CRISPR/CAS9靶向肝癌细胞Oct3/4增强化疗药物作用效果的研究
2022-02-23张树交查中明
张树交 查中明 武 阳
人类肝细胞癌(HCC)是最常见的恶性肿瘤之一,被公认为是全球所有与癌症相关的死亡率的第二大诱因[1]。 HCC的死亡率高达95%,5年生存率极短,仅为6.9%[2]。与HCC发生率密切相关的危险因素包括乙型肝炎病毒(HBV)和丙型肝炎病毒(HCV)感染,吸烟,饮酒,饮食中摄入黄曲霉毒素,糖尿病和肥胖[3]。因此,至关重要的是发现影响化疗药物对HCC发挥作用的关键分子机制,以改善HCC患者的诊断和治疗。Oct3/4是POU家族的转录因子,与八聚体序列基序结合以激活下游基因的表达[4]。Oct3/4的表达升高与多种癌症相关,包括膀胱癌,非小细胞肺癌和肝细胞癌[5]。尽管已经进行了许多研究,但是Oct3/4在癌症中的预后意义仍然存在争议,并且Oct3/4与化疗的关系尚未得到充分认识[6]。基于此,本研究旨在探讨CRISPR/CAS9靶向敲除Oct3/4基因后化疗药物索拉菲尼(Sorafenib)对肝癌细胞的作用影响。
1 材料与方法
1.1 材料
Li-7、HepG2、Huh7、BEL-7405、LX-2细胞均购自普诺赛公司(货号:CL-0139、CL-0103、CL-0120、CL-0321、CL-0560)。Sorafenib购自深圳市益百顺科技有限公司(货号:Sorafenib)。pX459、,pX460-1、pX461-1均购自Addgene。Cas9慢病毒购自吉满生物科技(上海)有限公司(货号:GM-KC-100001)。Oct3/4和GAPDH抗体均购自abcam(货号:ab124826、ab8245),Total RNA Isolation Kit购自南京诺唯赞生物科技股份有限公司(货号:RC101-01)。逆转录反应试剂盒购自北京拜尔迪生物技术有限公司(货号:DEM201-50T)。细胞凋亡检测试剂盒购自上海碧云天生物技术有限公司(货号:C1062L)。
1.2 方法
1.2.1 细胞培养和加药处理 Li-7、HepG2、Huh7、BEL-7405、LX-2均用RPMI-1640(补充10%胎牛血清和1%双抗)维持,并放于含5%CO2的37 ℃培养箱培养。索拉菲尼(Sorafenib)的处理终浓度为20 μM。
1.2.2 CRISPR/CAS9基因敲除与阳性细胞的筛选 Cas9慢病毒感染HepG2细胞,并使用2 μg/ml的嘌呤霉素进行筛选Cas9阳性细胞。使用CRISPR设计工具(http://crispr.mit.edu/)设计了靶向OCT3/4基因片段的单向导RNA(sgRNA)序列:5'- CCAAGCTCCTGAAGCAGAAG-3'。pX459(包含U6启动子-sgRNA插入位点-sgRNA的3个载体),pX460-1(包含U6启动子-sgRNA插入位点-sgRNA支架和CAG启动子-Cas9-T2A-嘌呤霉素N-乙酰基转移酶基因-牛生长激素多腺苷酸化信号)和pX461-1(包含U6启动子-sgRNA插入位点-sgRNA支架,以及CAG启动子-嘌呤霉素N-乙酰基转移酶(PuroR)-牛生长激素聚腺苷酸化信号)用于sgRNA的亚克隆。转染后2周以低密度培养细胞,允许单个菌落(50个菌落)扩增,然后通过PCR测序进行选择。
1.2.3 RNA抽取与实时荧光定量PCR 使用Total RNA Isolation Kit从细胞系中分离出总RNA。使用凝胶电泳和2000 Nanodrop分光光度计评估提取的RNA的质量和数量。使用随机六聚体引物和MMLV逆转录酶对总RNA(1 μg)进行反转录。为了定量Oct3/4和GAPDH的转录水平,定量RT-PCR反应包含Premix Ex Taq预混液、2 μl cDNA模板和引物,在Rotor-Gene Q实时PCR循环仪中进行。扩增步骤为:95 ℃5 min,然后进行40个循环,分别是94 ℃30 s,57.5 ℃ 30 s和72 ℃30 s。使用2-△△ct计算相对表达量。
1.2.4 免疫印迹 使用哺乳动物蛋白提取试剂制备总蛋白样品。使用二辛可宁酸蛋白质测定法测定蛋白质浓度。蛋白质样品(每个样品60 μg)通过8%SDS-PAGE分离并转移到硝酸纤维素膜上。然后将膜用5%脱脂奶在37 ℃下封闭1 h,然后在4 ℃下与一抗连续孵育12 h,在4 ℃与相应的二抗连续孵育4 h。最后,使用增强的化学发光法观察印迹并定量使用ChemiImager系统。 GAPDH用作内部参考。每个实验至少重复3次,每个样品重复3次。
1.2.5 细胞凋亡的检测 根据制造商的说明书,使用膜联蛋白V-异硫氰酸荧光素(FITC)/碘化丙啶(PI)测定法评估HepG2细胞的凋亡。对于流式细胞术分析,用Sorafenib处理HepG2细胞,然后收集并用冷PBS溶液洗涤2次。将细胞悬浮于400 μl结合缓冲液中,加入5 μl Annexin V-FITC孵育15 min,随后在37 ℃与10 μl PI孵育5 min。立即使用流式细胞仪和Cell Quest 4.0.2软件分析染色的细胞。
1.2.6 DNA损伤的检测 使用碱性单细胞凝胶电泳评估DNA损伤。Sorafenib处理后,将HepG2细胞在4 ℃的细胞裂解液(2.5 M NaCl,100 mM Na2EDTA,10 mM Tris,pH 10.0;1%Triton X-100和10%DMSO)中浸没1 h,然后置于电泳溶液(300 mM NaOH,1 mM Na2EDTA,pH>13)在4 ℃下放置40 min。 电泳(25 V,~300 mA)并随后用Tris-HCl(400 mM,pH 7.5)中和后,将细胞用PI(5 mg/l;Sigma-Aldrich)染色。 最后,使用CASP彗星分析1.2.2软件在荧光显微镜下以40倍的放大倍率分析彗星图像。 DNA的Olive尾矩(称为尾巴中的DNA百分比)用于评估DNA损伤的程度。
1.3 统计学分析
2 结果
2.1 Oct3/4在人肝癌细胞和人肝正常细胞中的表达
实时荧光定量PCR发现Oct3/4在人肝癌细胞Li-7、HepG2、Huh7、BEL-7405中的mRNA表达水平显著高于人肝正常细胞LX-2(P<0.05),见图1A。同时,Oct3/4在人肝癌细胞Li-7、HepG2、Huh7、BEL-7405中的蛋白表达水平显著高于人肝正常细胞LX-2,见图1B。
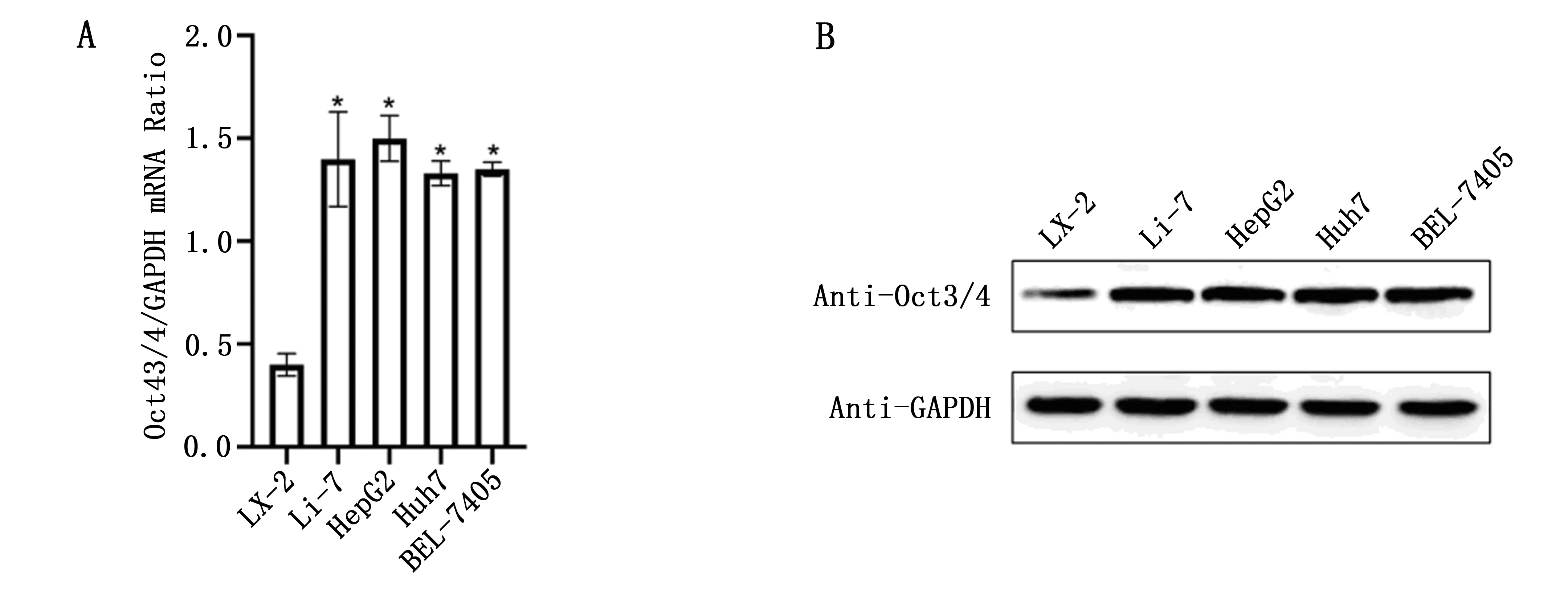
图1 Oct3/4在人肝癌细胞和人肝正常细胞中的表达
2.2 CRISPR/CAS9技术构建Oct3/4敲除的肝癌细胞系
设计gRNA靶向Oct3/4第1个外显子,见图2A。使用CRISPR/CAS9对HepG2细胞进行基因编辑后,挑取嘌呤霉素筛选出的2个单克隆细胞株进行Oct3/4表达水平的检测,发现1#号单克隆细胞株Oct3/4表达水平低于野生型WT,2#号单克隆细胞株没有表达Oct3/4,见图2B。提取1#号和2#号单克隆基因组后PCR进行测序,发现1#号基因组上的Oct3/4第1个外显子只有1条染色体缺失了2个碱基,另一条染色体并未缺失;2#号基因组上的Oct3/4第1个外显子2条染色体均存在不同程度的缺失,一条缺失2个碱基,另一条缺失4个碱基,见图2C。
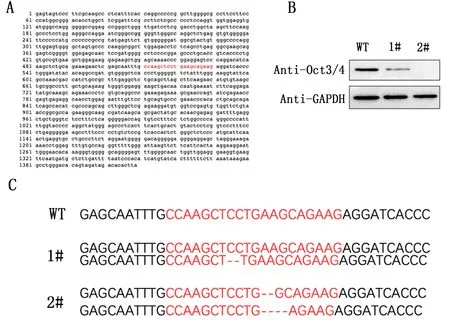
图2 CRISPR/CAS9技术构建Oct3/4敲除的肝癌细胞系
2.3 Oct3/4敲除对化疗药物作用效果的影响
选取2#号细胞株作为后续Oct3/4 KO细胞株。使用索拉菲尼处理野生型HepG2细胞和Oct3/4 KO HepG2细胞,发现Oct3/4 KO细胞株的凋亡水平显著上升(P<0.05),DNA损伤水平显著上升(P<0.05),见图3A和3B。
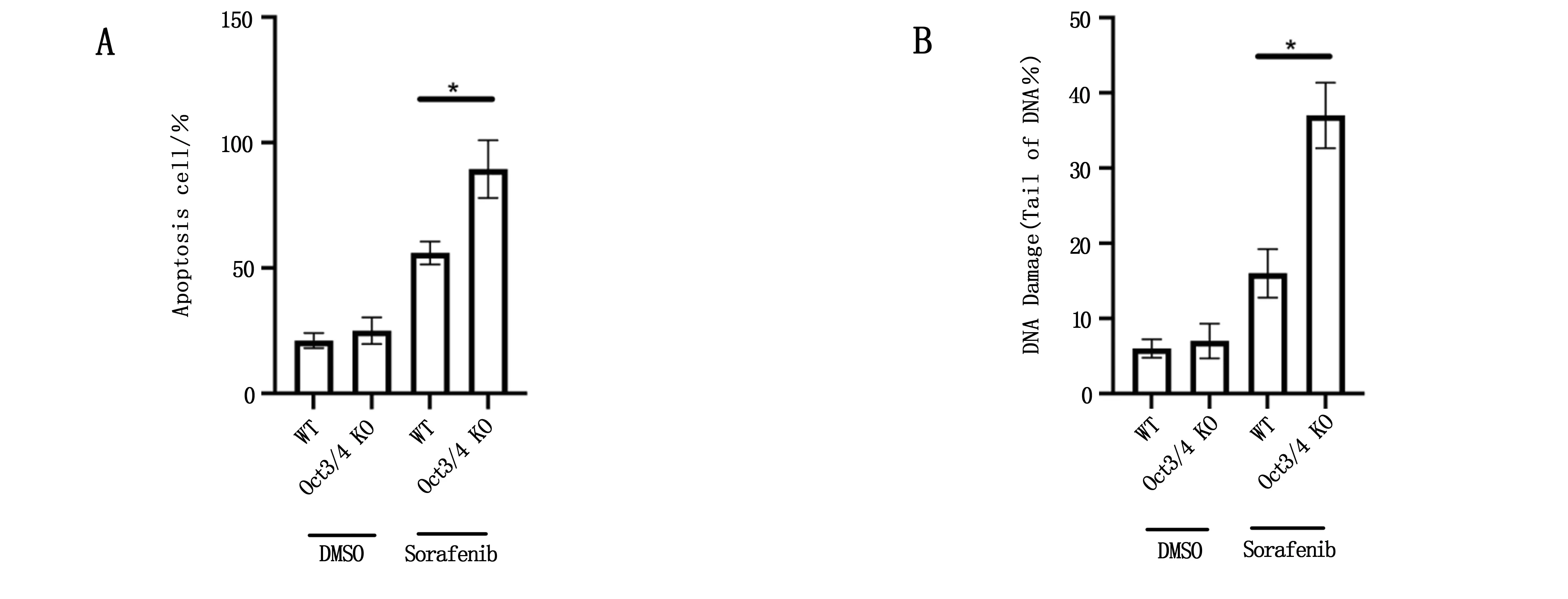
图3 Oct3/4敲除对化疗药物作用效果的影响
3 讨论
肝癌是世界范围内的常见恶性肿瘤,每年造成700000多例死亡。HCC是原发性肝癌的主要类型[7],在病因上与乙型肝炎病毒(HBV)和丙型肝炎病毒(HCV)感染、肝硬化、酒精中毒和非酒精性脂肪肝疾病(NAFLD)相关[8]。手术切除是肝癌患者的主要治疗方法。然而,由于侵袭性生长和晚期症状表现,大多数HCC患者被诊断为晚期,不符合手术治疗的条件[9]。晚期HCC患者的中位生存期约为9个月,而5年总生存率仅为10%,治疗晚期HCC患者的选择仍然非常有限,因此有必要进一步开发新的治疗方案。
OCT3/4在胚胎干细胞,生殖细胞和各种人类癌症中表达[10]。众所周知,它是干细胞多能性和自我更新的主要调节剂[11]。越来越多的证据表明[12],在包括前列腺癌、黑色素瘤和HCC在内的许多癌症中,OCT3/4表达与肿瘤起始之间存在相关性。已有研究发现OCT3/4表达在HCC对药物敏感性中起关键作用[13]。在本研究中,Oct3/4在人肝癌细胞Li-7、HepG2、Huh7、BEL-7405中的mRNA表达水平显著高于人肝正常细胞LX-2(P<0.05)。同时,Oct3/4在人肝癌细胞Li-7、HepG2、Huh7、BEL-7405中的蛋白表达水平显著高于人肝正常细胞LX-2。因此,Oct3/4可能是促进肝癌发生发展和耐药性的关键因子。
CRISPR/Cas9系统最早是在细菌和古细菌中发现的一种自适应免疫机制,可防止病毒DNA入侵[14]。在哺乳动物细胞中,CRISPR/Cas9系统已经过工程改造,可为特定基因敲除引入移码突变[15]。由于容易编程和高基因编辑功效,CRISPR/Cas9系统已越来越多地用于研究各种生物系统中基因功能的丧失[16]。在本研究中,使用CRISPR/CAS9对HepG2细胞进行基因编辑后,挑取嘌呤霉素筛选出的2个单克隆细胞株进行Oct3/4表达水平的检测,1#号单克隆细胞株Oct3/4表达水平低于野生型WT,2#号单克隆细胞株没有表达Oct3/4。测序后发现1#号基因组上的Oct3/4第1个外显子只有1条染色体缺失了2个碱基,另一条染色体并未缺失;2#号基因组上的Oct3/4第1个外显子2条染色体均存在不同程度的缺失,一条缺失2个碱基,另一条缺失4个碱基。
索拉非尼靶向多种酪氨酸激酶,包括RAF,VEGFR和PDGFR,以抑制其下游增殖和生存信号通路[17]。先前的研究表明[18],索拉非尼治疗部分通过引起肿瘤细胞DNA损伤和凋亡来抑制肿瘤生长。索拉非尼因其可耐受的安全性和可控的副作用,在临床上是一种有吸引力的分子靶向药物。但是,索拉非尼治疗HCC的临床疗效不高,只能将患者的平均总生存期延长3个月[19]。耐药性的发展被认为是导致HCC患者索拉非尼治疗失败的主要障碍[20]。为了克服索拉非尼耐药性,与其他抗癌药物尤其是靶向与索拉非尼耐药性有关的分子的药物联合开发,越来越受到重视。本研究使用Sorafenib处理野生型HepG2细胞和2# Oct3/4 KO HepG2细胞后,Oct3/4 KO细胞株的凋亡水平显著上升(P<0.05),DNA损伤水平显著上升(P<0.05)。因此,Oct3/4基因可能是HCC耐索拉菲尼的原因之一。然而,索拉非尼耐药的潜在机制很复杂,并且仍然难以捉摸。对索拉非尼耐药性的分子基础进行进一步的研究,可能为确定合理的联合疗法克服索拉非尼耐药性的新靶标提供参考。
综上所述,Oct3/4基因的敲除能够显著提高化疗药物Sorafenib促肝癌细胞凋亡和DNA损伤的作用。
