山东省6个豆梨野生群体的遗传多样性分析
2022-02-23李永涛王振猛杨庆山王莉莉刘德玺魏海霞
李永涛,王振猛,杨庆山,周 健,王莉莉,刘德玺,魏海霞
(山东省林业科学研究院 a.国家林业和草原局滨海盐碱地生态修复工程技术研究中心;b.山东耐盐碱树种研究国家长期科研试验基地,山东 济南 250014)
豆梨Pyrus calleryana为蔷薇科Rosaceae 梨属Pyrus落叶乔木,是最古老的梨属种之一,现广泛分布于华东、华南的11 个省(市、自治区)。作为我国原生梨属植物,目前是北方栽培梨的优良砧木之一,也是春季赏花、夏季观果、秋季赏叶的优质景观树种,且根、叶及果实均可入药,具有较高的经济价值和观赏价值[1-2]。山东省作为豆梨最北端的适生分布区,由于地理隔离、基因漂变等多种原因,使得植株在形态特征、生态适应性等方面都形成了一系列与环境相适应的特征,部分种质在抗逆性、观赏性等方面均具有较好的育种价值[3]。
遗传多样性是物种长期进化的结果,丰富的种质资源遗传多样性是育种的前提和物质基础[4-6]。豆梨属于典型的异质种质,野生种质经过长期自然选择,种内变异丰富,优异种质较多,因此对豆梨野生种质资源进行遗传多样性研究具有重要意义。目前有关豆梨种质资源遗传多样性的研究较少,在形态学标记方面,刘超等[7]采用主成分分析、聚类分析等方法对南方6 省78 份豆梨资源的叶片、果实等16 个表型性状进行研究,得出不同居群间存在着丰富的变异。分子标记方面,刘晶[8]利用SSR 标记技术,得出了浙江省豆梨资源保持着较高的遗传多样性水平,并提出了资源保护措施。形态学标记虽具有重要地位,但很难确切地反映不同种质间的遗传变异特性[9-10]。AFLP分子标记技术作为种质资源遗传多样性和遗传结构研究最有效的方法之一[11],现已广泛用于梨[12]、苹果[13]、楸树[14]等多年生木本植物。为明确山东省野生豆梨群体分布的遗传多样性特点,本研究运用AFLP 分子标记技术对采自不同区域分布的61 份豆梨种质进行了遗传多样性及其亲缘关系分析,以期为野生豆梨优良种质资源的收集、保存及育种材料的选择提供理论依据。
1 材料和方法
1.1 试验材料
试验材料保存在山东省林科院盐碱地造林试验站豆梨资源圃内,该材料于2019年期间采集于山东省五莲山、九仙山、横山、昆嵛山、蒙山及崂山6 个地区,采集时同一区域各单株间隔150 m以上,通过嫁接方式保存。其中,五莲山3 份(编号:C1~C3),九仙山10 份(编号:C4~C13),横山9 份(编号:C14~C22),昆嵛山7 份(编号:C23~C29),蒙山23 份(编号:C30~C52),崂山9 份(编号:C53~C61),共计61 份。
采集新梢顶端的新鲜幼嫩叶片,放入自封袋中用硅胶干燥保存。
1.2 试验方法
1.2.1 DNA 的提取
豆梨叶片DNA 的提取采用CTAB 法,提取后采用0.8%的琼脂糖凝胶电泳检测,-20℃保存备用。
1.2.2 AFLP 扩增及检测
AFLP 扩增过程中的酶切连接、预扩增反应、选择性扩增反应体系及条件等试验流程严格按照李永涛等[14]的方法进行。其中限制性内切酶选用EcoRI 和MseI 两种,酶切与连接同时进行,在37℃保温5 h 后,8℃保温4 h,于4℃过夜;预扩增反应后将预扩增产物稀释15 倍后再进行选择性扩增。
PCR 扩增在Gene Amp PCR System 9600 扩增仪上进行,内切酶和连接酶购自New England Biolabs 公司,通用引物由北京鼎国昌盛生物技术有限公司合成。扩增产物在4%聚丙烯酰胺变性凝胶上电泳。
1.3 数据统计分析
选取70~500 bp 的DNA 条带,在同一迁移位置采取0/1 赋值记带,有条带记为“1”,无条带记为“0”。遗传多样性的计算采用POPGENE version 软件进行,计算参数包括多态位点百分率(PPB)、有效等位基因数(Ne)、观测等位基因数(Na)、Nei’s 多样性指数(H)及Shannon’s信息指数(I);遗传相似性系数利用NTSYSpc-2.11F 计算;依据相似性系数对各群体进行UPGMA 聚类分析[15]、主效应分析PCA(principal component analysis)和分子方差分析(AMOVA)。
2 结果与分析
2.1 AFLP 扩增结果分析
采用筛选的8 对AFLP 引物对61 个野生豆梨单株资源进行了扩增(表1),8 对引物组合共扩增出清晰条带1 344 条,其中多态性条带1 340 条,占总条带数的99.75%,平均每对引物扩增出168条带,多态性条带比率在99.29%~100.00%之间,说明筛选的8 对引物扩增多态性较高。同时,扩增条带数最多的引物组合为E-AGC/M-CTC,共扩增出200 条带,扩增效率最高(100%);而扩增条带数最少的引物组合为E- AGG/M-CAG,扩增出141 条带,扩增效率最低(99.29%)。
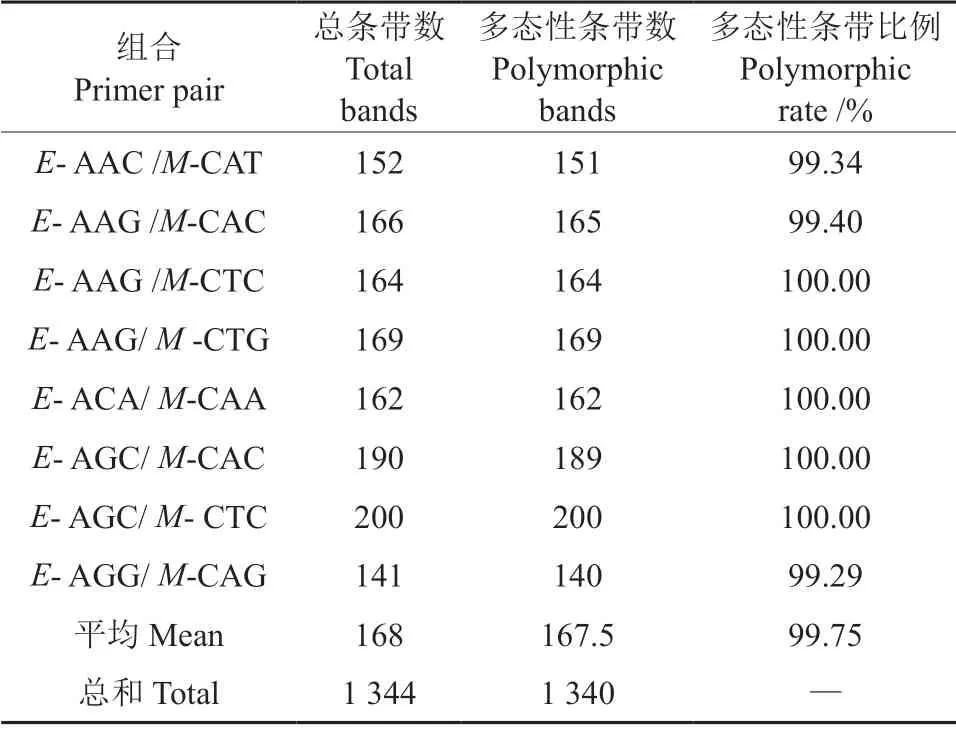
表1 8 对引物组合扩增的AFLP 条带结果Table 1 Amplified bands with eight pairs of AFLP primers
由图1可以看出,引物组合P-ACG/M-CAC的图谱显示扩增多态性好,条带清晰,易于分辨,说明所选用的引物组合适于豆梨资源的鉴别。
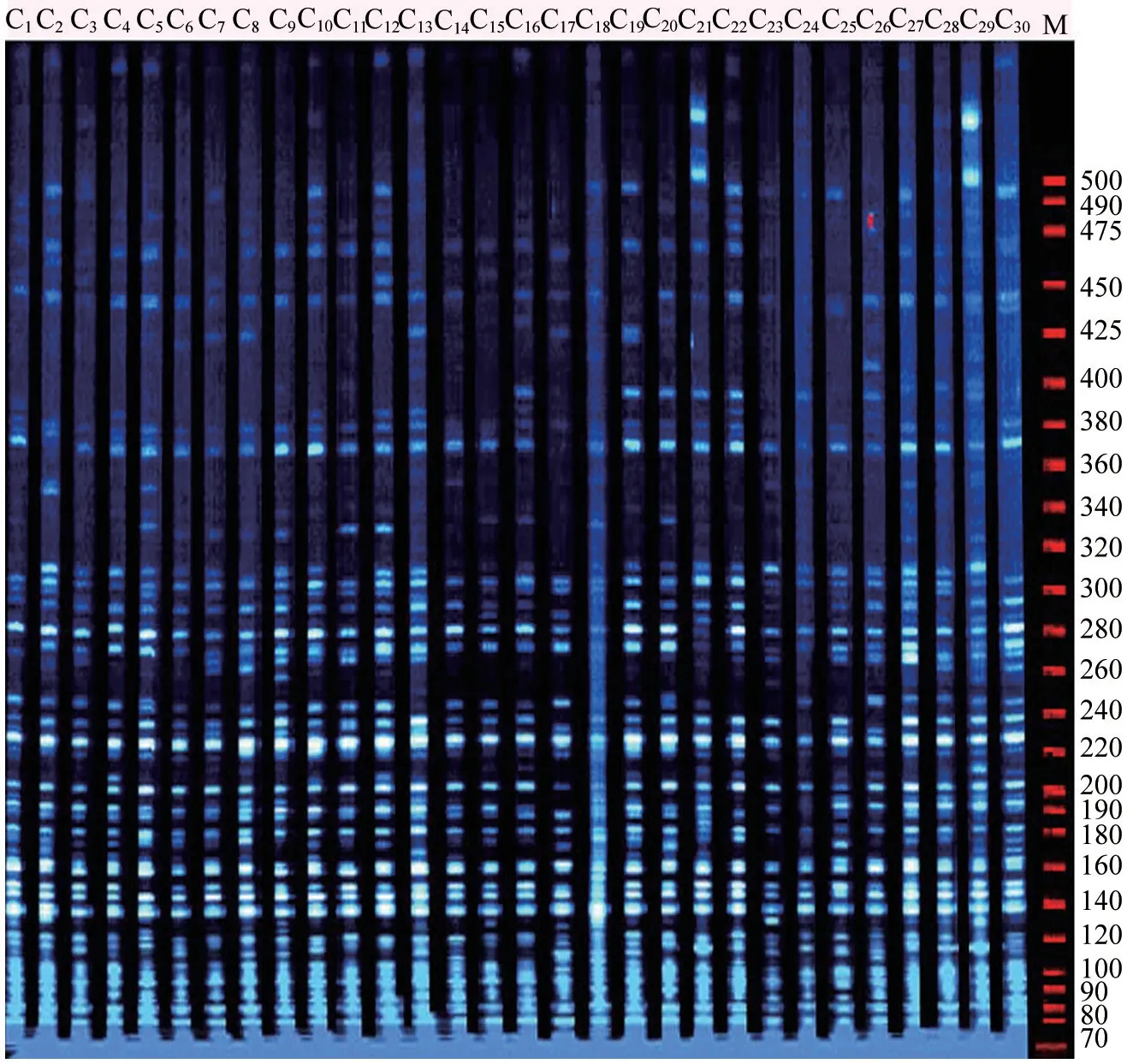
图1 引物组合E-AAC/M-CAT 对C1~C30 号豆梨材料的AFLP 扩增图谱Fig.1 AFLP fingerprinting of the primer pair E- AAC /M-CAT for No.C1-C30 tartary Pyrus calleryana accessions
2.2 群体遗传多样性与遗传结构分析
根据供试材料的采集区域,将61 个野生豆梨单株划分为6 个群体。通过对6 个豆梨群体的遗传多样性分析(表2),各群体资源在3~23 株不等,其中M 区(蒙山)资源最多(23 株),W区(五莲山)和K 区(昆嵛山)资源最少(3 株和7 株)。虽然K 区资源份数较少,但Nei’s 多样性指数(H=0.133 3)和Shannon’s信息指数(I=0.206 6)均较高,表明种质资源数量的多少与遗传多样性无直接关联性。
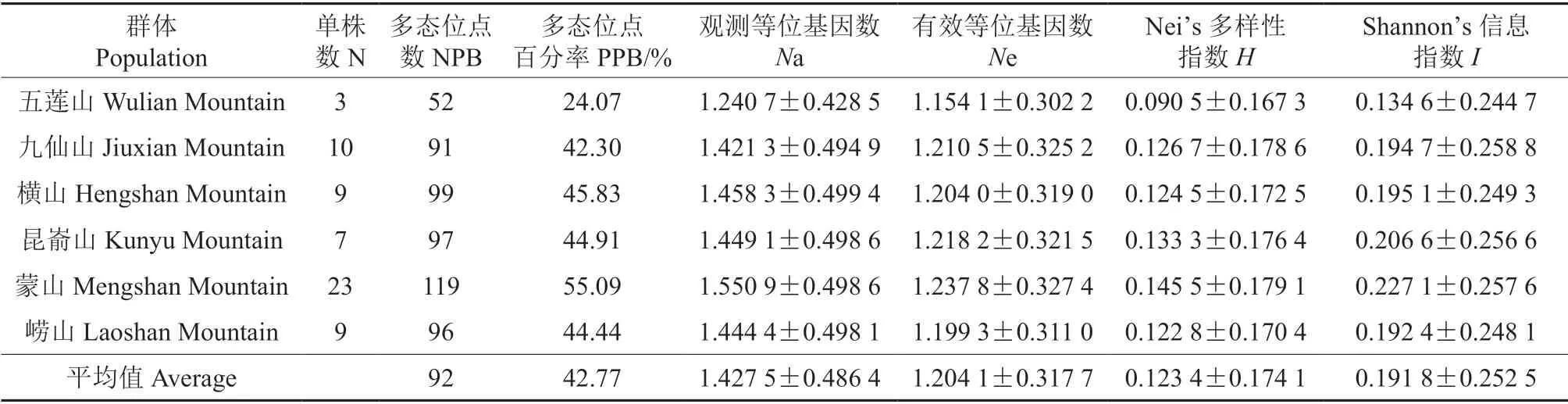
表2 6 个豆梨野生群体的遗传多样性分析Table 2 Analysis result of genetic diversity of six populations of Pyrus calleryana
在群体水平上,豆梨的多态位点百分率(PPB)的变化范围为24.07%~55.09%,平均值为42.77%;观测等位基因数(Na)为1.240 7~1.550 9,平均值为1.427 5;有效等位基因数(Ne)为1.154 1~1.237 8,平均值为1.204 1;Nei’s 多样性指数(H)为0.090 5~0.145 5,平均值为0.123 4;Shannon’s 信息指数(I)为0.134 6~0.227 1,平均值为0.191 8。其中Na、Ne、H和I指标均以蒙山和昆嵛山群体的最大,五莲山群体的最小。通过AMOVA 分析(表3)显示,12.53%的变异来自群体间,87.47%的变异来自群体内,这与群体的遗传多样性分析结果基本一致。
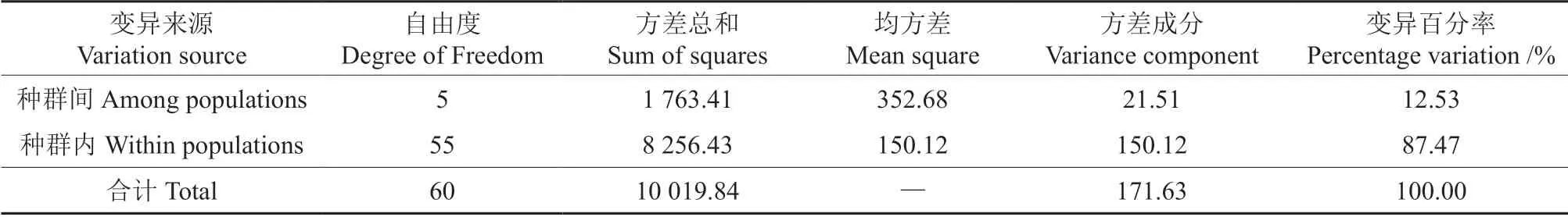
表3 豆梨种群的分子方差分析(AMOVA)Table 3 Molecular variance analysis (AMOVA) of Pyrus calleryana
2.3 特异性条带分析
通过对61 个野生豆梨单株AFLP 标记的分析,8 对引物组合共产生特异性条带179 条。不同引物组合扩增出的特异性条带数存在一定差异,其中引物组合E-AGG/M-CAG 产生的特异性条带最多,共扩增出32 条特异性条带。引物组合E-AGC/M-CAC 产生的特异性条带数最少,共扩增出14 条特异性条带。而不同单株间产生的特异性条带也不同,其中产生特异性条带数最多的是C13号单株,共有12 条;其次是C2和C54单株,均有8 条特异性条带;而产生特异性条带最少的单株为C4、C7、C20、C22、C40、C42、C44、C47、C50、C57和C59,共11 株,均有1 条特异性条带;此外还有7 个单株未扩增出特异性条带,通过特异性条带分析可以对88.52%的野生豆梨单株进行区分鉴别。
2.4 遗传相似性分析及聚类分析
通过计算61 个野生豆梨单株间的遗传相似性系数,不同单株间的遗传相似性系数介于0.737 8~0.884 3 之间,平均值为0.806 8。其中,单株间遗传相似性系数最大是C53号单株与C54号单株,达到0.884 3,且两个单株均分布于崂山地区,表明二者的遗传距离最近,差异性最小。C12号单株与C39号单株间的遗传相似性系数最小,为0.737 8,二者分别位于九仙山和蒙山地区,表明两个单株间的遗传距离最远,差异性最大。同时,C6号单株与其它单株间的平均遗传相似性系数最大,为0.826 7,而C39号单株与其他单株间的遗传相似性系数平均值最小,为0.770 9,说明C6号单株与其他单株间具有较高的遗传相似性,而C39号单株与其他单株间的遗传相似性较小。
通过UPGMA 法对61 个单株进行聚类分析(图2)得出,在遗传相似系数0.806 处,可将6 个群体分为5 组。其中,第Ⅰ组包括1 个单株,C13位于九仙山;第Ⅱ组包括12 个单株,其中C32、C33、C35、C37~C41、C46号9 个单株位于蒙山,C55、C56、C61号3 个单株位于崂山;第Ⅲ组包括2个单株,其中C21号单株位于横山,C29号单株位于昆嵛山;第Ⅳ组包括7 个单株,其中C31号单株位于蒙山,C53、C54、C57~C60号6 个单株位于崂山;第Ⅴ组包括39 个单株,涵盖五莲山、九仙山、横山、昆嵛山和蒙山5 个群体的部分单株。其中,C1~C3号3 个单株位于五莲山,C4~C12号9 个单株位于九仙山,C14~C20、C22号8 个单株位于横山,C23~C28号6 个单株位于昆嵛山,C30、C34、C36、C42~C45、C47~C52号13 个单株则位于蒙山地区。而当在遗传相似性系数0.824 处分割时,可将第Ⅴ组分为8 个亚组,其中第1 亚组包括C1、C2号2 个单株,而第2 亚组仅C3号1 个单株,且第1、2 亚组均分布于五莲山地区;第3 亚组包括15 个单株,其中C4、C6~C12号8 个单株分布于九仙山地区,C14~C16、C18~C20和C22号7个单株分布于横山地区;第4 亚组包括6 个单株,为C23~C28号6 个单株,均分布于昆嵛山地区;第5 亚组与第6 亚组各包括1 个单株,为C5号和C30号单株,各分布于九仙山和蒙山地区;第7 亚组包括12 个单株,分别为C34、C36、C42~C45及C47~C52号单株,均位于蒙山地区;第8 亚组包括C17号1 个单株,分布于横山地区。聚类分析结果表明,61 个野生豆梨单株的分组情况与地理分布位置并不完全一致,部分来源相同的单株未完全聚在同一类群中。
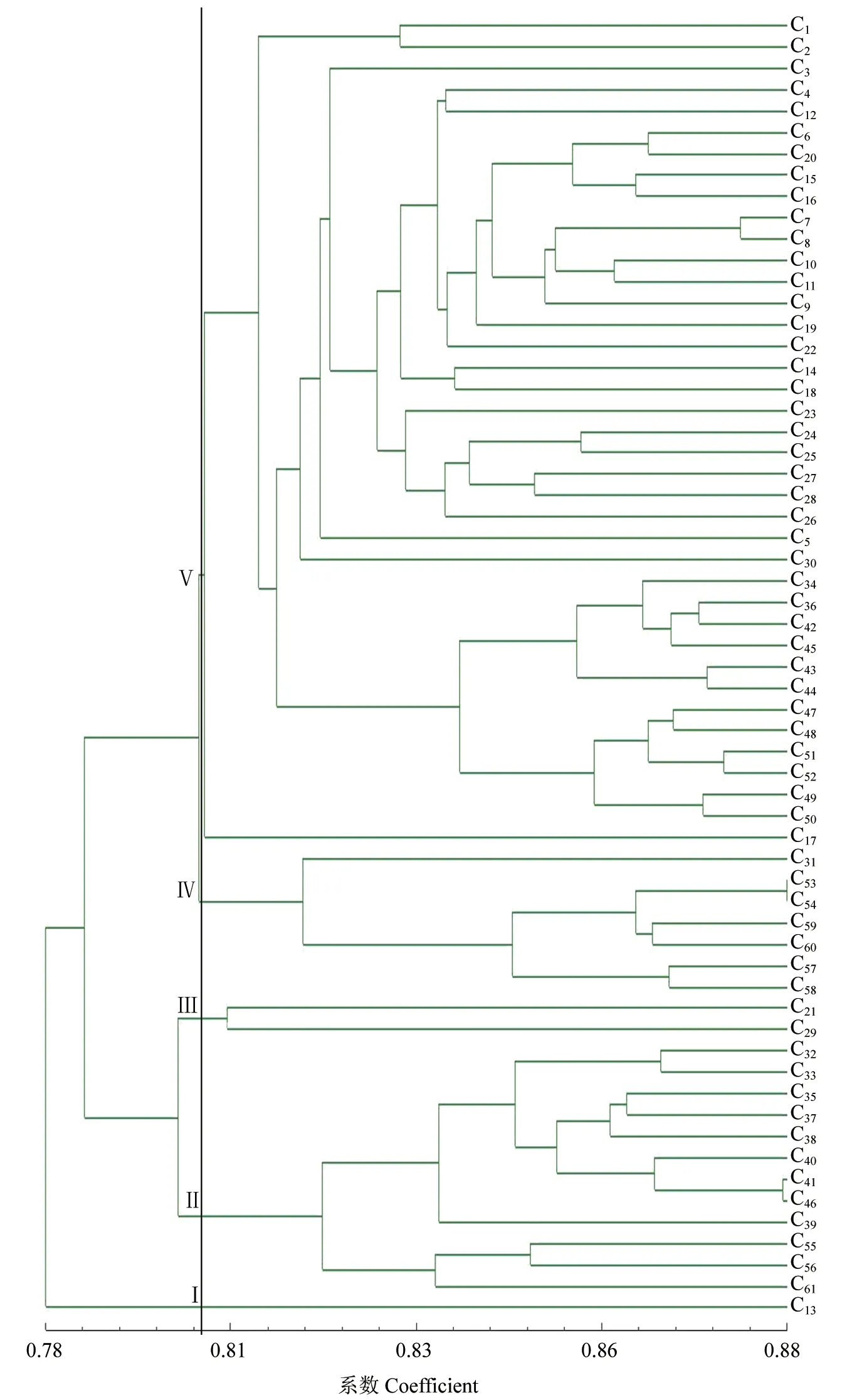
图261 份豆梨单株的AFLP 聚类结果Fig.2 Dendrogram of 61 accessions of Pyrus calleryana germplasm with AFLP markers
2.5 主效应分析
通过主效应分析(图3),根据距离位置远近61 个野生单株可划分为5 个主要类群,其中第1个类群包括28个单株,分别来源于五莲山、九仙山、横山、昆嵛山、蒙山;第2 个类群包括2 个单株,来源于横山和昆嵛山地区;第3 个类群包括12 个单株,来源于蒙山、崂山地区;第4 个类群包括12 个单株,来源于蒙山地区;第5 个类群包括7个单株,来源于蒙山和崂山地区。从图3可以看出,6 个群体之间虽存在一定程度的基因混杂,但主效应分析结果与聚类分析的结果基本一致。
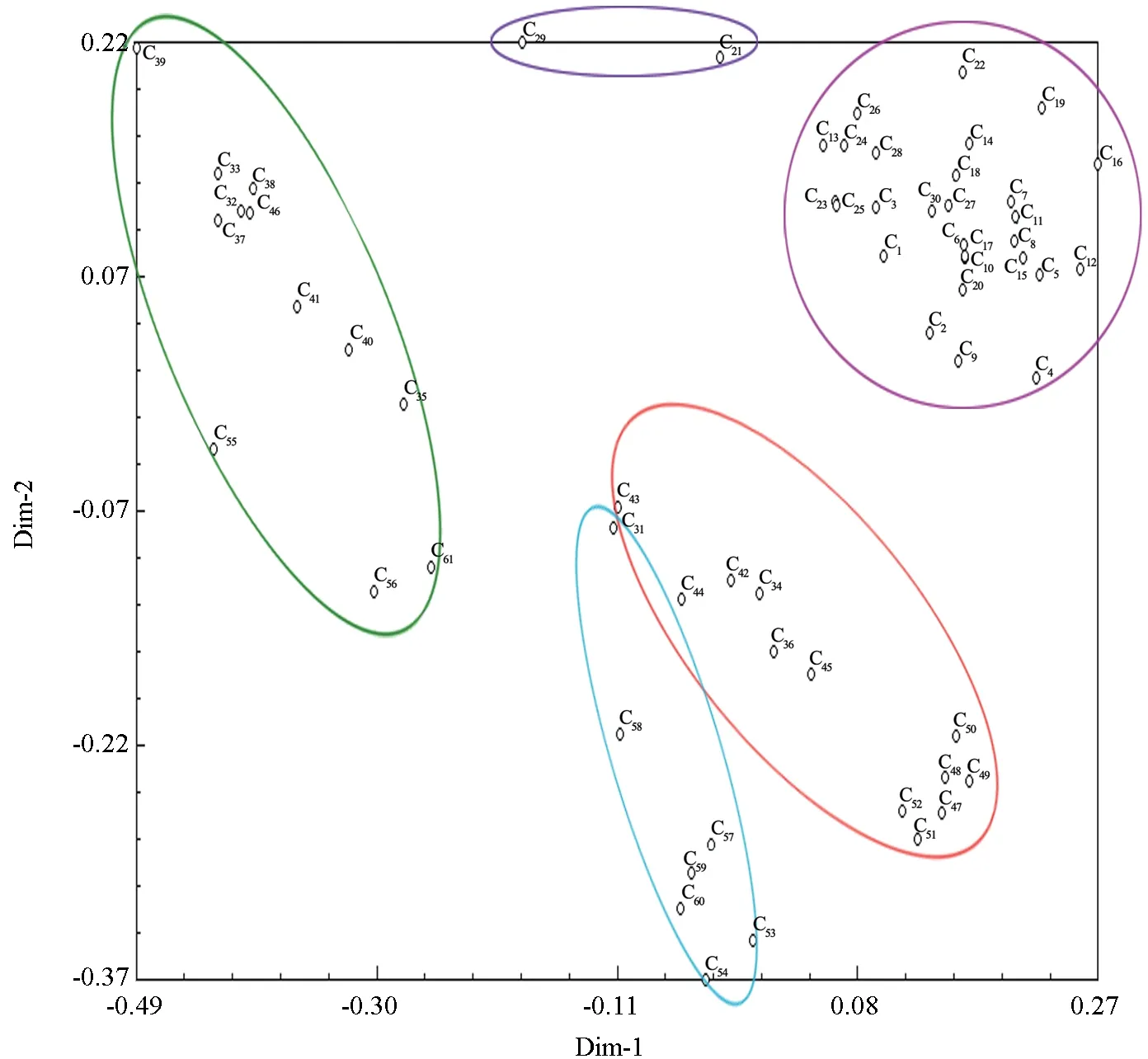
图3 豆梨种质材料的AFLP 主效应分析Fig.3 Principal components analysis on Pyrus calleryana germplasms using AFLP data
3 讨论与结论
3.1 讨 论
本研究基于AFLP 标记对山东省61 份豆梨种质资源进行遗传多样性分析,共获得清晰多态性条带1 340 条,多态性比例达99.75%,比王艳芳[16]对27 份梨资源中扩增获得的多态性条带比例(92.20%)高,比董美超等[17]对90 份鳄梨种质扩增获得的多态性条带比例稍低(99.83%),但多态性比例均达到了90%以上。同时,本研究8对引物组合共扩增出179 条特异性条带,通过这些特异性条带可将88.52%的野生豆梨单株区分开来,表明AFLP 标记能很好地反映61 份野生豆梨种质的遗传多样性信息,尤其是E-AAG/M-CTC、E-AAG/M-CTG 等5 对引物组合扩增条带稳定、多态性最高,可广泛用于其它豆梨种质的遗传多样性分析及亲缘关系的鉴定评价。
种质资源遗传多样性研究是种质资源利用的基础,且物种的遗传多样性受多种因素影响[18]。本研究得出6 个野生豆梨群体的有效等位基因数、Nei’s 多样性指数及Shannon’s 信息指数分别为1.204 1、0.123 4 和0.191 8,说明在居群水平上豆梨存在较丰富的遗传变异度和较高的遗传多样性水平。通过ANOVA 分析显示,群体间变异达到12.53%,群体内变异达到87.47%,且遗传变异主要来源于群体内,这与刘晶[8]利用SSR 标记对中国豆梨种质资源的研究结果类似。这种居群间较低的遗传分化可能与其虫媒授粉特性及异交为主的混合交配等植物学特性[19]紧密相关。此外,豆梨种子的传播在减少居群间遗传分化方面也起着非常重要的作用。因为,豆梨种实较小,味道香甜,是鸟类等动物的主要食物之一,极易造成迁徙过程中的种子携带,从而减少了地理隔离的影响[20-21],因此在花粉和种子两个方面可以同时形成远距离传播。
基于遗传相似性和聚类分析结果得出,豆梨资源的聚类结果与其群体的地理分布格局具有一定的相关性,在遗传相似系数0.806 处可将61 份资源分为5 大组,其中第Ⅰ组有1 个单株,来源于九仙山,说明该单株亲缘关系较其他单株较远;第Ⅱ组共有12 个单株,来源于蒙山和崂山;第Ⅲ组共有7 个单株,来源于横山和昆嵛山;第Ⅳ组共有7 个单株,来源于蒙山和崂山,说明以上各组内同一地区或毗邻地区资源的亲缘关系相对较近;第Ⅴ组包括39 份资源,占总资源的63.93%。在相似系数为0.826 时,又可将其分为8 个亚组,其中第2、5、6、7 亚组均包括1 份资源,说明这4 份资源的亲缘关系较其它单株较远;第1 亚组共有2 份资源,均来源于五莲山地区;第4 亚组共有12 份资源,均来源蒙山地区;第3 亚组共有15份资源,来源于九仙山和横山。说明亚组内资源之间亲缘关系相对较近,位于同一地区的资源更倾向于聚为一类,而从地理位置上看,五莲山、九仙山、横山、蒙山均位于鲁南地区,隶属泰沂山脉,距离相对较近,气候环境类似,生态因子差异较小所致。AFLP 聚类结果分析也未能将61份资源完全按照地理来源聚类,不同群体之间存在一定的交叉,部分单株之间距离虽然较远,但仍被聚为一个类群,如蒙山与崂山、横山与昆嵛山的部分单株,这说明豆梨的授粉特性、种子散布模式和地理分布等还不能解释其居群结构和遗传分化的全部原因。此外,采集区域的气候环境等生境因素的影响以及资源采集时可能造成的豆梨与其它梨属资源混淆等原因也具有一定关系。需要指出的是,本研究供试材料取自山东省6 个地区,地域性相对较窄,供试样本偏少,不利于全面综合分析豆梨的遗传多样性。今后有必要对我国适生分布区的豆梨资源开展大尺度系统研究,以便为更加科学合理地利用其资源奠定基础。
3.2 结 论
综上,61 份野生豆梨种质资源的遗传基础总体差异较小,亲缘关系较近。来自九仙山和蒙山地区的种质资源遗传多样性相对较丰富,C12号与C39号单株较为特殊,在杂交育种时可重点利用。此外,山东省野生豆梨群体间存在一定的基因交流,各群体内遗传分化程度大于群体间,且群体间的地理来源和遗传变异程度并没有严格的相关性,这为进一步了解山东省野生豆梨的资源现状、制定合理的育种方案提供了理论依据。
