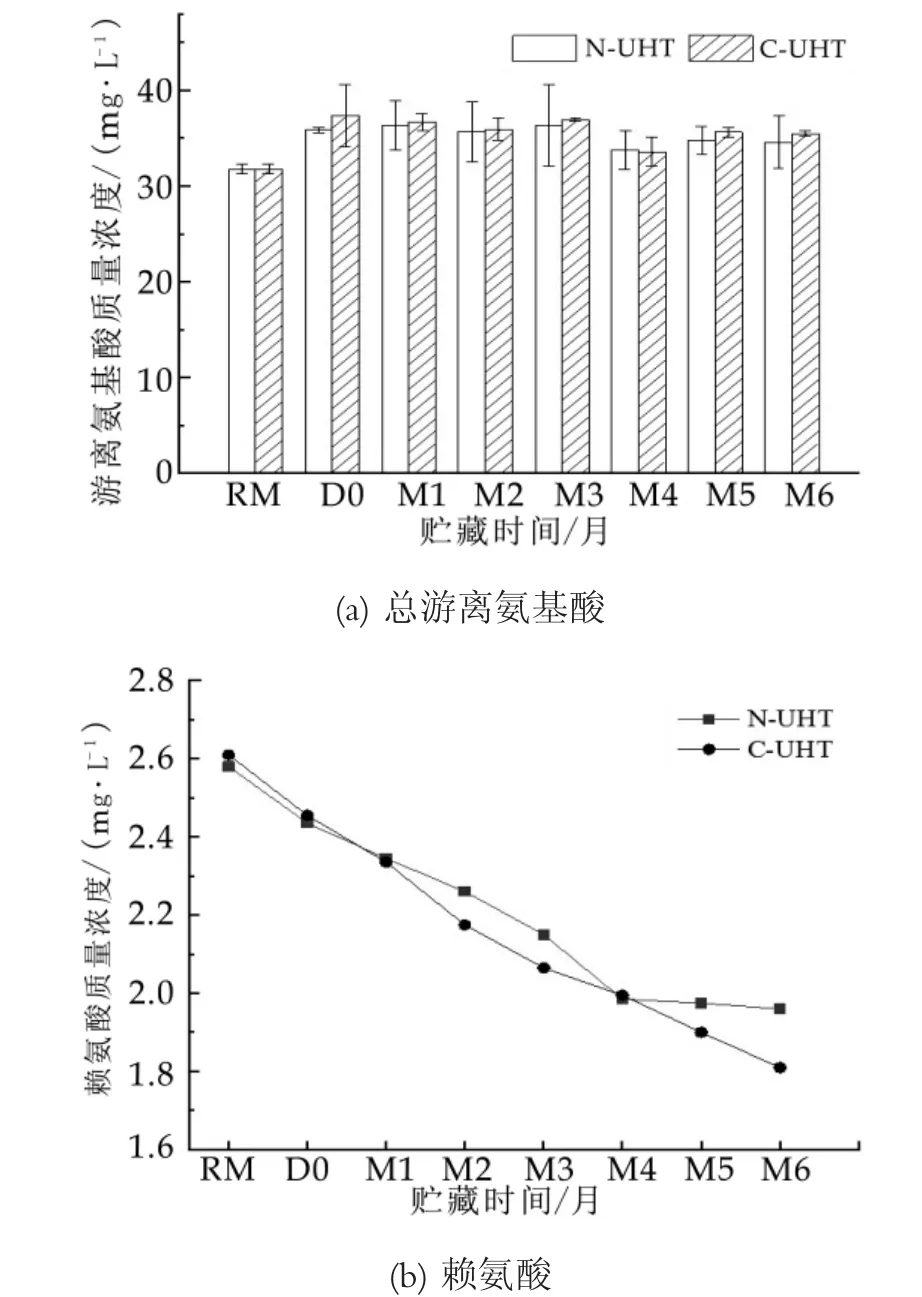
图3 UHT乳贮藏期间游离氨基酸质量浓度变化
2.4 热处理和贮藏过程中乳蛋白的降解
乳蛋白主要成分有αs1-CN、αs2-CN、β-CN、κ-CN、β-Lg、α-La、BSA,天然牛乳中这几种蛋白的比例和质量浓度基本上是固定的。热处理和贮藏过程乳蛋白发生了氧化、水解、降解等作用,乳蛋白组成的质量浓度发生了改变,影响了乳胶体的稳定性和牛乳的营养性。热处理导致所有的乳蛋白成分发生了降解,热处理后N-UHT和C-UHT中κ-CN质量浓度由原乳的4.61 g/L分别减少到4.45 g/L和4.27 g/L,αs1-CN质量浓度由17.48 g/L分别减少到16.93 g/L和17.31 g/L,β-CN质量浓度由12.12 g/L分别减少到10.98 g/L和10.51 g/L,α-La质量浓度由1.68 g/L分别减少到1.61 g/L和1.61 g/L,β-Lg质量浓度由3.92 g/L分别减少到3.74 g/L和3.64 g/L,如图4(a)所示。随着贮藏时间的增加,C-UHT和N-UHT中乳蛋白质量浓度逐渐降低,在贮藏2个月内,C-UHT和N-UHT之间变化不显著,贮藏时间超过2个月后,N-UHT中各种乳蛋白组分的降解程度显著高于C-UHT(P<0.05)。贮藏6个月后,N-UHT中总蛋白质量浓度由N-UHT-D0的37.71 g/L减少到N-UHT-6m的29.57 g/L;C-UHT中总蛋白质量浓度由C-UHTD0的37.34 g/L减少到C-UHT-6m的31.42 g/L,如图4(b)所示。

图4 热处理和贮藏过程中乳蛋白质量浓度变化
热诱导非酶脱酰胺反应、美拉德反应是引起乳蛋白降解的主要原因[9],贮藏过程中持续发生的蛋白氧化和蛋白水解加剧了乳蛋白的降解[10]。细菌蛋白酶优先水解酪蛋白,顺序为:κ-CN>β-CN>αs-CN[10];纤溶蛋白酶作为主要的内源性酶,其不水解乳清蛋白,其首选的底物是β-CN,对αs1-CN和αs2-CN也能水解,但对κ-CN有抑制作用[10]。蛋白水解释放的疏水性肽段参与了老化凝胶和苦味形成,造成UHT乳贮藏过程物理稳定性下降[30]。微生物源的蛋白酶和源于体细胞自溶所释放的酶会加剧UHT乳贮藏期内蛋白水解,体细胞中含有纤溶蛋白酶原激活剂,可增强非活性纤溶蛋白酶原向纤溶蛋白酶的转化,提高纤溶蛋白酶活力[31]。CUHT乳是原料乳采用离心除菌技术,有效地除去了原料乳中微生物与体细胞,所以在贮藏期C-UHT中蛋白水解程度低于N-UHT,体现在C-UHT中蛋白降解程度低于NUHT,能够在贮藏期内较好地保持乳蛋白营养价值。
3 结 论
热处理贮藏过程中酪蛋白质量浓度变化不显著,乳清蛋白质量浓度逐渐降低,NPN质量分数逐渐增加;贮藏6个月后,N-UHT和C-UHT的NPN从4.2%分别增加到8.7%和7.9%。热处理后酪蛋白解聚进入了乳清相,贮藏过程中酪蛋白继续从胶束上解聚;贮藏6个月后,N-UHT和C-UHT中酪蛋白在胶束相的质量分数由100%分别降低到53.4%和58.2%;N-UHT和C-UHT中乳清蛋白在乳清相的质量分数由95.5%分别降低到56.6%和54.8%,乳清蛋白的降解率远远低于酪蛋白。热处理导致所有的乳蛋白组分发生了降解,贮藏过程中乳蛋白继续降解,贮藏6个月后,N-UHT和C-UHT中总蛋白质量浓度由39.81 g/L分别降解到29.57 g/L和31.42 g/L;热处理使游离氨基酸总量由原乳的31.4 mg/L增加到35.1~35.7 mg/L,贮藏过程中游离氨基酸总量变化不显著。C-UHT乳的乳蛋白降解和乳蛋白相转移程度显著低于NUHT乳,因此,离心预除菌工艺通过改善原料乳的质量可以显著地改善UHT乳贮藏期内乳蛋白的质量,对保证贮藏期内UHT乳的品质和质量具有重要的作用。
