外泌体在肝脏损伤中的研究进展
2022-02-24王子明
王子明,王 华,2,3
(1.安徽医科大学第一附属医院肿瘤科,2.安徽医科大学药学院,肝病研究所,3.炎症免疫性疾病安徽省实验室,安徽 合肥 230032)
在多细胞生物中,体内稳态的维持需要不同功能的细胞相互作用与协调,而机体在面对不同的应激和损伤时也需要细胞间的通讯来应答。研究发现[1],细胞不仅能够通过直接接触和分泌各种激素、细胞因子等化学信号传递信息,而且还能通过膜衍生的纳米级的细胞外囊泡进行交流,外泌体作为细胞外囊泡的一种类型,携带着大量的蛋白质、核酸、脂质和代谢产物等,在细胞间和器官间的信息传递中有着不可或缺的作用。
各种类型肝损伤的发病机制目前尚不完全清楚,外泌体作为细胞间通讯的新介质,能够在肝脏受到损伤时持续分泌,其携带的生物信息可以传递给邻近或者远距离细胞,从而引发受体细胞相关功能的改变,在多种类型肝损伤的发生发展中扮演着不同的角色。本文就外泌体在不同类型肝损伤中发挥的潜在作用予以综述。
1 外泌体的产生、释放与作用方式
外泌体是由细胞分泌的直径约为40~160 nm的磷脂双分子层结构,其产生与释放包括内体分选转运复合体(endosomal sorting complex required for transport,ESCRT)依赖和非依赖两种方式,ESCRT由4种可溶性多蛋白复合物组成,包括ESCRT-0、ESCRT-Ⅰ、ESCRT-Ⅱ和ESCRT-Ⅲ,参与了蛋白质的分选以及外泌体的装载等,是进化上高度保守的过程[1-2]。
外泌体依赖于ESCRT的发生机制:首先在胞吞和细胞膜内陷的作用下,早期核内体形成,之后不断成熟形成晚期核内体,晚期核内体将其限制膜的一部分向腔内萌发形成腔内囊泡(intraluminal vesicles,ILVs),随后ESCRT被招募到内体膜的胞质一侧,将所选蛋白分选入ILVs,ILVs不断累积并逐渐成熟,从而形成了多泡体(multivesicular bodies,MVBs)。随后MVBs既可以与溶酶体融合被降解,也能够与质膜融合,完成外泌体的胞外分泌[3-4]。脂质驱动可能在ESCRT非依赖机制中发挥作用,这其中研究较多的是神经酰胺,参与了ILVs的生成[1]。
外泌体通过多种方式与靶细胞相互作用[3,5]:(1)配受体相互识别激活细胞内信号。(2)靶细胞通过细胞膜融合吸收外泌体。(3)靶细胞通过内吞机制(吞噬作用、大胞饮作用或受体介导的内吞作用)内化外泌体。
2 外泌体与肝损伤
生理条件下的外泌体能够持续分泌,而在应激或病理状态下,外泌体的内容物可能发生定性和定量的变化[6]。肝损伤可由脂质、酒精、胆汁、药物以及病毒等多种原因引起,不同类型肝损伤释放的外泌体装载不同的内容物,作用于受体细胞后产生不同的影响,从而推动或者延缓肝脏疾病的发生发展。近年来,微小RNA(microRNA,miRNA),一种高度保守的非编码小RNA,在各种疾病中的作用已被广泛研究,miRNA是生物体内基因表达的关键调控因子,外泌体携带包括miRNA在内的丰富的核酸,可以通过翻译阻滞或mRNA降解来抑制靶向mRNA的蛋白表达过程[7]。下文将重点介绍外泌体中的miRNA在各种类型肝损伤中的作用。
2.1 外泌体与非酒精性肝病非酒精性肝病包括单纯的肝脏脂肪变性、非酒精性脂肪性肝炎(nonalcoholic steatohepatitis,NASH)以及相关的肝硬化和肝细胞癌一系列疾病。NASH是非酒精性脂肪肝发展为肝硬化并最终进展为肝细胞癌的关键事件,炎症的出现是NASH的特征之一。脂毒性的肝细胞能够通过各种分子途径诱发肝细胞应激,引发肝细胞损伤,从而激活炎性细胞并释放炎症因子,外泌体在肝细胞和炎症细胞的这种交叉对话中起到媒介作用。研究发现,脂毒性的肝细胞能够释放携带有miR-192-5p的外泌体,作用于巨噬细胞后,通过调节Rictor/Akt/FoxO1信号通路促进巨噬细胞向M1型极化,并刺激促炎因子的释放,从而推动了炎症的进展[8]。另外,脂质还能够抑制肝细胞中MVBs在溶酶体的降解,导致含有miR-122-5p的外泌体分泌增加,并激活巨噬细胞向M1型极化,促进了炎症的发生与发展[9]。Hirsova等[6]研究发现,脂毒性肝细胞能够释放含有肿瘤坏死因子相关凋亡诱导配体的外泌体,作用于位于巨噬细胞上的死亡受体5(death receptor 5,DR5),并以DR5依赖的方式激活NF-κB信号通路,从而促进白细胞介素-1β(interleukin-1β,IL-1β)与白细胞介素-6(interleukin-6,IL-6)的释放。这些研究都表明,外泌体作为载体,携带着脂毒性肝细胞发出的信号分子,诱导巨噬细胞的激活以及炎症因子的释放,推动了非酒精性肝病的炎症进展,这部分解释了NASH发生的潜在机制。
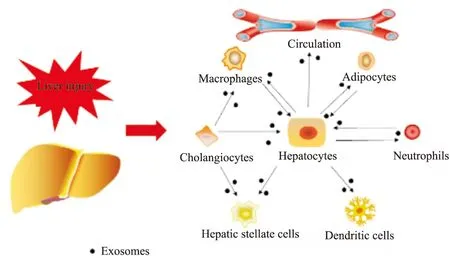
Fig 1 Roles of exosomes in liver injury
内质网参与调节脂代谢稳态,脂毒性物质(如饱和游离脂肪酸等)能够诱发内质网应激,可能会导致NASH的发生与发展[10]。研究发现[11],内质网应激诱导的脂肪细胞分泌的外泌体能够通过醛酮还原酶家族1成员B7触发NASH。另外,脂质诱导肝细胞发生内质网应激后,肌醇依赖性激酶1α通过X盒结合蛋白1促进神经酰胺的新生合成增加,介导肝细胞以ESCRT非依赖的方式释放外泌体后招募单核细胞到达肝脏,推动了NASH的进程[10]。这也说明了内质网应激作为脂毒性亚细胞应激的突出类型,其能够从外泌体的生物学过程解释NASH的病理过程,有利于对疾病发生机制的全面理解。
另外,有趣的是,髓样特异性IL-6能够促进巨噬细胞和中性粒细胞释放含有miR-223的外泌体,作用于肝细胞后,减少肝细胞中具有PDZ结合域的转录活化因子(transcriptional activator with PDZ-binding motif,TAZ)的表达,TAZ具有促纤维化的作用,抑制TAZ能够延缓非酒精性肝病向NASH乃至肝纤维化的进展,从而为非酒精性肝病的治疗提供了新思路[12]。NASH作为非酒精性肝病进展的关键事件,早期给予适当的干预,能够明显减少不良的预后,外泌体在这一事件的进展中发挥重要作用,将其作为治疗靶点将具有良好的临床前景。
此外,外泌体在非酒精性肝病中的作用不仅仅局限于推动疾病的进展,还充当了机体不同器官之间交流的媒介。研究发现[7],脂肪变的肝细胞释放的外泌体携带microRNA-1,能够抑制血管稳态的调控因子,从而引起内皮炎症的发生和动脉粥样硬化的形成,推动了心血管疾病的发生发展。此研究揭示了肝源性外泌体在肝脏和血管系统之间的远距离通讯中的重要作用。另外,Zhao等[13]研究发现,脂质超载后,肝细胞释放富含let-7e-5p等miRNA的外泌体,作用于脂肪细胞后,导致脂肪重构,从而引起脂质在肝脏、脂肪等器官之间重新分布,这也说明了肝脏不仅仅是一个被动的异位沉积器官,而且能够感知不同代谢状态,并通过外泌体协调功能,以重塑脂肪组织,来适应脂肪超载引起的代谢变化。
2.2 外泌体与酒精性肝损伤最近,大量的研究证明了酒精性肝损伤中肝细胞释放的外泌体介导了酒精性肝病(alcoholic liver disease,ALD)的炎症进程。如蛋白质组学分析已表明酒精性肝损伤循环中的外泌体增加,并携带酒精相关特异性蛋白质,且循环外泌体中的热休克蛋白90介导了巨噬细胞在肝脏的激活[14]。另外,酒精性肝细胞释放富含线粒体双链RNA的外泌体,通过刺激库普弗细胞上的Toll样受体3,促进了IL-1β的产生与分泌,激活的库普弗细胞进一步招募γδT细胞并促进了白细胞介素-17A(interleukin-17A,IL-17A)的分泌,加速了炎症的进展[15]。
另外,有研究对ALD中外泌体分泌的增多做出了部分解释,通过慢性酒精消耗可以提高肝脏中miR-155的水平,从而抑制自噬过程中的mTOR通路,下调溶酶体相关膜蛋白的表达,破坏溶酶体的功能,自噬体与多泡体结合,并将其内容物释放到细胞外,外泌体的分泌增加[16],此研究也进一步表明,在ALD中,酒精可以阻断自噬体和溶酶体融合并促进外泌体的分泌,自噬和外泌体的生物发生是两个复杂的过程,而由于其部分共同的分子机制,使得miR-155能够在自噬和外泌体分泌之间产生串扰作用,因此,自噬和外泌体分泌之间的相互作用与影响需要进一步研究。
目前,有关外泌体在ALD中的研究大多集中在炎症,然而ALD也是一种代谢性疾病,外泌体在ALD的代谢过程中是否发挥作用,尚需进一步探索。另外,外泌体在体液中广泛存在,且携带酒精相关特异性组分,也可作为诊断酒精性肝损伤的标志物,但其特异性与灵敏度尚需评价。
2.3 外泌体与胆汁淤积性肝损伤胆汁流动障碍和胆汁酸代谢紊乱是胆汁淤积性肝损伤的主要诱因,肝内胆汁酸的淤积激活炎症反应,导致肝内胆管破坏以及肝细胞损伤,最终导致胆管炎、肝纤维化和潜在的肝硬化。胆管细胞分泌的外泌体在胆汁淤积性肝损伤的发生以及疾病的进展中扮演着重要角色。研究表明[17],在胆汁淤积的情况下,胆管细胞上调了长链非编码RNA H19(long non-coding RNA H19,lncRNA H19)的表达,由外泌体装载并作用于肝细胞,在肝细胞中通过抑制启动子活性和降低mRNA稳定性来抑制小异源二聚体伴侣受体的表达,从而导致了胆汁淤积性肝损伤的发生与发展。此外,外泌体介导的巨噬细胞的活化在胆汁淤积性肝损伤的炎症进展中也起着关键性的作用,同样在胆汁淤积的情况下,携带有lncRNA H19的外泌体被库普弗细胞摄取后,诱导了炎症因子的表达、单核细胞的趋化以及M1型巨噬细胞的极化,从而导致了肝脏的炎症进展[18]。另外,肝星状细胞的激活是促进慢性胆汁淤积性肝病进展的主要驱动力。有研究进一步表明,胆管细胞释放的含lncRNA H19的外泌体能够作用于肝星状细胞,从而促进肝星状细胞的活化和增殖,加速了胆汁淤积性肝纤维化的进程[19]。
2.4 外泌体与药物性肝损伤目前普遍认为,药物引起的肝细胞损伤可能是由免疫反应介导的,并且大多数延迟时间较长的特异型药物性肝损伤是由适应性免疫攻击引起的[20]。外泌体可能在肝脏对药物的免疫应答中发挥作用。Ogese等[21]通过对药物性肝损伤中肝源性外泌体内包裹的蛋白质进行表征,发现经药物修饰后的蛋白可以通过外泌体从组织细胞转移到免疫系统,最终以肽片段的形式呈递至树突状细胞表面的人白细胞抗原分子,触发抗原特异性T细胞反应,介导肝细胞的损伤。另外,在药物性肝损伤当中,肝细胞来源的外泌体能够被单核细胞吸收并传递miRNA,改变单核细胞转录组,对单核细胞进行免疫抑制,从而促进肝脏的免疫耐受,T细胞反应可能正是由于免疫耐受的丧失导致的[22]。另外,Yarana等[23]认为,化疗药物阿霉素处理后的肝源性外泌体中的4-羟基壬烯醛——阿霉素诱导的氧化应激过程中产生的毒性脂质过氧化物,能够上调巨噬细胞抗氧化酶硫氧还蛋白1水平,并通过调节抗炎与促炎之间的平衡从而保护肝脏免受炎症损伤,这也表明了并不是所有暴露于氧化应激条件下释放的外泌体都是有害的,外泌体也是一种新型的氧化还原免疫应答介质,提高了我们对癌症治疗过程中先天免疫应答的认识。
2.5 外泌体与病毒性肝损伤外泌体在病毒性肝损伤中可能扮演着不同的角色。首先,被乙型肝炎病毒(hepatitis B virus,HBV)感染后的肝细胞分泌的外泌体被证实具有免疫抑制功能[24]。另外,外泌体可以将干扰素诱导跨膜蛋白2转运至树突状细胞,抑制干扰素α(interferon-α,IFN-α)的激活,并阻断了外源性IFN-α的抗HBV治疗效果[25]。其次,在丙型肝炎病毒(hepatitis C virus,HCV)感染导致的病毒性肝损伤当中,被HCV感染的肝细胞分泌的外泌体中的miR-192通过上调肝星状细胞中转化生长因子β1的表达,刺激肝星状细胞向肌成纤维细胞活化,推动了肝纤维化的进程[26]。
有趣的是,在病毒性肝损伤当中,外泌体不仅在加速病毒复制以及推动纤维化进程中发挥作用,还可以通过干扰素抑制病毒复制从而介导抗病毒反应。巨噬细胞来源的含有hsa-miR-574-5p的外泌体能够抑制肝细胞中HBV的复制和转录[27]。Zhao等[28]的研究发现,HBV感染的肝细胞衍生的外泌体含有HBV编码的miRNA,即HBV-miR-3,下调了巨噬细胞中细胞因子信号转导抑制因子的表达,从而激活JAK/STAT信号通路,增强了干扰素介导的抗病毒反应,并且巨噬细胞向M1表型极化,IL-6分泌增加,减轻了病毒性肝损伤。
外泌体对病毒性肝损伤的影响是多方面的,其可以清除病毒复制的不利因素,加速纤维化进程,也能够通过细胞间通讯的作用激活先天免疫应答来抑制病毒复制,因此,外泌体在病毒性肝损伤当中的作用是复杂的,尚需进一步的研究与探讨。
3 结论
外泌体中含有蛋白质、核酸、脂质等多种生物大分子,其介导的细胞通讯作用在机体中可能扮演着不同的角色,不同类型的细胞可以在生理条件下通过外泌体维持机体稳态,也能够通过激活受体细胞的不同通路改变其功能,进而保护肝脏免受损伤或导致各种类型肝脏疾病的发生发展,因此,外泌体在不同条件下功能的复杂性需要进一步研究。另外,病理生理条件下的肝源性外泌体的作用不仅仅局限于肝脏的内环境,还可以通过远距离的通讯影响其他组织器官,所以,外泌体对机体的影响是广泛而深远的。同时,在循环和肝脏中存在的特异性外泌体又使得其在肝损伤的诊断、治疗和预后等方面具有较好的临床前景,可作为肝损伤新型的治疗载体、免疫调节剂等。外泌体的研究与应用也不仅仅局限于肝脏疾病,同时为许多疾病的诊断和治疗提供了新的方法和策略。但是,外泌体的分离纯化等技术尚需简化和优化,虽然许多新的方法已经开发,但是尚未进行有力的评价,未来对于外泌体介导的疾病发生的具体机制也要进一步研究。
