植物内生固氮菌及其固氮机理研究进展
2022-02-11王玉虎赵明敏郑红丽
王玉虎, 赵明敏, 郑红丽
内蒙古农业大学园艺与植物保护学院,呼和浩特010019
植物内生固氮菌(endophytic diazotroph)是指与宿主植物进行联合固氮并且定殖于植物体内的一类微生物[1]。它们不需要与植物产生结瘤,就可以侵入到宿主植物体内并定殖,可以直接或间接提供植物生长所需要的氮源,除了固氮作用外已经被证实的有产IAA、溶磷、产铁载体、拮抗病原菌等功能,可用于生物防治、促进植物生长等领域。植物内生固氮菌的研究始于20世纪60年代,之后在牧草、甘蔗、水稻等多种作物中均分离出了内生固氮菌。由于内生固氮菌定殖在植物体内,相比于根际和共生固氮菌位置优势更明显。因此,研究内生固氮菌与植物之间的关系对农业生产、可持续发展等具有重要意义。
1 植物内生固氮菌的发现
1961 年,Dôbereiner 等[2]在尝试研究甘蔗固氮系统时,首次从根际分离到固氮细菌——拜叶林克氏菌(Beijerinck),这也是研究者首次证实禾本科植物存在生物固氮潜力。Dôbereiner 等[3]在1974 年研究了热带牧草生物固氮系统,其由固氮螺菌(Azospirillum)和牧草形成,而且还发现固氮螺菌大多数聚集在牧草根表,但是有一部分菌株可以穿过根表皮细胞壁侵入到牧草根内部,这些侵入到植物体内的菌不与宿主植物形成特殊的共生组织,而是处于一种没有表型特征而且松散的共生状态,这一现象被称为联合共生固氮作用(associative symbiotic nitrogen fixation),这种固氮类型是介于根际自生固氮和结瘤固氮之间的过渡类型。Dôbereiner 团队还从甘蔗组织内部发现并分离出固氮菌,因其具有将乙醇氧化成乙酸的能力而且能够在酸性条件下固氮,所以被命名为醋酸固氮菌(Acetobacter diazotrophicus)。这些菌在甘蔗根和茎中大量存在,并且可以在含糖培养基上表现出最佳生长态势,这正是其在甘蔗汁中普遍存在的生长条件[4],之后又被重命名为固氮葡糖醋杆菌[5]。Helvécio 等[6]采用乙炔还原法测定了2 种热带牧草中的固氮酶活性,并用15N 标记证实了植物的固氮能力。1978 年Tarrand 等[7]命名了拜叶林克氏菌属(Azospirillum lipoferum)和巴西固氮螺菌属(Azospirillum brasilense)的新型菌株,这2种菌是由Dôbereiner 从马唐属和小麦中分离出来的。虽然越来越多的固氮菌被分离出来,但是对于固氮菌的划分一直并未明确。直到1997 年Joséi 等[8]研究了根际、内生、兼性和专性固氮菌,然后根据固氮菌菌体是否可以侵入植物组织内部并定殖,将固氮菌分为根际固氮菌(rhizosphere diazotrophs)和内生固氮菌(endophytic diazotrophs),之后相继发现甘蔗、水稻、玉米、牧草等具有重要经济价值的作物中均存在明显的生物固氮现象。
植物内生固氮菌能否顺利侵入植物体内并定殖是发生联合固氮作用的决定性因素[9]。植物内生固氮菌由于在植物组织内部,相比于根外占据着更加有利的地位,具有供应营养物质、与病原菌竞争营养和空间、分泌植物生长激素等优势,相较于根外更容易形成高效固氮体系。内生固氮菌与植物的独特关系和环境特点潜力,使其成为了一个具有重要意义的研究方向。
2 内生固氮菌与多种作物共生
内生固氮菌根据其特点不同,又可分为兼性内生固氮菌和专性内生固氮菌2 类。国外在甘蔗上的内生固氮菌相关研究报道较多。1961 年,Döbereiner[2]首先在甘蔗根际分离到大量的Beijerinck属细菌,并提出了甘蔗联合固氮作用(sugarcane associative nitrogen fixation)的概念。随后陆续报道在甘蔗的根、茎、叶及根际中分离到了多种固氮细菌。2011 年我国科学家在广西种植的不同甘蔗品种中分离出内生固氮菌[10]。黄杏等[11]通过采用3 种不同固氮菌的混合菌液接种甘蔗,发现甘蔗的根系活力、碳水化合物及蛋白质含量均得到明显提高。罗霆等[12]采用15N 同位素稀释法证明在接种固氮菌后,甘蔗的固氮量和固氮百分比均得到提高,同时,魏春燕等[13]发现甘蔗基因型对甘蔗与固氮菌间的联合互作有影响。最近,史国英等[14]从广西甘蔗茎基部组织分离筛选到1 株内生固氮菌株NN08200,其固氮促生效果明显,有望研发成为优良固氮微生物肥料生产菌种,并可进一步用于甘蔗联合固氮菌作用机理的相关研究。
除了甘蔗之外,国内的科学家在水稻和玉米上的内生固氮菌研究报道最多,如丘元盛等[15]在广东省水稻根分离出来的粪产碱菌(Alcaligenes faccalis)。研究者还从水稻上分离到内生阴沟肠杆菌(Enterobacter cloacae)[16]。华南农业大学谭志远团队[17]在野生稻内生固氮菌方面的研究比较深入,已取得了一系列研究成果。该团队在普通野生稻、澳洲野生稻、岑溪药用野生稻、藤县药用野生稻中均分离到多种内生固氮菌,并证明这些分离到的菌不仅在遗传方面具有多样性,而且还含有一些高固氮酶活性的菌株,可以达到42.52 μmol C2H4·mL-1·h-1。该团队将普通野生稻分离出的内生固氮菌进行回接实验,结果表明其能够显著促进水稻生长。野生稻不仅是我国的一种重要种质资源,也是宝贵的功能微生物资源库[18]。在野生稻内已发现的固氮菌目前有31 个属74 个种,其中兼性内生固氮菌占绝大部分,并且寄主范围广、分布地域全球化。野生稻内生固氮菌存在巨大发掘空间,现有研究大多集中于其促进生长、抗病害作用等方面。
自从1996 年美国科学家Palus 等[19]从甜玉米茎内分离到可固氮的肺炎克雷伯氏菌(Kelebesilla pneumomiae)以来,此后相继在玉米的根、茎、叶上分离到内生固氮菌[20]。其中,同一玉米样品根内生固氮菌的多样性最高,其次是叶,茎中最低[21]。一些玉米根、茎、叶上分离到的固氮菌株,如L1A1、L1A4、L1B9 和 L2A2 是多功能固氮菌,有研究者对这4 株菌的固氮、溶磷、拮抗、促生等一系列指标进行评价后,将这4菌株接种到小麦上,发现小麦的农艺性状有明显改善,植株更加健壮,幼苗的长势也更好,并且在一定程度上可以提高小麦的抗干旱能力[22]。覃姚红等[23]研究表明,玉米种子内生固氮菌对小麦生长及抗旱能力也有促进作用。此外,科学家们在小麦、禾本科牧草、蔬菜、果树、林木上也分离鉴定出了不同类型的内生固氮菌,并研究了固氮活性及其菌株特性[24-28],这引起了人们对植物内生生物固氮作用的广泛关注。
近年来,内生固氮菌在薯类作物上的报道较少。Sessitsch 等[29]用 TRFLP 和 DGGE 技术相结合分析了不同马铃薯品种的地上茎、根及块茎内生细菌的多样性。Burkholderia vietnamiensisstrain RS1 是从甘薯根部分离获得的内生固氮菌,其基因组大小为6 542 727 bp[30]。基因组中含有固氮基因 nif 基因簇(46.4 kb),由61 个开放阅读区组成框架,包括nifQXNEKDHOTSZZBAUVPW 和与nif相关的基因(modACE、fdxBN 和fixAB)。此外,基因组中鉴定出几个与植物生长促进有关的基因,如产铁载体基因和1-氨基环丙烷-1-羧酸的降解基因[30]。张磊等[31]从内蒙古武川县的马铃薯根样品中采用固氮基因nifH扩增方法获得8 株内生固氮菌,其中3 株为短芽孢杆菌属(Brevibacterium),3 株为芽孢杆菌属(Bacillus),2 株为泛菌属(Pantoea)。目前在不同植物上分离得到的内生固氮菌详见表1。

表1 不同植物上分离到的内生固氮菌Table 1 Endophytic nitrogen-fixing bacteria isolated from different plants
3 植物内生固氮菌的侵入途径与定殖
根据已有研究,植物体内的内生菌来源主要有2 种方式,第1 种是通过种子和块根、块茎等无性繁殖材料。这些菌侵入到种子内并且能够系统地定殖,从种子萌发一直到植物开花结实始终存在于植物体内,伴随着植物整个生长周期,且植物与内生菌形成了互利共生的关系,如赛鲁普蒂卡草螺菌是前人从水稻的种子内分离得到的[32-33]。第2 种方式是通过根围和叶围侵入,以根围侵入的方式为主。Hallmann 等[32]在分离的棉花内生菌中发现有82%也存在于根际中并且能够分离得到。Boddey[34]研究认为,甘蔗固氮菌是通过茎切口进入并且繁殖内生寄生菌。在玉米和其他禾本科牧草的内生固氮菌也多数来自根围。Sessitsch等[35]研究了马铃薯根、地上茎、块茎3 个不同部位的内生菌,内生菌多样性方面健康马铃薯植株明显高于受胁迫植株,并证明内生菌相当一部分起源于宿主植物根际。
种子的内生固氮菌主要通过种皮裂开的缝隙、种脐或者维管系统进入种子内部,并借助种子调运进行远距离传播[36]。来源于根围或叶围的内生菌侵入寄主植物主要通过植物自然孔口(气孔、表皮裂隙、皮孔等)、伤口(机械损伤、病害损伤、自然灾害损伤等)和根毛。植物根生长的初期因幼根组织较弱,凯氏带尚未形成,成为固氮菌侵入的最佳时期。根皮层细胞间、叶肉、叶薄壁组织和维管组织中的木质部是内生固氮菌侵入后定殖最多的部位,根(根毛)表皮细胞同样分布着较多的内生固氮菌[37]。内生固氮菌还可以利用宿主植物体内的分泌物或者其他产物,如重氮营养醋杆菌(Acetobacter diazotrophicus)能够利用甘蔗细胞间溶液生存并定殖于甘蔗体内,这是因为细胞间溶液的蔗糖浓度和pH比较适合,并且成分接近培养重氮营养醋酸杆菌的培养基。固氮醋酸杆菌的侵入是由根尖和侧根的发生部位开始,它们从根部侵入后进入上皮细胞,从茎部的木质部导管向上传输到幼嫩组织和叶肉组织,且能侵入甘蔗根毛形成侵染线样结构[38](表2)。也有研究表明,内生固氮菌表面的纤维素酶和果胶酶是可以改变寄主植物细胞膜通透性的,从而引起寄主植物细胞K+外流,细胞间隙溶液的 pH 从 5.5 升高到 7.0~7.5,使寄主植物细胞中蔗糖、氨基酸等营养物质流入细胞间隙,从而为内生固氮菌的定殖创造条件[39]。

表2 几种代表性内生固氮菌侵入植物的途径Table 2 Pathway of several representative endophytic nitrogen-fixing bacteria invading plants
据推测,内生固氮菌定殖前会与植物进行一系列的相互作用,这些作用与根瘤菌和植物特异性结合比较相似。根瘤菌表面产生的多糖可以与豆科植物根系分泌的豆凝素特异性识别,然后结合并吸附在根毛上。某些细菌的表面结构能够在内生固氮菌侵入过程中起到接触和识别作用,特别是革兰氏阴性细菌外膜上LPS的O-抗原特异侧链在细菌的特异性识别和接触过程中发挥重要作用[40]。此外,也有研究者推测内生固氮菌的定殖部位与高效固氮机制是由于内生固氮能够诱导呼吸系统变化,使环境中的氧分压降低,呼吸作用变得更加活跃并且能够高效固氮。推测是在这种“超诱导”的状态下,固氮酶的铁蛋白在细胞内膜上聚集,与形成的高效固氮功能膜“diazosomes”有关[41]。
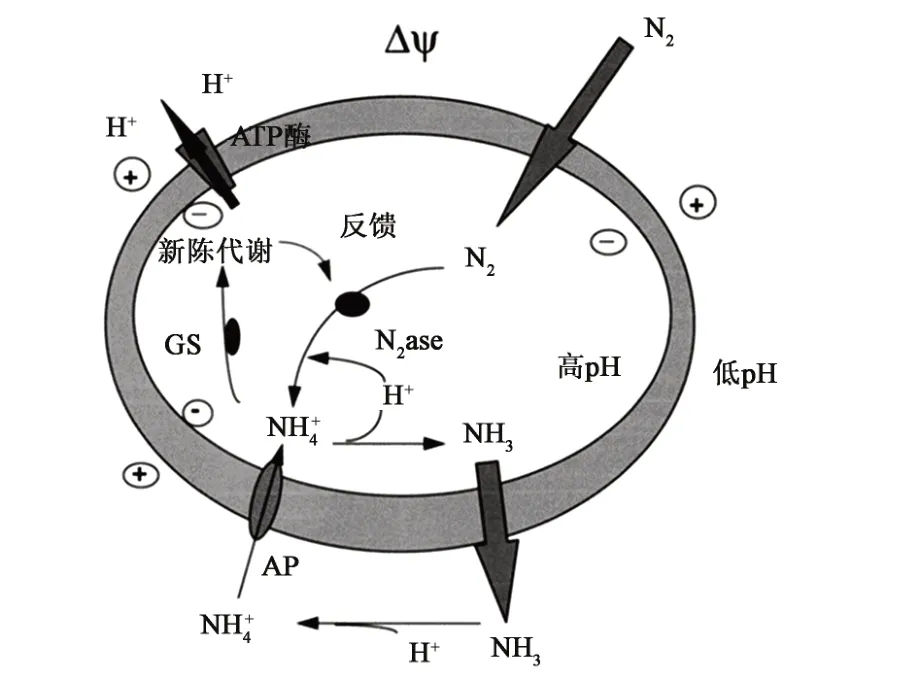
图1 氮从重氮营养体向寄主植物的转移模型[31]Fig.1 Model for nitrogen transfer from diazotroph to host plant[31]
4 植物内生固氮菌的固氮机理
固氮菌之所以能够固氮是因为其体内有一种具有特殊功能的蛋白质——固氮酶,而固氮功能与固氮酶结构、调控基因、活性密切相关。
4.1 固氮酶结构及调控基因
固氮酶由固氮基因nif编码,nif一般存在于细菌的质粒或者染色体上。在固氮菌的基因组中,固氮相关的nif(vnf/anf)基因都是作为一个或几个基因簇存在的,称为固氮基因簇(nitrogen-fixationgene cluster),是生物固氮遗传基础的核心[42]。人们可利用已知固氮菌nif基因簇中的共有基因簇特征序列,进行筛选发现新的固氮菌。目前已知的固氮酶包括钼铁(MoFe)固氮酶、钒铁(VFe)固氮酶和铁铁(FeFe)固氮酶3 种类型。虽然这3 种固氮酶中所含金属离子不同,但是这些固氮酶亚型在结构催化机制和系统进化上是相关的[43]。固氮酶由两部分组成(图2):nifH编码的铁蛋白(又称为固氮还原酶)和nifDK编码的钼铁蛋白(又称为固氮酶),nifD编码钼铁蛋白的 α 亚基,nifK编码钼铁蛋白的β 亚基[44]。当这2 种蛋白形成一种固氮酶复合物后才具有催化氮还原成氨的能力,单独任何一个都不具有固氮活性。铁蛋白负责转移电子给钼铁蛋白,其是1 个由Fe4-S4簇桥连的同源二聚体[45],包含在2个亚基之间的界面上,通过每个亚基中的2 个半胱氨酸残基与之结合。钼铁蛋白负责铁蛋白与铁钼辅因子之间的电子转移,它是 1 个 α2β2异四聚体,含有 1 个 P 簇(Fe8-S7)。铁钼辅因子是1 个(7Fe-Mo-9S-homocitrate-X)簇,作为底物结合和还原的位点偶联Mg-ATP 水解释放的自由能,铁蛋白与钼铁蛋白共同催化N2还原为NH3的反应。如图2所示,当铁蛋白与钼铁蛋白互作时,Fe 蛋白中的Fe4S4团簇与MoFe 蛋白中的P团簇接近,然后电子被转移到铁钼辅因子,N2分子被还原成NH3分子。整个反应可以写为式(1)。

图2 固氮酶复合物(PDB ID 1N2C)三维结构[44]Fig 2 Three dimensional structure of the nitrogenase complex(PDB ID 1N2C)[44]
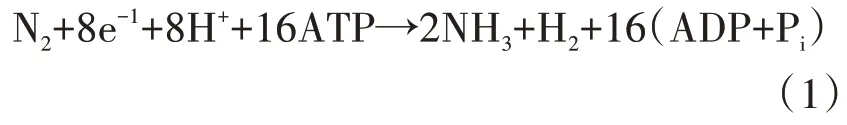
比较所有测序的固氮菌基因簇表明,至少有6 个保守基因:nifH、nifD、nifK、nifE、nifN和nifB,并且它们都是固氮所必需的[46],其中,6 个共有基因中至少有3 个位于1 个连续的区域内,通常是nifHDK成簇存在;也有nifDK与nifEN相邻的情况;也有的6 个共有基因位于基因组内的一个单独区域内,但是这种情况较少见。
4.2 固氮酶活性调节机制
在某些固氮光合细菌和固氮螺菌中,固氮活性在代谢水平上受到调节,这种调节最早在深红红螺菌(Rhodospirillum rubrum)中发现,固氮酶活性调节表现为一种关闭效应(switch-off)。当光关闭或者将效应物(铵离子或谷氨酰胺)添加到固氮培养物中时,固氮酶活性丧失。当光被接通或添加的效应物被代谢,固氮酶活性恢复。在巴西固氮螺菌中能量的关闭效应是由氧气的去除引起的。在分子水平上,固氮酶活性丧失是由于铁蛋白亚基之一精氨酸的可逆ADP 核糖基化(图3),在深红红螺菌中为Arg101。Arg101 位于Fe 蛋白和MoFe 蛋白的相互作用区域内,因此,修饰在空间上阻碍了两种蛋白的相互作用,从而无法发生电子转移。催化可逆修饰的两种酶是DraG(dinitrogenase reductase activating glycohydrolase)和 DraT(dinitrogenase reductase ADP-ribosyl transferase)。目前人们对DraG 和DraT 在分子水平上的了解十分有限,关于DraT 的调控知之甚少,但可以确定PII蛋白在响应细胞的氮状态时参与了这种调控。PII蛋白构成1 个三聚体蛋白家族,在原核细胞中调节与氮代谢相关或部分氮代谢的过程。DraG活性调节已确定与深红红螺菌中的色素细胞(质)膜相关[44]。

图3 DraT和DraG对Fe蛋白的可逆ADP核糖基化作用[46]Fig 3 Reversible ADP-ribosylation of the Fe protein by DraT and DraG[46]
目前,在甘蔗固氮葡糖醋杆菌(Gluconacetobacter diazotrophicus)上的研究已经十分成熟,在固氮机理、酶和基因等方面的研究比较深入。先前的研究发现固氮葡糖醋杆菌上的nifA基因位于染色体上,并且对O2和NH4+敏感,该基因的排列和功能与其他固氮菌相似,固氮酶的最适催化pH和最大活性pH 为7.0,并且固定同化的氮是谷氨酰胺(Gln)合成酶催化的。有研究者从固氮酶上分离到结构基因GlnA,并且利用插入突变技术确定了表达调控蛋白的GlnB同源基因(GlnB,Glnk1和Glnk2)的存在,这些调控蛋白能够感知铵离子浓度,进而调控固氮酶的表达和活性[47]。巴西杆菌的固氮调控模型也已经被提出,其中PⅡ蛋白直接或间接地控制了nif基因转录激活因子NifA的活性,尿苷化形式的PⅡ蛋白是活性nifA 蛋白所必需的,而且尿苷化形式的PⅡ蛋白是双功能尿苷转移酶/尿苷去除酶GlnD 催化的。此外,有研究者利用GlnD突变株证实GlnD 蛋白参与解除NifL 对NifA活性的抑制[48]。
4.3 固氮酶氧保护机制
固氮酶还有一种防止氧伤害的保护机制[49]。固氮作用必须在无氧条件下进行,这是因为固氮酶对氧极其敏感。钼铁固氮酶是一种氧敏感酶,氧气会影响固氮酶活性,但是大多数固氮菌必须在有氧条件下才能生活,因此固氮菌自身产生了一种保护机制来防止氧损伤固氮酶。目前已经明确的固氮酶保护机制,有呼吸保护、构象保护、“双保险”或调节机制3种。
4.3.1 呼吸保护 固氮菌属的许多细菌通过提高自身的呼吸强度迅速耗尽固氮部位周围的氧,以使固氮酶处于无氧的环境中从而免受氧的伤害。在呼吸保护中,呼吸链的组成部分涉及氰化物敏感的细胞色素带抗氰化物细胞色素巴酚氧化酶,在高氧条件下固氮酶保护起主要作用。
4.3.2 构象保护 大多数固氮菌体内均含有一种特殊功能的蛋白质——Fe-SⅡ蛋白,其能够起到构象保护的作用,在氧分压增高时,该蛋白与固氮酶结合,此时,固氮作用随着固氮酶构象的改变而丧失;固氮酶周围的氧浓度降低后,该蛋白便从酶分子上解离,固氮酶恢复原有的构象和固氮能力。Gluconacetobacter diazotrophicus还可以利用菌落黏液的路径长度来维持最佳的O2浓度,且不抑制固氮酶活性和细胞代谢。呼吸保护和构象保护均涉及Fe-SⅡ蛋白协同作用。
4.3.3 “双保险”式调节机制 通过加强呼吸作用与改变构象两者相互协调,组成一个“双保险”式调节机制,以保证固氮酶的活性。
此外,要实现生物固氮还必须具备以下几个基本条件:还原型电子供体、ATP 和Mg2+离子、来源于水的质子、严格的无氧环境。实际上固氮酶的机理非常复杂,至今仍有很多问题尚不明确。
5 植物内生固氮菌对植物的促生作用及其机制
内生固氮菌侵入植物体内后定殖在木质部导管并发挥固氮能力。木质部导管可以提供细菌生长所需要的能源、满足低氧分压的环境,还可以提供交换代谢产物的场所,避免与土壤微生物之间的竞争和土壤化合态氮对固氮酶活性的抑制作用,固定的氮无需经过其他方式即可以直接被植物吸收利用。目前已发现的植物内生固氮菌多通过以下几方面发挥促生作用。
5.1 为植物提供氮源
内生固氮菌都含有固氮酶,能够大量定殖在植物组织内部,而且无需形成特殊的共生结构或组织就能发挥固氮作用,这与固氮菌能够形成被膜或基质包围的内生特征有关[50]。它们可以定殖在宿主植物体内,能够侵入宿主植物的大部分营养器官内并且发挥固氮作用,为植物提供可利用的氮源,有些内生固氮菌固定的氮素还可以直接被植物吸收同化。由目前的研究表明,这一系列作用不会引起寄主植物产生任何不良反应[51],有些可以将大气中固定的氮素一部分提供给宿主吸收利用,其余部分被细菌自身转化利用,当细菌死亡后释放出的有机氮也被植物吸收利用。
5.2 产生植物激素类物质
内生固氮菌能分泌的刺激植物生长的植物激素类物质有吲哚乙酸、赤霉素、细胞分裂素等[52]。分离自墨西哥的重氮营养醋杆菌(Acetobacteria)具有产生长素(IAA)的能力[53]。林敏[54]分析水稻根系成分时,发现粪产碱菌(A.faecalis)存在于水稻根际并且能产生赤霉素、生长素及玉米素,可以促进水稻根系生长发育,同时刺激水稻根的分泌作用。田宏等[55]从草坪草根际得到分离物,对分离物成分进行分析并发现大部分都能够产IAA,但产IAA 的能力差异较大,浓度在 3.18~22.3 mg·L-1之间。植物根内生细菌可以通过吲哚-3-丙酮酸(IPyA)、吲哚-3-乙酰胺(IAM)、吲哚-3-乙腈(IAN)和色胺(TAM)等途径来合成IAA[56],从而促进宿主植物的生长或者调节植物生理代谢活动。也有内生固氮菌通过产生铁载体和1-氨基环丙烷-1-羧酸(1-aminocyclopropane-1-carboxylate,ACC)脱氨酶等其他物质来促进植物生长[57]。阳洁等[58]从野生稻中分离到多种内生固氮菌,接种后能显著促进水稻生长。尹坤等[59]从野生稻体内分离到1 株变栖克雷伯氏菌(K.variicola),这株菌不仅具有较强的分泌生长素和铁载体的能力,还能够有效促进水稻的萌发和生长。
5.3 促进内根际的生理变化
研究发现,植物内生固氮菌具有促进内根际的生理变化作用。主要表现在促进根际多元酚的分泌、促进植物根际ATP 含量增加、促使根部氧化酶活性降低,提高还原酶活性。以固氮葡糖醋杆菌为例,其具有糖酵解、戊糖磷酸途径和一种独特的细胞外醛糖氧化途径,利用含吡罗-喹啉-醌(PQQ)的葡萄糖脱氢酶将胞外葡萄糖氧化为葡萄糖酸酯[60]。在此过程中产生的额外还原当量主要通过产生ATP 的电子传递链传递,ATP 同时用于固定N2过程,这一途径在高能量需求和碳限制条件下均可运行,同时还具有溶磷等功能以改善根际内生理条件。
5.4 生物防治作用
内生固氮菌对植物病害有显著的抑制作用。Braun 等[61]在研究成团肠杆菌时发现,接种成团肠杆菌的小麦可以有效防治丁香假单胞菌引起的小麦粒疫病,并且防治率可以达到45%~74%。杨海莲等[62]也发现采用阴沟肠杆菌接种水稻可以对水稻纹枯病、稻瘟病和白枯病起到一定防治效果。目前,对固氮菌生防作用的机理有2种推测,一种是固氮菌通过在与致病菌的竞争过程中占据有利生态地位达到抑制致病菌的作用;另一种认为可能是内生固氮菌产生了某种特异性抑菌物质,从而发挥抗病功能[63]。
6 展望
植物内生固氮菌作为植物微生态系统的天然成分,由于长期的共同进化,植物内生固氮菌与宿主建立了良好的互利共生关系。内生固氮菌寄生于宿主植物组织内,并不引起直观的表型变化,使该领域的研究具有一定的难度;同时,由于内生固氮菌的生境特殊,并受多种因素的影响,限制了内生固氮接种剂在农业生产中的应用。近年来,随着一些新研究方法的应用,包括化学分析、遗传工程、分子生物学、免疫学等,推动了固氮菌领域的研究深度,使人们进一步了解内生固氮菌与植物之间的相互关系,并促进了内生固氮菌合理运用。但目前,仍有很多问题尚未解决,如内生固氮菌的来源、侵入的分子机制、作用特点及人工接种其他植物是否会造成影响,此外,针对固氮机理方面的研究也并不深入,生防机制方面也只是推测并未明确。因此,明确植物和内生固氮菌的互作关系,以及内生固氮菌与植物根形成固氮体系的机理等,将更有利于人们了解内生固氮菌并充分利用,对农业可持续发展具有重要意义。
