瑞安市腹泻患者沙门菌耐药性和分子分型研究
2022-02-04李娜周邦瑶苏海珍胡玉琴叶环环刘敏芝李毅
李娜 周邦瑶 苏海珍 胡玉琴 叶环环 刘敏芝 李毅
沙门菌可引起人类肠道传染病和伤寒、副伤寒病症,亦可使家禽、畜类致病,是一种人畜共患病的病原体。在中国乃至世界各地,非伤寒沙门菌是引起人类食源性疾病、造成重大经济损失的重要元凶[1-2]。Majowicz等[3]估计,全球每年约有9 380万例沙门菌感染患者,病死15.5万例,其中8 030万病例来源于食源性感染。沙门菌血清型众多,全球范围内已经鉴定出的沙门菌血清型多达2 500个[4]。不同血清型沙门菌遗传表型不同,这为确定感染源及临床控制感染造成一定难度。本研究收集瑞安市人民医院2016—2019年从腹泻患者中分离到的沙门菌,通过血清型鉴定和药物敏感性分析,了解瑞安市沙门菌的血清型分布和耐药性的总体概况,为临床科学用药提供实验室参考数据;并利用脉冲场凝胶电泳(pulsed field gel electrophoresis,PFGE)技术分析沙门菌分子分型,为细菌溯源提供理论依据。
1 材料和方法
1.1 菌株来源 2016—2019年从瑞安市人民医院腹泻患者中分离到沙门菌共134株,其中2株分离自血液,其余132株分离自粪便。
1.2 主要试剂 沙门菌血清诱导琼脂(批号:295652149)和沙门菌血清套装(规格:1 ml,60支/盒,批号:K60V-50)购自丹麦SSI公司。药敏纸片及MH琼脂(批号:2487022)购自英国OXOID公司。14种药敏纸片分别为氨苄西林(批号:2858417),阿莫西林/克拉维酸2∶1(批号:2491412),头孢他啶(批号:2912216),头孢噻肟(批号:2518697),头孢吡肟(批号:2836423),亚胺培南(批号:2897217),氨曲南(批号:2518702),萘啶酸(批号:2492578),环丙沙星(批号:2539801),磺胺异恶唑(批号:2984526),甲氧苄啶(批号:2538102),复方新诺明(批号:2518734),四环素(批号:2525329),氯霉素(批号:2506047)。Xba I核酸内切酶(批号:AGZ0057A)购自宝生物工程(大连)有限公司,蛋白酶K(批号:812KA0961)购自上海生工生物工程有限公司。使用法国梅里埃VITEK-2全自动生化鉴定系统,美国Bio-Rad公司CHEF-Mapper®XA脉冲场凝胶电泳仪及Gel XR凝胶成像仪。
1.3 方法
1.3.1 菌株鉴定及血清学分型 按照国家标准《食品微生物学检验沙门氏菌检验》(GB 4789.4-2016)对菌株进行鉴定与血清凝集试验。依据《White-Kauffmann-Leminor抗原表(第9版)》和《沙门菌属血清型诊断》确定血清型[4]。
1.3.2 药物敏感性检测 采用K-B纸片法,根据美国临床实验室标准化协会(clinical and laboratory standards institute,CLSI)药敏试验标准 M-100(2019)[5]判定药敏结果。以氨苄西林、阿莫西林/克拉维酸2:1、头孢他啶、头孢噻肟、头孢吡肟、亚胺培南、氨曲南、萘啶酸、环丙沙星、磺胺异恶唑、甲氧苄啶、复方新诺明、四环素、氯霉素等14种药物作为实验抗菌药物。以大肠埃希菌ATCC25922为质控菌株。
1.3.3 PFGE分子分型 随机选取部分菌株,参照国家食源性致病菌病原细菌分子分型监测网络(PulseNet)中沙门菌PFGE分型的标准操作方法进行。将新鲜菌株悬浮于盛有1 ml细胞悬液的Falcon 2054管,调整细菌密度,使吸光度值为4.0~4.2。取400 μl上述细菌菌悬液于1.5 ml离心管中,加入20 μl蛋白酶K(20 g/L),混匀,加入400 μl SeaKem Gold琼脂糖混匀,将上述混合物加入加样孔内,待胶块凝固后,加入25 μl蛋白酶K(20 g/L)的裂解液裂解细菌2 h。加入45 U XbaⅠ酶,37℃酶切3 h,以布伦登卢普沙门菌菌株H9812作为分子质量标准。脉冲场凝胶电泳条件:6.0 V/cm,2.16~63.8 S,19 h,电泳结束后,用Gel Red染色并用纯水脱色。采用凝胶成像仪获取图像,并转换成TIFF图像格式。用BioNumerics 7.5软件对PFGE结果进行分析,用非加权配对算术平均法(unweighted pair group average method,UPGMA)进行聚类,构建聚类树。
1.4 统计学处理 使用SPSS19.0统计软件。计数资料采用χ2检验或Fisher确切概率法,P<0.05为差异有统计学意义。
2 结果
2.1 沙门菌菌株来源的人群分布 134株菌株来源人群中,最小年龄<1岁,最大年龄为87岁。5岁以下婴幼儿25人,占18.7%,5~12岁6人,13~20岁7人,21~40岁36人,41~60岁43人,>60岁17人。男性83人,占61.9%,女性51人。其中0~2岁的婴幼儿达21人。
2.2 沙门菌血清型分布 134株沙门菌分别分布在B群、C1群、C2群、C3群、D1群、E1群和E4群,共24个血清型,占比最大的是鼠伤寒沙门菌,有52株,占38.8%,肠炎沙门菌31株,占23.1%,伦敦沙门菌13株,占9.7%,见表1。来自0~12岁儿童的菌株有31株,其中鼠伤寒沙门菌21株,占67.7%,肠炎沙门菌2株,占6.5%;来自21~60岁组的菌株有79株,其中鼠伤寒沙门菌19株,占24.1%,肠炎沙门菌25株,占31.6%。随着年龄增高,鼠伤寒沙门菌的占比呈降低趋势,>60岁组中,鼠伤寒沙门菌占比再次升高,见图1。≤12岁组和>60岁组中,鼠伤寒沙门菌占比远高于肠炎沙门菌;在21~60岁中青年组,肠炎沙门菌占比稍高于鼠伤寒沙门菌。≤12岁组和21~60岁组的血清型分布比较差异有统计学意义(P<0.05),见表2。

表1 134株沙门菌血清型分布情况

图1 各年龄组血清型占比

表2 134株沙门菌血清型分布情况
2.3 沙门菌药敏试验 134株沙门菌对氨苄西林的耐药率最高,达73.1%;其次为磺胺异恶唑、四环素和萘啶酸,耐药率分别为62.7%、56.0%和53.0%。复方新诺明的耐药率为32.8%。对环丙沙星耐药的菌株有13株,占9.7%,中介菌株有84株,占62.7%,敏感菌株37株,仅占27.6%。沙门菌对阿莫西林/克拉维酸2∶1、头孢他啶、亚胺培南、氨曲南具有较高的敏感性,耐药率均<10.0%。值得注意的是,2016—2019年,沙门菌对环丙沙星的耐药呈逐年递增趋势,如2016年耐药率为0,2017年为5.6%,2018年为11.5%,至2019年上升为15.8%。多重耐药菌93株,占69.4%,见表3。

表3 沙门菌对14种抗菌药物的耐药率(%)
比较鼠伤寒沙门菌和肠炎沙门菌的耐药性可知,肠炎沙门菌对萘啶酸耐药率为100%,鼠伤寒沙门菌对萘啶酸耐药率为48.1%,差异有统计学意义(P<0.05)。复方新诺明、氯霉素、甲氧苄啶、四环素的耐药率,鼠伤寒沙门菌高于肠炎沙门菌,差异有统计学意义(P<0.05);尤其是对四环素的耐药率,鼠伤寒沙门菌高达80.8%,而肠炎沙门菌为9.7%。其他9种药,两种菌耐药率差异无统计学意义(P>0.05)。52株鼠伤沙门菌中,多重耐药菌株有44株,占84.6%,大多为四环素和氨苄西林耐药。31株肠炎沙门菌中,多重耐药菌株有23株,占74.2%,耐药谱以萘啶酸、氨苄西林、磺胺异恶唑为主,见表4。
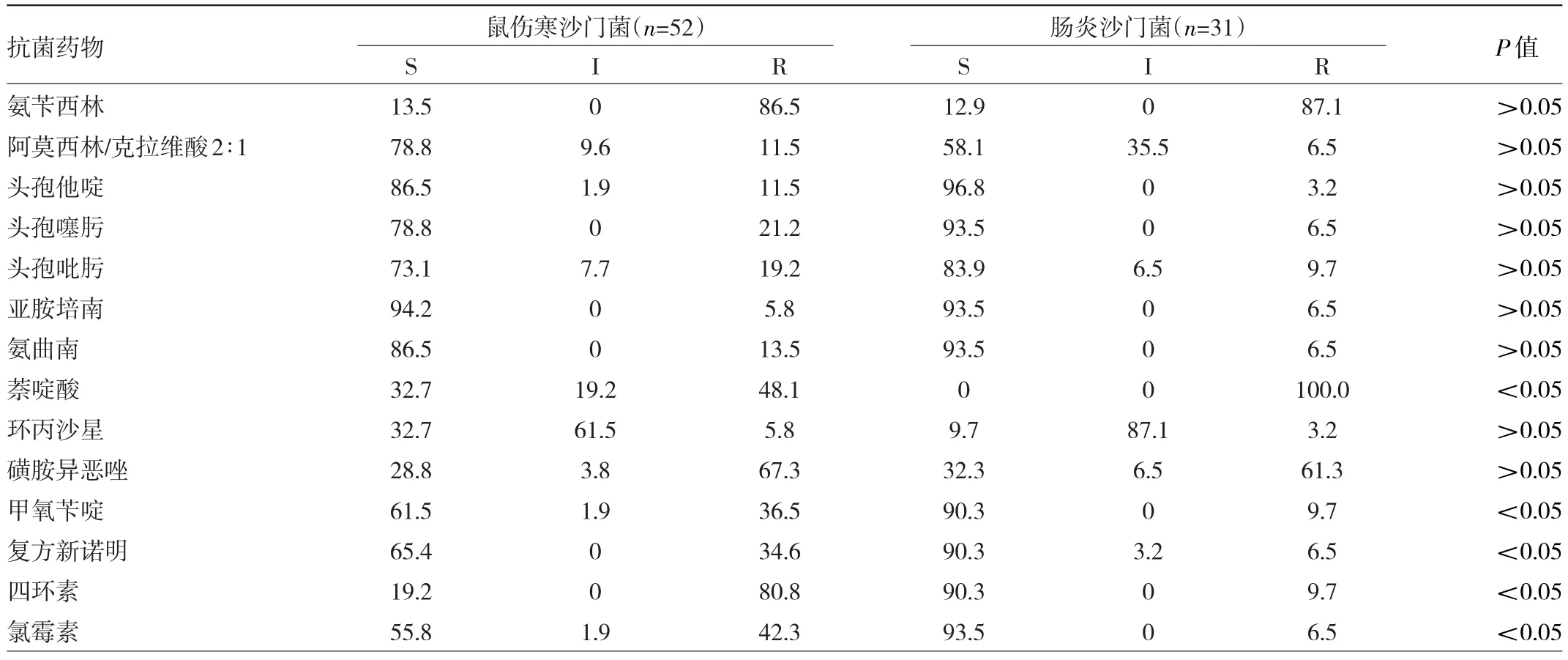
表4 鼠伤寒沙门菌和肠炎沙门菌耐药率比较(%)
2.4 PFGE分型结果 随机选取的40株沙门菌中鼠伤寒沙门菌18株,肠炎沙门菌14株,伦敦沙门菌3株,阿西纳沙门菌、山夫登堡沙门菌、利物浦沙门菌、汤卜逊沙门菌、罗森沙门菌各1株。UPGMA聚类显示大致分为鼠伤寒沙门菌、肠炎沙门菌、伦敦沙门菌为代表的3个支系,共29种带型,相似度为63%~100%。其中鼠伤寒沙门菌共16种带型,肠炎沙门菌共5种带型,除1株编号为SM167的肠炎沙门菌的带型与鼠伤寒沙门菌那支相近外,其余13株肠炎沙门菌带型相似度≥92.7%。3株伦敦沙门菌有3种带型,见图2。就菌株耐药性而言,编号为SM167的肠炎沙门菌对8种抗生素耐药,其余13株肠炎沙门菌对1~4种抗生素耐药。聚类在鼠伤寒支系中的阿西纳沙门菌、山夫登堡沙门菌和聚类在肠炎支系的利物浦沙门菌,对所有药物均敏感。编号SM43和SM93,SM44和SM47,具有相同的PFGE带型,却表现出不同的耐药谱。
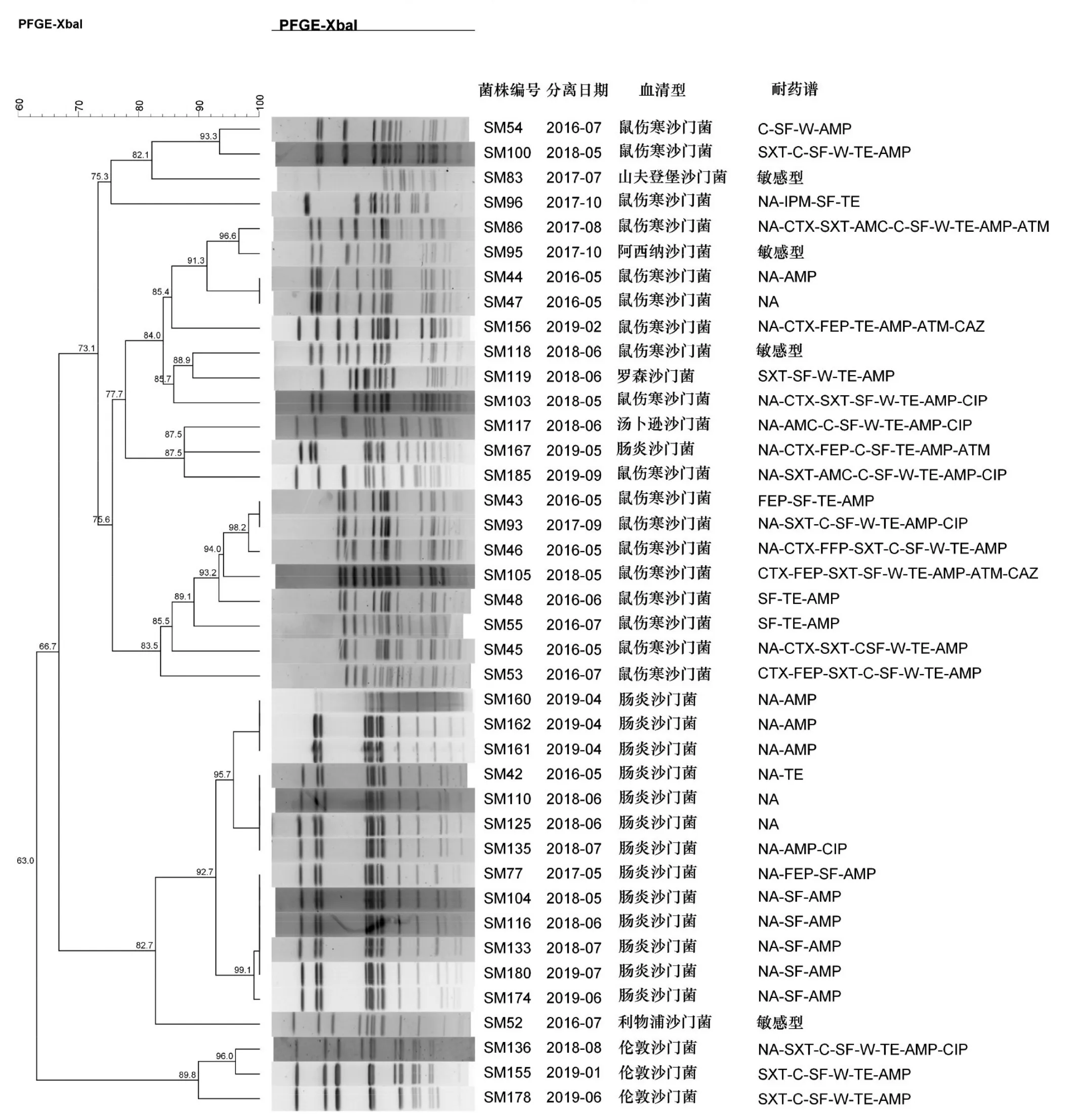
图2 40株沙门菌PFGE聚类分析图谱
3 讨论
近几年研究显示,沙门菌是中国儿童细菌性腹泻的主要病原菌[6-7]。2020年全国第七次人口普查结果显示,瑞安市0~14岁人口占14.52%,15~59岁占69.23%,>60岁占16.24%。本研究中23.1%的菌株来源人群年龄分布为0~12岁(31人),高于全部人群分布的构成比;其中0~2岁婴幼儿有21人,占15.7%。可见婴幼儿和低龄儿童是沙门菌的易感人群。相比健康人群摄入大量的非伤寒沙门菌才能引起疾病,一般幼儿和老年人以及慢性疾病患者更易感染,且症状严重[8]。
本研究分离的沙门菌血清型以鼠伤寒和肠炎沙门菌为主,与上海市和宝鸡市的一致[9-10],而向蓉等[11]和谭南等[12]发现中山市以鼠伤寒沙门菌和斯坦利沙门菌为主,安徽省马鞍山市以阿贡纳沙门菌和肠炎沙门菌为主[13],均与本研究不同。不同年龄组,沙门菌的血清型构成不同。随着年龄升高,鼠伤寒沙门菌的占比,呈现降低的趋势;但在>60岁组别中,占比又有所回升。本地区12岁以下儿童感染的优势血清型菌种为鼠伤寒沙门菌,占比高达67.7%,血清型比较单一;21~60岁成年人的感染菌株主要为肠炎沙门菌和鼠伤寒沙门菌,其他各血清型共占44.3%,血清型丰富。沙门菌感染是食源性疾病,当人群摄入被沙门菌污染的食品或接触带菌动物及其环境,就会引发疾病[14-15]。成年人饮食习惯多样、饮食结构丰富,与动物接触机会更多,可能是21~60岁成人组沙门菌血清型丰富的一个重要原因。
本研究发现,鼠伤寒沙门菌对复方新诺明、氯霉素、甲氧苄啶、四环素的耐药率显著高于肠炎沙门菌。而对于萘啶酸,31株肠炎沙门菌表现为全部耐药,显著高于鼠伤寒沙门菌(48.1%)。提示在研究沙门菌耐药性时,应注意肠炎沙门菌和鼠伤寒沙门的菌株数量占比,两者构成比不同会影响总体耐药率。一项针对2005—2014年沙门菌耐药性监测的研究显示,鼠伤寒沙门菌的耐药率普遍高于其他血清型[16];杨翊等[17]研究也显示,鼠伤寒沙门菌多重耐药率高于其他血清型,与本研究结果相似。高耐药株容易在同属同型菌株之间通过物流、旅游等人口流动发生传播扩散[18],提示需加强食品流通环节的监测,控制潜在的食源性爆发。
美国临床实验室标准化协会(CLSI)指出分离自粪便标本的沙门菌,药敏试验只需常规试验氨苄西林、一种氟喹诺酮类和甲氧苄啶/磺胺甲恶唑并报告[5]。临床上使用氨苄西林和喹诺酮类治疗非伤寒沙门菌感染,可能导致治疗失败。本地区的沙门菌对阿莫西林/克拉维酸2∶1、亚胺培南、氨曲南、头孢他啶仍具有较高敏感度。CLSI-2019公布的药敏试验标准(M-100)指出,除沙门菌属以外的肠杆菌科细菌对于环丙沙星中介的抑菌圈直径为22~25 mm,而沙门菌的抑菌圈直径为21~30 mm[5]。根据这一标准,本研究中多数沙门菌表现对环丙沙星中介。但值得关注的是,2016—2019年,沙门菌对环丙沙星的耐药率表现逐年上升趋势。
作为一种成熟且应用极广的分子分型技术,PFGE通过比较细菌分离株的指纹图谱,判定菌种间的遗传亲缘关系,被应用到流行病学的溯源研究中。PFGE结果显示,本地区2016—2019年的鼠伤寒沙门菌在分子水平存在一定程度的分化,而肠炎沙门菌除了1株特殊的分离株之外,其余菌株大多成簇聚类,相似度超过92.7。鼠伤寒沙门菌PFGE条带相似度很高的菌株,耐药谱却存在一定的差异。这可能是因为菌株耐药基因发生突变的位点并不在Xba I酶切位点上,无法由分型中表现出耐药性[19]。相比之下,肠炎沙门菌的耐药谱与PFGE条带重叠性较高,仅出现耐萘啶酸和氨苄西林的菌株聚类,耐萘啶酸、氨苄西林和磺胺异恶唑的菌株聚类。随着时间的演变,本地区沙门菌的耐药情况仍会继续变化。将全基因组测序方法应用于流行病学溯源调查以及临床用药指导。
志谢 感谢上海市疾病预防控制中心许学斌老师在血清凝集实验和论文写作等方面给予的指导
