AJUBA在食管鳞状细胞癌KYSE150细胞中的功能
2022-02-28路子扬苏丽萍苏天园肖锦玲管亚玲蒲红伟
路子扬,季 敏,苏丽萍,刘 丽,苏天园,肖锦玲,管亚玲,蒲红伟
(新疆医科大学1基础医学院,2第一附属医院病理科,3远程医疗中心,4学科建设科,乌鲁木齐 830054)
食管癌(esophageal cancer,EC)是食管上皮细胞的恶性肿瘤,以食管鳞状细胞癌(esophageal squa⁃mous cell carcinoma ,ESCC)为主,是发病率较高的十大肿瘤之一,其死亡率占世界肿瘤死亡率的第六位[1]。近年来食管癌的发病率虽呈现阶段性下降,但患食管癌总人数已超过45 万,死于食管癌的人数高达30万[2]。食管癌早期症状并不明显,患者常在肿瘤增大,吞咽有异物感时进行检查,此时肿瘤已发展至中晚期,导致治疗效果差,术后五年生存率低[3],因此早期诊断及治疗意义重大。基因序列监测技术的发展使肿瘤治疗与精准治疗相结合,提高了肿瘤的早期诊断与预后。已有多项研究表明,AJUBA 参与食管癌、皮肤癌、结直肠癌的发展过程,但有关AJUBA在食管鳞癌中的具体作用机制的研究较少。本研究通过分析AJUBA 在食管鳞癌发展过程的作用机制,为其早期进展与预后提供理论依据,对寻找新的治疗靶点具有重要意义。
1 材料与方法
1.1 试剂与仪器食管鳞状细胞癌(ESCC)细胞株KYSE150 购自上海中国科学院细胞库。慢病毒由上海吉凯公司构建,AJUBA 抗体及GAPDH 内参抗体均购自英国ABCAM 公司。细胞计数(CCK-8)检测试剂盒、ECL荧光显色试剂盒购自北京博奥森公司。高糖培养基、0.25%胰酶、双抗和胎牛血清均购自美国Gibco 公司,BCA 蛋白定量试剂盒购于中国索莱宝公司。 TRIzol 试剂、primeScriptTMone stEP RT-PCR kit购于美国Invitrogen 公司,QuantiNova SYBR Green PCR Kit 购于德国QIAGEN 公司,8.0 μm 孔径的Tran⁃swell小室购于美国Corning公司。
1.2 方法
1.2.1 细胞培养 人正常食管鳞状上皮(SHEE)细胞系、KYSE150 细胞系,采用含10%胎牛血清与1%的双抗的高糖培养基(49:1),置于37℃,5%CO2细胞培养箱中培养。实验均在细胞对数生长期进行。
1.2.2 细胞转染 将对数生长期的KYSE150 细胞用胰酶消化,按每孔5×105个细胞接种至6 孔板,加入培养液1 mL,置于培养箱培养24 h。第2 天,计算慢病毒所需用量,-80℃冰箱取出,冰浴融化备用。显微镜下观察细胞生长状态,细胞生长至80%开始转染。先换培养液,再将慢病毒液加入6孔板中,轻柔混匀,置于培养箱过夜,孵育12 h,取出6 孔板,弃除上清液,加入10%胎牛血清(FBS)培养液,继续培养72~96 h,观察荧光表达情况,感染率高于80%,可用于后续实验。细胞稳定传代2~4 代后,进行RNA 及蛋白质含量的检测。将细胞分为对照组与敲降组。其中对照组包括KYSE150 组(未转染的KYSE150 细胞)和KYSE150shnon 组(转染随机序列),敲降组为KYSE150shAJUBA 组(分别转染KYSE150shAJUBA-1、KYSE150shAJUBA-2、KYSE150shAJUBA-3)。
1.2.3 CCK-8 细胞增殖实验 细胞生长至对数期时通过胰酶消化细胞制成细胞悬液,并进行细胞计数。调整细胞密度后接种至96 孔板内,每孔1 000 个细胞,置于细胞培养箱中。接种细胞4h 后,显微镜下观察细胞已贴壁,弃去旧培养基,每孔加入完全培养基100 μL,CCK-8 10 μL,每孔总体积共110 μL。根据预实验结果,培养箱中孵育2 h。将96 孔板置于酶标仪中震荡30 s,吸光度450 nm,记录吸光度(OD)值,标计为0 h。此后24、48、72、96 及120 h 的操作同上,每孔设置3个复孔。实验共重复三次。
1.2.4 Transwell 细胞迁移实验 采用细胞周期抑制剂Mitomycin B 进行预处理,以饥饿细胞24 h。每孔细胞数为1×105个。在Transwell 小室下室中的每个小室预先加入500 μL 含10 %胎牛血清的完全培养基,上室加入细胞悬液,将24孔板置于细胞培养箱培养,4%组织细胞固定液4 ℃固定30 min,0.1%结晶紫溶液700 μL,室温下染色5 min。下室倒置于显微镜下,每个小室随机选取5 个视野,拍照,通过Image J软件进行计数。实验共重复三次。
1.2.5 实时荧光定量聚合酶链反应(qRT-PCR) 采用Trizol 法提取细胞总RNA。取1 μg 总RNA 参照说明书进行反转录反应,反应体系37℃,30 min 反应,85℃5 s 终止反应。qRT-PCR 反应依照试剂盒说明进行。引物序列如下:AJUBA-正向引物:5'-CCAAG⁃TATACTGTGTCACCGACT-3',AJUBA-反向引物:5'-CAAAGTGATAATCCCGGTCCA-3',GAPDH-正向引物:5'-GAAGGTGAAGGTCGGAGTC-3',GAPDH-反向引物:5'-GAAGATGGTGATGGGATTTC-3'。实验共重复三次。
1.2.6 Western blotting 实验 将KYSE150 细胞株与人正常食管鳞状上皮(SHEE)细胞株用胰酶消化,PBS 清洗3 次,加入细胞裂解液,冰上裂解30 min,12 000 r/min,4℃离心30 min,收集上清液。采用二喹啉甲基(BCA)蛋白定量法进行十二烷基磺酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE)。转膜完成后,5 %脱脂牛奶封闭2 h,加入一抗,4 ℃孵育过夜。Tris-HCl缓冲盐溶液加离子去污剂(TBST)洗涤3次后,加入二抗,室温孵1 h,通过底物化学发光法(ECL)曝光检测蛋白质的表达情况。实验共重复三次。
1.3 统计学处理采用SPSS 21.0统计学软件进行分析,计量资料以均数±标准差(±s)表示,组间比较采用t检验,多组间比较采用单因素方差分析与非参数检验。检验水准α=0.05。
2 结果
2.1 慢病毒转染敲降AJUBA 在KYSE150 中的表达情况在KYSE150 细胞株中慢病毒转染shAJUBA 96 h 后,荧光显微镜下的KYSE150shnon 与KYSE150shAJUBA 细胞呈绿色荧光,感染率均高于80%。随后进行流式筛选出感染细胞进行培养,见图1A。Western blotting 结果显示,与对照组相比,敲降组AJUBA 蛋白表达水平下降,且KYSE150shAJUBA-3下降最为显著(P<0.000 1),见图1B、C;qRT-PCR 结果显示,敲降组AJUBA 的mRNA 水平下调,KYSE150shAJUBA-3 的mRNA 表达水平下降最显著(P<0.000 1),见 图1D。 挑 选 敲 降 率 最 高 的KYSE150shAJUBA-3用于后续实验。
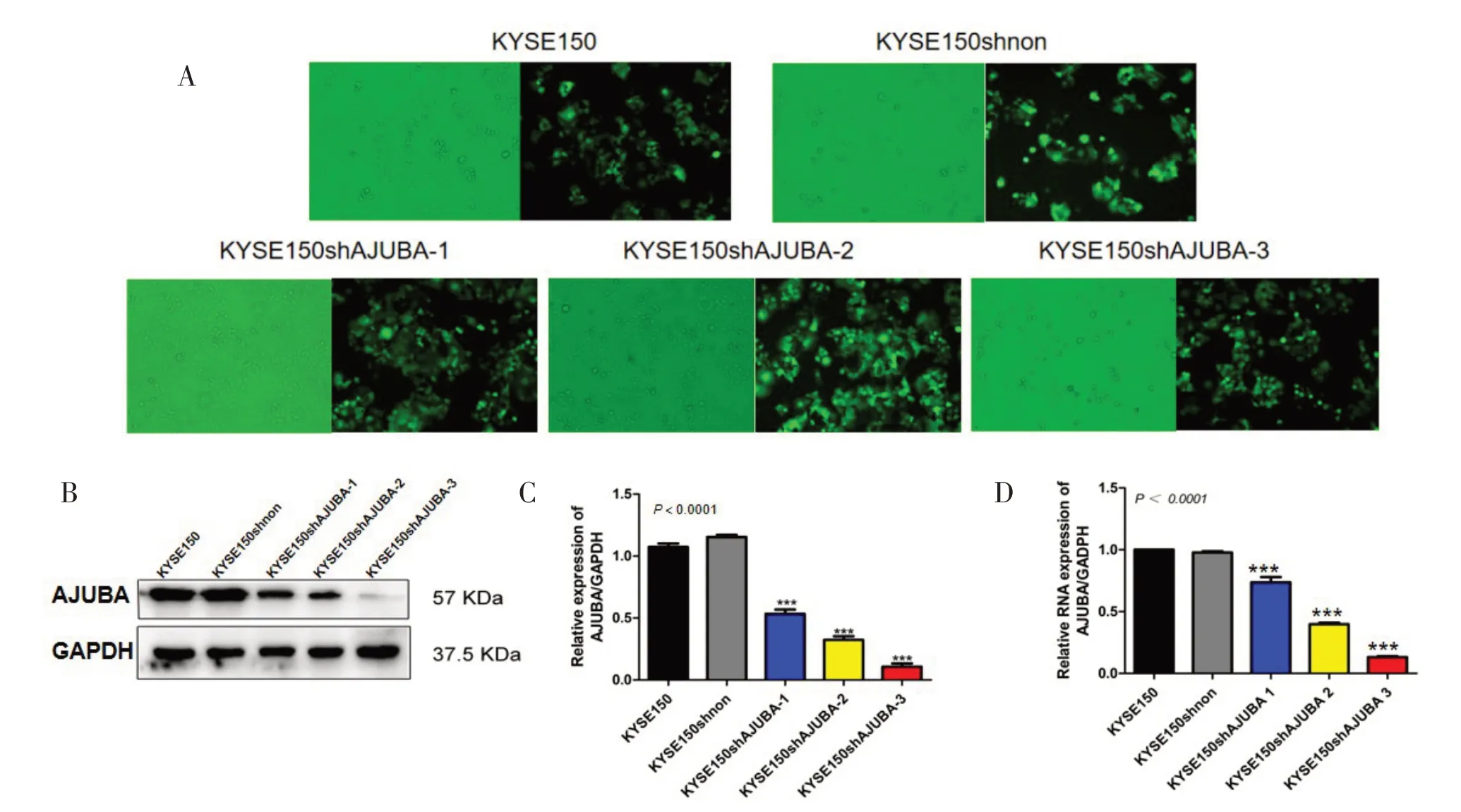
图1 KYSE150敲降AJUBA检测AJUBA敲降效率
2.2 敲降AJUBA表达后抑制KYSE150细胞的增殖能力结果CCK-8增殖实验检测结果显示,KYSE150与KYSE150shnon相比,两组的细胞增殖速度无差异(P>0.05),而KYSE150shAJUBA分别与KYSE150、KYSE150shnon相比较,KYSE150shAJUBA在各个时间点的增殖速度均降低,差异有统计学意义(P<0.001),见图2。

图2 AJUBA敲降后对细胞增殖能力的影响(***P<0.001)
2.3 敲降AJUBA表达后抑制KYSE150细胞的迁移能力对照组中KYSE150与KYSE150shnon Tran⁃swell小室下室的细胞数目无明显差异(P>0.05);敲降组KYSE150shAJUBA组与对照组相比,敲降组细胞迁移数目减少,迁移能力显著下降,差异具有统计学意义(P<0.000 1),见图3。
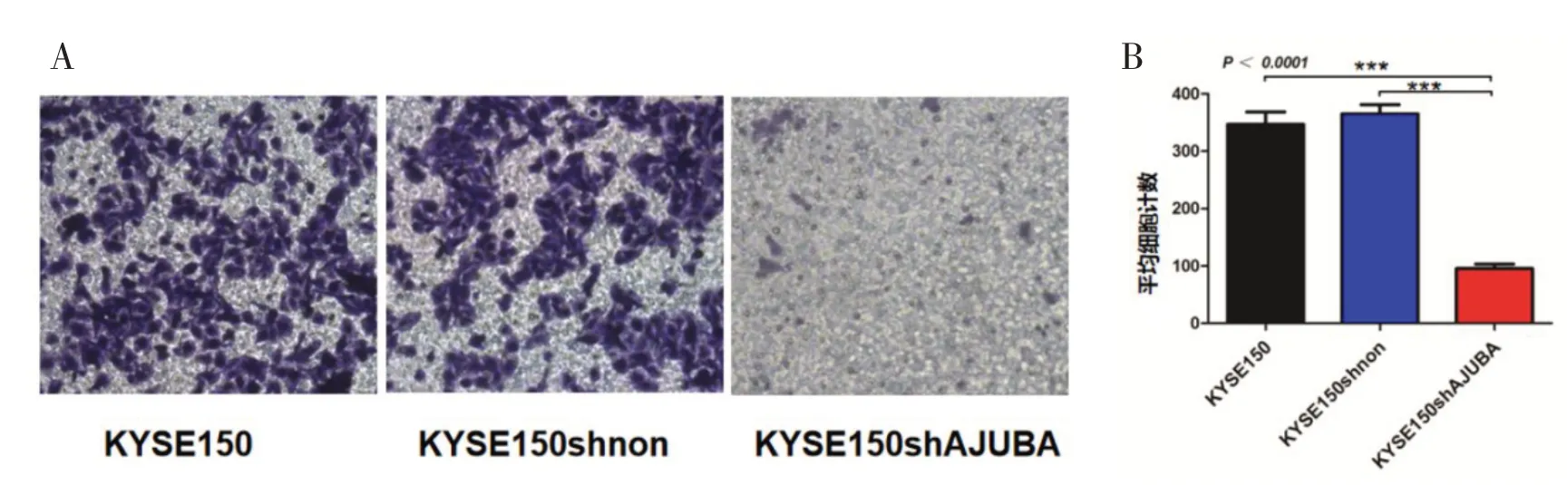
图3 AJUBA敲降对ESCC细胞迁移能力的影响
3 讨论
食管鳞癌的发展是由环境因素、遗传、生活习惯与病毒等多因素导致的。寻找影响食管鳞癌发展的因素对及时筛查食管鳞癌及食管鳞癌的后续治疗具有重要意义。
AJUBA 是一种多LIM 结构域蛋白,属于AJUBA/ZyCin 家族[4],其结构及亚细胞定位在生长因子、细胞基质黏附与机械信号等外部信号的转导中起着接头蛋白和支架蛋白的作用[5]。AJUBA 蛋白主要定位于细胞核与细胞质,作为穿梭于不同亚细胞之间的蛋白质,AJUBA 蛋白在细胞表面及细胞核之间传递信号中发挥着重要作用[6]。已有研究表明,AJUBA 在食管鳞癌细胞中高表达发挥促癌作用并与食管鳞癌预后相关[7-8],但因其研究结果较少仍待进一步研究探讨。本研究前期实验结果表明,AJUBA 在食管鳞癌组织细胞中高表达,并高于细胞核与细胞质,而在正常食管鳞状组织细胞中,AJUBA 在细胞核低表达[8]。AJUBA 在正常食管鳞状组织细胞核中表达,而ESCC组织细胞的细胞核与细胞质中表达,这可能与AJU⁃BA蛋白质在细胞核中主要是作为转录抑制因子来抑制基因的转录;在细胞质中,AJUBA 蛋白质在细胞质中发挥有丝分裂作用;在细胞表面,AJUBA 蛋白可以与钙黏蛋白(E-cadherin)结合从而进行细胞连接[5-6,9]。因此,AJUBA 可以执行多种细胞功能,包括细胞黏附、运动、有丝分裂、存活、基因表达、microR⁃NA 处理及机械力传感[10-11]。研究表明,AJUBA 通过调节Wnt、RAS/ERK、JAK/STAT 和Hippo 等主要信号通路[12],并作为Snail、Sp1 和核激素受体等关键转录因子的共同调节因子参与肿瘤进展[13-15]。
为探讨AJUBA 在食管鳞癌细胞中高表达所发挥的生物学功能,本研究通过慢病毒转染稳定下调KYSE150 中AJUBA 蛋白的表达,采用CCK-8 增殖实验检测敲降AJUBA 蛋白在食管鳞癌细胞中的表达对细胞增殖的影响,结果显示敲降AJUBA 后食管鳞癌细胞的细胞增殖速度受到了显著抑制。这与AJUBA在多种恶性肿瘤中促进细胞增殖的研究结果相符[11-13],可能是AJUBA 的过表达或AJUBA 的LIM 前(Pre-LIM)结构域的过表达激活AJUBA,使原本在正常食管鳞状上皮细胞的细胞核中发挥转录抑制功能的AJUBA 转移至细胞质内发挥中心体功能促进有丝分裂,导致细胞增殖。敲降ESCC 细胞中AJUBA 的表达后Transwell 小室下室的细胞数目相较于对照组显著减少,敲降AJUBA 的表达将会降低细胞的侵袭与迁移能力,这表明AJUBA 可能促进食管鳞癌中上皮细胞-间充质转化(Epithelial-mesenchymal transition,EMT)的发生,AJUBA 增多将促进ESCC 细胞的侵袭与迁移。这与AJUBA 在其他肿瘤中过表达促进肿瘤细胞增殖与迁移的结果相符[8,12,16]。AJUBA 与细胞质中不同的多聚体蛋白复合物有关,它可通过其LIM基序与α-catenin 相互作用或通过其Pre-LIM 区域与F-肌动蛋白(F-actin)相互作用控制细胞黏附的蛋白质复合物结合[17]。因此,AJUBA 通过将E-cadherin/αcatenin 黏附受体复合物与细胞骨架连接,有助于黏附连接的形成与稳定[16]。EMT 是肿瘤转移的可逆的初始过程,在此过程中,极化的上皮细胞通过细胞骨架重排转变为分离的间充质细胞,导致黏附减少和细胞运动增加,增强肿瘤的传播[18]。Zhang 等[19]的研究表明,AJUBA 参与并促进肝细胞癌的EMT 过程,在肝细胞癌中AJUBA 的高表达将提高细胞侵袭性及发生不良预后的概率。
综上所述,AJUBA 促进食管鳞状细胞癌的增殖与侵袭,可作为ESCC 患者预测肿瘤大小与浸润深度的潜在分子标志物,有望成为ESCC治疗的新靶点。
