长期高温与增施CO2对番茄叶片光合作用及淀粉含量的影响
2022-01-28范玉洁姜慧敏温祥珍白龙强李亚灵
范玉洁,姜慧敏,温祥珍,白龙强,李亚灵
(山西农业大学园艺学院,山西 太谷 030801)
北方日光温室夏秋栽培常因温度较高,导致作物产生高温障碍、光合速率下降、植株衰老加速、产量降低、品质变劣[1]。CO2是植物光合作用的重要原料,其浓度的升高可以对植物光合生理产生显著的影响[2-3]。CURE等[4]研究发现,短期内提高CO2浓度可以使小麦、玉米和大豆等作物的光合速率提高约50%。VU 等[5]研究认为,植物光合作用的最适温度范围会随着CO2浓度的升高而增加5~10 ℃。因而增施CO2被作为缓解温室高温逆境,促进作物高产优质栽培的重要技术措施[6]。但长期增施CO2常引起同化物,特别是淀粉在叶肉中积累,进而引发反馈抑制,降低光合速率,即发生所谓的“光合适应”[7]。调控同化产物的转运与分配是克服“光合适应”的重要途径。目前,关于高温与加富CO2耦合条件对作物库源动态影响的研究较少。本试验以盆栽番茄为试验材料,探究了温室长时间高温与加富CO2耦合条件下,番茄叶片光合日变化与同化产物的积累情况,旨在为合理利用增施CO2缓解高温胁迫对设施番茄生产的不利影响提供科学依据。
1 材料和方法
1.1 试验材料
试验所用番茄为无限生长型品种鸿途,具有良好的丰产性和抗病性。番茄幼苗购自山西巨鑫伟业农业科技开发有限公司,选择长势一致的五叶一心幼苗,移栽至营养钵(25 cm×25 cm)缓苗,钵内基质同育苗基质。
1.2 试验方法
试验在山西农业大学园艺学院园艺实验站的4 座自然光照气候室中进行。气候室坐北朝南,每座的长度、宽度、前屋面高度和后屋面高度分别为3、2.4、1.5和2.3 m。每座气候室中均安装有温度、CO2浓度和空气湿度传感器,传感器数据传输至SG605 温度和CO2浓度一体式控制器(北京烁光盛业科技发展有限公司)。控制器每10 min采集一次数据并根据试验设定要求自动打开或关闭空调机和CO2钢瓶使气候室环境稳定在设定水平。每天7:00—17:00进行温度和CO2浓度控制。试验设置常温(25~28 ℃)和高温(37~40 ℃)2个温度处理,每个温度处理下设置一个正常C02浓度(300~400 μmol/L)和一个高C02浓度(600~800 μmol/L),因此,试验共设置4个处理,分别记作常温(对照)、常温+CO2、高温和高温+CO2。阴雨天不进行处理。每个处理设置3 个重复,每个重复20 株番茄,试验共处理45 d。
1.3 测定项目及方法
1.3.1 光合气体交换参数的测定 处理45 d时,使用Li-6400 光合仪(美国LI-COR 公司)测定仪测定番茄植株的功能叶(由上向下数第4 片叶)的净光合速率(Pn)、胞间CO2浓度(Ci)、蒸腾速率(Tr)和气孔导度(Gs)。
1.3.2 生物量的测定 将番茄植株的地上部与地下部分开,用去离子水冲洗干净后用吸水纸轻轻擦干,然后将各部分放在烘箱内先用105 ℃进行杀青20 min,然后在85 ℃烘干至恒质量,最后用电子天平分别称取地上部和根系的干质量。
1.3.3 叶片中同化产物含量的测定 由上向下数第4片功能叶片的可溶性糖、淀粉含量分别采用蒽酮比色法[8]和碘滴定法[9]进行测定。
1.4 数据分析
数据使用SPSS 25.0 软件进行统计分析,采用Duncan法进行方差分析。用GraphPad Prism 6软件对数据进行作图。
2 结果与分析
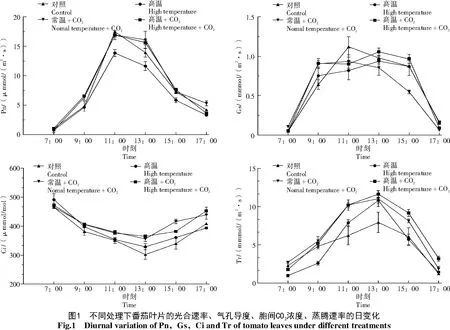
2.1 高温和增施CO2处理对番茄叶片光合参数日变化的影响
从图1可以看出,各处理叶片光合速率随着日出后光照增强而迅速升高,在11:00,所有处理的净光合速率出现峰值,但高温处理较对照低20.7%,差异显著(P<0.05)。在13:00,对照与单纯高温处理的Pn 快速降低,而常温+CO2与高温+CO22 个处理依然保持较高的Pn值。15:00,可能由于生长室侧壁遮光的影响,所有处理叶片的净光合速率均有较大幅度的下降,但高温处理仍然较对照低19.6%,差异显著(P<0.05)。高温+CO2处理可以缓解高温对光合作用的抑制,使9:00、11:00、13:00和15:00的番茄净光合速率较高温处理分别增加了42.2%、22.5%、34.3%和30.5%,差异显著(P<0.05)。
气孔导度(Gs)是气孔与大气间通量估算的重要参数。前人研究表明,气孔导度随温度升高而升高[10],但随CO2浓度升高而降低[11]。由图1 可知,各处理的气孔导度呈现先升高后下降的趋势,但各处理间差异不明显,仅在15:00时常温+CO2处理较对照低37.2%,差异显著(P<0.05),这可能是由于气孔导度对瞬时光合有效辐射(PAR)、叶片与空气间的水汽压亏损(VPD)、空气温度等反应敏感造成的[12]。
不同处理的Ci值日变化与Pn表现出相反的变化趋势,在一天中呈早晚高、中午低的变化趋势。高温+CO2和常温+CO2处理的全天Ci值均高于高温处理和对照。
蒸腾作用产生的拉力是植物水分吸收和运输的主要动力,与植物的光合作用关系密切。由图1可知,不同处理的番茄叶片Tr 值日变化呈单峰曲线,峰值出现在13:00。此时,高温处理Tr 值较对照高36.3%,差异显著(P<0.05),这可能是由于蒸腾作用随着温度的上升而增强造成的[13]。
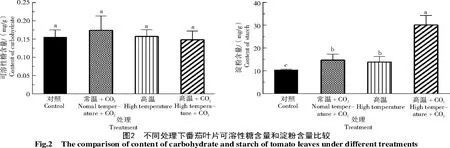
2.2 高温和增施CO2 处理对番茄叶片淀粉和可溶性糖含量的影响
可溶性糖和淀粉是主要的光合碳同化物[14]。图2 是试验期结束时的功能叶片可溶性糖含量和淀粉含量。由图2可知,所有处理的番茄叶片可溶性糖含量无显著差异。番茄叶片淀粉积累量在高温和高CO2浓度下显著增加:常温+CO2处理叶片淀粉含量比对照提高41.9%,高温处理比对照提高33.9%,差异显著(P<0.05);而高温+CO2处理又比高温处理的叶片淀粉含量增加116.9%,比对照则增加了190.5%。
2.3 高温和增施CO2 处理对番茄干物质积累的影响
干物质积累状况反映了植物的生长发育状况[15]。从图3可以看出,高温抑制了番茄的生长,高温处理比对照的总干质量降低了18.3%,差异显著(P<0.05);高温+CO2处理较常温+CO2处理的总干质量降低了18.6%,差异显著(P<0.05)。但高温+CO2处理比高温处理的总干质量增加了16.7%,差异显著(P<0.05),增加幅度较常温下增施CO2的效果(常温+CO2比对照总干质量增加30.6%)低,表明增施CO2部分缓解了高温胁迫对番茄生长的抑制作用。

3 讨论
作物叶片光合速率的高低与品种特性、温度、光照及CO2浓度等环境因子密切相关。本试验中,高温与增施CO2均对番茄的光合速率产生了显著影响。高温胁迫降低了番茄叶片的净光合速率,这与张洁等[10]的研究结果一致。这可能与高温胁迫影响了植物叶片光合系统的电子传递、光合磷酸化、暗反应酶活性,甚至造成光合机构损伤等有关[16]。在常温条件下,增施CO2显著提高了番茄叶片的Pn值,这与增施CO2增加了番茄叶片的胞间CO2浓度,进而增强了Rubisco的羧化作用有关[11,17]。同时,在高温条件下,增施CO2显著缓解了高温对番茄叶片光合作用的抑制,提高了午间高温时段的光合速率,这与朱世东等[18]在樱桃番茄上的研究结果一致。这可能与增施CO2在一定程度上增加了番茄光合对高温的适应性,从而部分缓解了高温胁迫的危害有关。前人研究表明,提高CO2浓度和环境温度对植物光合作用具有协同作用,植物CO2饱和点会随温度的升高而提高[19],而光合作用的最适温度范围也会随着CO2浓度的升高而提高[4]。
本试验中,高温和增施CO2对番茄叶片可溶性糖含量无影响,而对叶片淀粉含量的影响较大,这与薛义霞等[1]在番茄中的研究结果类似,这可能与植物存在通过调整源叶中的糖浓度以维持源库间一定的浓度梯度的适应性反应机制有关[14]。同时,本试验中在常温条件下增施CO2使叶片淀粉含量增加41.9%,但在高温条件下增施CO2使番茄叶片淀粉含量增加116.9%,表明高温下同化物输出率降低。可能是由于持续的高温胁迫造成植株韧皮部中的胼胝体增多,堵塞维管束,影响了同化物的输出造成的[10]。而淀粉的积累可能是导致高温下长期增施CO2后黄瓜净光合速率下降的重要原因[20]。但也有研究者得出了不同的结论,潘璐等[6]研究表明,高温处理促进了淀粉的分解,降低了高温处理下黄瓜叶片的淀粉含量。这除与作物和品种特性不同外,还可能与不同研究的空气湿度、昼夜温差等环境条件存在差异有关[1,21]。后续将进一步研究通过增施CO2缓解高温胁迫时,空气湿度、昼夜温差等环境条件对光合产物运转和分配的影响,进而为提高CO2的利用效率和经济效益提供参考。
4 结论
增施CO2可以缓解高温胁迫对番茄的抑制,增加生物量,但长期的CO2加富措施导致同化物输出率降低,淀粉在叶片中积累。番茄栽培中,在通过增施CO2缓解高温胁迫时,应进一步优化其他环控措施,提高同化物的输出与利用效率。
