香椿半同胞家系在川中丘陵区生长变异及早期选择
2022-02-23肖兴翠王树山杨勇智杨槟豪徐志伦漆峥嵘
肖兴翠,王树山,杨勇智,曾 军,杨槟豪,徐志伦,漆峥嵘
(1.四川省林业科学研究院,四川 成都 610081;2.蓬溪县金果子水果种植专业合作社,四川 遂宁 629100;3.蓬溪县自然资源和规划局,四川 遂宁 629100)
香椿Toona sinensis是楝科香椿属落叶乔木,生长迅速,树干通直,木材色泽红润、花纹美丽,且具有香味,有“中国桃花心木”之称[1]。香椿是我国特有的速生珍贵用材树种,也是四川乡土珍贵用材树种,较桢楠、柏木等珍贵树种速生,18~20 a 可达到大径材标准,是目前世行造林、储备林建设的主要造林树种之一。从2000年开始四川省就把香椿作为优先发展的6 个珍贵用材树种之一,通过退耕还林在川中丘陵区、川南地区大面积种植,经过近20年的发展,目前四川的香椿人工林面积已有近6.7 hm2。然而前期香椿造林因良种缺乏、种苗质量差,造林效果一直不理想。
国内尚无香椿良种,选育适合各香椿造林区域的良种成为香椿育种研究的重点。在香椿良种选育方面,通过家系选择,刘正周等[2]从来自4个省的9 个种源中选择出3 个适合河南南阳的优良种源,宋志姣[3]从9 个种源中分别选择出适合广西南宁的2 个芽用种源和2 个材用种源,张海燕[4]从来自14 个省的29 个香椿种源中选择出2个适合福建邵武的优良种源,并从70 个香椿优树半同胞子代家系中选择出11 个早期优良家系[5],左继林等[6-7]从11 个香椿种源中选择出福建霞蒲和湖南洞口2 个适合江西瑞昌的早期优良种源,高楠等[8]从12 个种源70 个香椿优树子代中选择出4 个适合福建永春和华安的早期优良家系,胡继文等[9]从49 个香椿无性系中选择出10 个适合河南造林的早期优良无性系。
四川香椿良种选育起步较晚,从“十二五”开始起步,先后认定了“蓬溪香椿母树林”“筠连香椿母树林”和“合江香椿母树林”3 个初级良种,但其材积遗传增益均有限。四川省林科院香椿育种团队从“十三五”开始,先后进行了省内外香椿优树选择、优良种质资源收集、分家系育苗,并进行了苗期生长比较与优良家系选择[10-11],2017年开始先后在秦巴山区[12]、川中丘陵区和川南地区建立了区域化试验林,旨在选育出香椿良种,为不同区域香椿造林提供良种。本研究以栽植在蓬溪县的45 个香椿半同胞家系为研究对象,通过对造林4年的生长变异进行分析,开展优良家系早期选择,为川中丘陵区香椿良种选育提供参考。
1 材料与方法
1.1 试验地概况
试验地位于四川省遂宁市蓬溪县吉星镇,与红椿良种选育试验林位于相同地块,地理位置是东经105°46′44″,北纬30°37′31″,海拔430~440 m。蓬溪县属中亚热带温湿季风气候区,气候温和,雨量充沛,无霜期长,雨热同季,年平均气温16.8℃,年平均降水量929.5 mm,年均日照数1 471.7 h。试验地造林前为抛荒田地,土壤为紫色土,有效土层厚度≥60 cm,pH 值8.1,表层土有机质含量13.3 g·kg-1,水解氮80.9 mg·kg-1,有效磷4.8 mg·kg-1,速效钾133.0 mg·kg-1,土壤肥力中等。
1.2 试验材料
参试的44 个香椿优树半同胞家系及对照主要来源于川中丘陵区的蓬溪和乐至及川南地区的筠连和高县,引进福建福安、霞浦和广西灌阳、兴安以及陕西白河的优树半同胞家系,对照为蓬溪本地种源,不同家系种源所在地及其气候情况见表1。

表1 香椿不同半同胞家系所在地理位置及气候条件Table 1 Geographical location and climatic conditions of different Toona sinensis half-sib families
1.3 造林方法
44 个香椿优树半同胞家系均是分单株采种,分家系育苗,培养轻基质容器苗。44 个香椿家系造林时均采用1年生轻基质容器苗,平均苗高20 cm,地径0.3~0.5 cm,对照为蓬溪种源混合种子培育的1年生裸根苗,造林时截干成80 cm,地径≥1 cm。2017年3月造林,采用随机区组设计,5 株单行小区,4 次重复。造林前1年的年底机械全垦整地,造林时小穴整地栽植,株行距为2 m×3 m,栽后浇水定根,并覆盖薄膜保湿。造林1~3 a 每年除草2~3 次,造林当年7月初施配方肥200 g·株-1,造林第2、3年每年5月前分别施配方肥250、500 g·株-1,配方肥为根据土壤养分状况和香椿的需肥特性配置,氮、磷、钾质量比为1.6∶1.5∶1.0。
1.4 数据调查与分析
造林当年年底测量树高,从第2年开始每年年底测量胸径、树高。采用Excel 2010 软件进行数据统计,采用SPSS 19.0 软件进行方差分析及LSD 多重检验。其中单株材积采用四川省和滇西北阔叶树二元立木材积表,V=5.275 071 6×10-5D1.9450324H0.9388533[13]。
遗传变异系数:C=S/X×100%。式中:S为标准差;X为某一性状的群体平均值。
家系遗传力:h2=(MSv-MSt)/MSv×100。式中:MSv 为处理间均方;MSt 为处理内均方[14-15]。
遗传增益:△G=h2Si/Xi。式中:h2为性状的遗传力;Si为家系选择差;Xi为个体平均数[16]。
2 结果与分析
2.1 不同家系生长与种源环境相关性分析
对香椿不同家系造林4 a 的胸径、树高和单株材积3 个生长指标与优树所在地理位置及气候进行相关性分析,结果见表2。

表2 香椿家系4年生生长指标与环境因子相关性分析†Table 2 Correlation analysis between growth indexes and environmental factors of the 4-year-old Toona sinensis families
由表2可见,4年生香椿家系单株材积与胸径和树高、胸径与树高之间均有极显著相关性。香椿家系生长与种源地的地理位置之间,胸径、树高、单株材积与纬度均呈极显著负相关关系,与海拔呈显著负相关,与经度呈负相关关系,表明在地理位置中,影响家系生长的因素大小为纬度>海拔>经度,在选择香椿种源时,可选择低纬度、低海拔、经度小的地区的种源对川中丘陵区香椿优良家系选择有利。香椿家系生长与种源地的气候因子之间,胸径、树高、单株材积与年均气温和年均降水量均呈极显著的正相关关系,与无霜期呈显著的正相关关系,表明可选择年均气温高、降水量大、无霜期长的地区的种源对川中丘陵区优良香椿家系选育有利。
2.2 不同家系生长变异与比较
44个香椿优树半同胞家系及对照4年生胸径、树高、单株材积和高径比方差分析见表3。
由表3可见,香椿不同家系间胸径、树高、单株材积和高径比均有极显著差异(P<0.01)。胸径、树高的遗传力略大于单株材积,均在0.60以上,表明不同香椿家系在4年生时生长受遗传控制较大。

表3 香椿不同家系4年生生长性状方差分析Table 3 Variance analysis of growth traits of the 4-year-old Toona sinensis families
2.2.1 不同家系胸径生长变异与比较
44 个香椿优树半同胞家系及对照4年生胸径生长变异见表4。不同香椿家系4年生平均胸径为7.14 cm,大小在2.36~10.47 cm 之间,高于平均值的有28 个家系,高于对照的有29 个家系。其中以蓬溪9 号的胸径最大,大小在7.38~10.22 cm之间,平均为8.79 cm,年均胸径生长量为2.20 cm,遗传增益达16.51%,显著(P<0.05)高于胸径≤6.51 cm 的13 个家系,与胸径在6.66~8.67 cm 之间的31 个家系间没有显著差异。其次是筠连66 号,平均胸径为8.67 cm,年均胸径生长量为2.17 cm,遗传增益为15.25%,显著高于胸径≤6.41 cm 的12 个家系,与蓬溪9 号及胸径在6.51~8.46 cm 之间的31 个家系之间没有显著差异。排在第3 位的有筠连50 号和57 号、福安112号、乐至10 号及灌阳3 号5 个家系,胸径在8.25~8.46 cm 之间,年均胸径生长量为2.02~2.12 cm,遗传增益在11.06%~13.16%之间,显著高于胸径≥5.99 cm 的8 个家系,5 个家系之间及其与蓬溪9 号、筠连66 号和胸径在6.35~8.12 cm 之间的30 个家系间没有显著差异。不同香椿家系胸径平均变异系数为21.97%。有23 个家系的变异系数小于15%,这些家系的胸径生长较为均匀;有22个家系的变异系数大于15%,表明这些家系的胸径生长有一定的个体分化。

表4 4年生香椿不同家系胸径生长变异†Table 4 DBH variation of the 4-year-old Toona sinensis families
2.2.2 不同家系树高生长变异与比较
44 个香椿优树半同胞家系及对照4年生树高生长变异见表5。不同家系4年生香椿的平均树高为7.54 m,大小在3.12~11.50 m 之间,高于平均值的有29 个家系,高于对照的有25 个家系。其中以福安112 号的树高最大,大小在7.45~11.30 m之间,平均为9.13 m,年均树高生长量为2.28 m,遗传增益为14.69%,显著(P<0.05)高于树高≤7.16 m 的13 个家系,与树高在7.39~8.91 m 之间的31 个家系间没有显著差异。其次是乐至10 号,平均树高为8.91 m,年均树高生长量为2.23 m,遗传增益为12.66%,显著高于树高≤6.93 m 的12个家系,与福安112 号及胸径在7.16~8.85 cm之间的32 个家系之间没有显著差异。排在第3 位的是蓬溪4 号和筠连66 号2 个家系,树高在8.82~8.85 m 之间,年均树高生长量为2.21 m,遗传增益为11.76%~12.08%,显著高于树高≥6.81 m的11 个家系,2 个家系之间及其与福安112 号、乐至10 号和树高在6.93~8.54 m 之间的30 个家系间没有显著差异。不同香椿家系树高平均变异系数为18.26%,有28 个家系的变异系数小于15%,这些家系的树高生长较为均匀;有17 个家系树高的变异系数大于15%,表明这些家系的树高生长有一定的个体分化。
表5 4年生香椿不同家系树高生长变异Table 5 Height variation of the 4-year-old Toona sinensis families
2.2.3 不同家系单株材积生长变异与比较
44 个香椿优树半同胞家系及对照4年生单株材积生长变异见表6。4年生香椿不同家系的平均单株材积为0.017 8 m3,高于对照,大小在0.001~0.048 m3之间,高于平均值的家系有27个,高于对照的家系有29 个。其中以福安112 号、乐至10 号、蓬溪9 号和筠连66 号4 个家系的单株材积最大,平均为0.027 8~0.028 0 m3,遗传增益为33.63%~34.48%,显著(P<0.05)高于单株材积≤0.016 3 m3的15 个家系,与单株材积在0.015 0~0.024 5 m3之间的28 个家系间没有显著差异。不同香椿家系单株材积平均变异系数为52.71%,除了筠连15 号的变异系数小于15%外,其余家系单株材积的变异系数均大于15%,表明这些家系4年生的单株材积生长存在一定的个体分化。

表6 4年生香椿不同家系单株材积生长变异Table 6 Individual volume variation of the 4-year-old Toona sinensis families
2.3 优良家系早期选择
单株材积是胸径和树高累积效应的结果,因此以单株材积为主,胸径和树高为辅,对4年生香椿家系进行优良家系早期选择。将44 个香椿半同胞家系及对照分为Ⅰ、Ⅱ、Ⅲ、Ⅳ 4 个等级,遗传增益Ⅰ>Ⅱ>Ⅲ>Ⅳ,定为优、良、中、差4等,结果见表7,不同等级包含的香椿家系见表8。

表7 香椿优良家系早期选择与遗传增益估算Table 7 Early selection and genetic gain estimation of superior families of Toona sinensis

表8 不同等级包含的香椿家系Table 8 Families of Toona sinensis with different grades
由表7~8 可见,Ⅰ等家系包含了福安112 号、乐至10 号、蓬溪9 号、筠连66 号4 个家系,入选率为8.89%,其中乐至10 号、蓬溪9 号2 个家系来自川中丘陵区,筠连66 号来自川南地区,福安112号来自福建福安,4 个家系的胸径、树高、单株材积平均值分别为8.54 cm、8.84 m 和0.027 9 m3,选择差分别为1.40 cm、1.30 m 和0.010 0 m3,遗传增益分别为13.95%、12.01%、33.84%,胸径、树高和单株材积的遗传增益均较大,为优等家系。Ⅱ等家系5 个,入选率为11.11%,其中筠连50、57号2 个家系来自川南地区,蓬溪1 号来自川中丘陵区,灌阳3 号和兴安4 号来自广西桂林,5 个家系的平均胸径、树高、单株材积分别为8.19 cm、8.10 m 和0.023 5 m3,选择差分别为1.05 cm、0.55 m和0.005 7 m3,遗传增益分别为10.51%、5.13%、19.23%,胸径和树高均有一定的增益,单株材积平均遗传增益大于10%,为良等家系,有一定的生长潜力,有待继续观测。Ⅲ等家系包括17个家系,胸径、树高、单株材积的选择差均为正值,但遗传增益均小于10%,为中等家系。Ⅳ等家系19 个,胸径、树高、单株材积的选择差和遗传增益均为负值,长势差,平均胸径、树高和单株材积均低于平均值,为差等家系。
2.4 优良家系生长规律
对选择出的福安112 号、乐至10 号、蓬溪9 号、筠连66 号4 个早期优良家系的生长规律进行研究,其胸径和树高的生长规律见图1。
由图1可见,随着树龄的增大,不同家系的胸径逐年增大,不同家系间的差异逐年变小,其中乐至10 号的胸径较其他3 个家系有减小的趋势。不同家系的树高处于快速生长阶段,随着树龄的增大而增大,在2年生时不同家系间的差异略微变大,3年生和4年生略微变小,筠连66 号的树高在2年生时最小,到3年生时与乐至10 号和福安112 号无明显差异,到4年生时,福安112 号的树高最大。总体而言,随着树龄的增大,4 个优良家系胸径之间的差异略微变小,树高之间的差异略微变大。
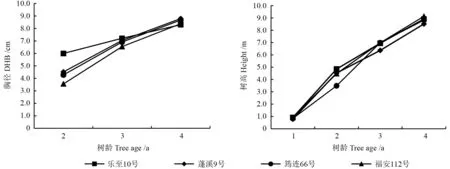
图1 优良家系胸径、树高的生长规律Fig.1 Growth rule of DBH and height of superior families
3 结论与讨论
香椿优树半同胞家系的生长除了与优树本身的遗传因子有关外,还与优树种源所在地的环境因子有关。香椿不同家系的胸径、树高和单株材积与纬度、经度和海拔呈不同程度的负相关性,与孙鸿有等[17]的研究结果一致,与年均气温、降水量和无霜期呈显著的正相关性,地理位置与香椿生长的相关性大小依次为纬度、海拔和经度,气候与香椿生长的相关性大小与年均气温和年均降水量相当,均大于无霜期。10 个香椿种源中总体上四川的种源好于广西和福建的,陕西的种源生长最差,与相关性分析结果一致。因此在进行良种选育时,应该在比试验区域所在位置纬度低、海拔低、经度小以及无霜期长、年均气温高、降水量大的种源中进行优树选择,构建半同胞家系,开展良种选育,更有利于选育出适合该区域的香椿良种。
44 个香椿优树半同胞家系及对照的平均胸径、树高和单株材积分别为7.14 cm、7.54 m 和0.017 8 m3,变幅分别在2.36~10.47 cm、3.12~11.50 m、0.001~0.048 m3之间,不同家系间均有极显著差异,与大部分香椿半同胞家系早期选择的研究结果一致[2-8],具有很好的早期选择基础。在44 个香椿优树半同胞家系及对照中,胸径生长以蓬溪9 号最大,其次是筠连66 号,平均胸径分别为8.79 cm 和8.67 cm,年均胸径生长量>2.12 cm,遗传增益分别为16.51%和15.25%;树高以福安112 号最大,其次是乐至10 号,平均树高分别为9.13 m 和8.91 m,年均树高生长量>2.22 m,遗传增益分别为14.69%和12.66%;单株材积最大的是福安112 号、乐至10 号、蓬溪9 号和筠连66号4 个家系,平均值为0.027 9 m3,平均遗传增益为33.84%。
变异系数的大小决定了林木生长的均匀程度。44 个香椿半同胞家系及对照中,有23 个家系胸径的变异系数小于15%,有28 家系树高的变异系数大于15%,单株材积的变异系数除了蓬溪15 号外均大于15%,变异系数总体比苗期大[10-11],表明香椿半同胞家系随着树龄的增长,家系内逐渐出现了一定的个体分化,但大部分家系的胸径和树高生长还是较为一致,有利于开展家系良种选育。
遗传力反映亲代将把性状遗传给子代的能力[18],本研究44 个香椿优树半同胞家系的胸径、树高和单株材积的遗传力分别为0.712、0.698 和0.604,胸径的遗传力略大于树高和单株材积,单株材积是胸径和树高累积效应的结果,以单株材积为主,胸径和树高为辅进行优良家系早期选择。为了提高选择效果,降低了入选率,提高了选择差[19],选择出福安112 号、乐至10 号、蓬溪9 号和筠连66 号4 个家系,入选率为8.89%。胸径、树高、单株材积的选择差分别为1.40 cm、1.30 m 和0.010 0 m3,遗传增益分别为13.95%、12.01%、33.84%。入选率很低,选择差较大,胸径、树高、单株材积的增益均较大,选择效果较好,经过半个以上轮伐期的观测,4 个家系中很有可能选育出适宜川中丘陵区发展的香椿良种。
在自然环境中选择优树即选择了好的变异,是良种选育的基础[20-21]。本研究利用优树的半同胞家系开展优良家系选择,取得了理想的选育结果,选育出4 个早期优良家系,有很大的生长潜力,但因只是4 a 的观测结果,且只是一个试验点的结果,下一步还需继续观测到半个轮伐期,且结合其他试验点的结果进行分析与比较,方可对其中的优良家系进行良种审定,为川中丘陵区的香椿造林提供良种。
