福建大型海岛山体植物组成及其空间特征影响因素研究
2022-01-26谢艳秋曾纪毅邓传远
谢艳秋,黄 晖,2,曾纪毅,张 群,杨 铭,邓传远*
(1 福建农林大学 园林学院,福州350002; 2 福建水利电力职业技术学院,福建三明 366000)
海岛是生物多样性的天然储存池,海岛面积仅占地球陆地面积的3.5%,但却拥有陆地上15%~20%的物种,是保护生物多样性的关键地区[1]。物种丰富度或物种组成格局及其形成机制一直是生态学的核心研究问题之一,随着物种丰富度格局研究的深入,国内外许多学者提出了环境能量假说[2]、水分能量假说[3]、空间假设理论[4]以及历史假说[5]等用以解释物种组成、物种丰富度的分布格局,这些假说间通常并不排斥,往往共同解释了物种丰富度、物种组成格局的形成机制。岛屿生物地理学理论认为,众多因子或因子集(气候、空间、进化历史和物种间的相互作用)影响物种丰富度、物种组成格局,其中空间假设理论中的“岛屿理论”强调了面积效应和隔离效应,这两效应表明物理空间特征可能是影响植物丰富度与物种组成的重要因子集之一[6-8]。但越来越多的研究表明,除面积、隔离度外,海拔[9]、形状[10]、纬度[11]等空间特征对物种丰富度与物种组成具有重要的影响。因此,空间特征因子集解释物种丰富度与物种组成分布格局成为研究的热点领域。
岛屿的物种丰富度随着物种组成的变化而出现波动[12],生态位理论认为两个地区之间的物种组成差异与环境呈正相关[13]。不同地区间植物组成的差异,常用物种相似性系数,如Jaccard指数作为Beta多样性指数的衡量指标[14]。对不同地区植物组成差异之间的研究[15]可为今后海岛生态保护区建立的优先排序和生物入侵的管理提供信息[16]。此外,如果一定面积内物种丰富度高,则植物组成较为复杂,往往由不同的植物类群组成,如不同的生活型植物类群或不同的植物区系地理成分类型。不同植物类群丰富度的影响因子不同,有些植物类群丰富度受面积影响,而有些植物类群丰富度受海拔影响。除了探讨物种丰富度与空间特征的关系,最近的研究尝试用合适的统计模型分析不同植物类群与空间特征的关系[17-18]。
中国海岛地貌主要以丘陵、台地为主,大部分岛屿的山体面积占海岛面积三分之二及以上[19],一些小型无居民海岛其整个海岛就是一个完整的山体结构。目前对系列邻近的小型无居民海岛物种组成与对应岛屿空间特征的相互影响关系已有研究[20-22],但关于大型岛屿内系列山体与空间特征关系的研究依旧薄弱。海坛岛为中国第五大岛,地貌类型主要为丘陵、台地等山体组成[23],整个海岛分布有大小不一的山体(丘陵和台地),山体部分边界与海水相连,部分边界与岛陆相连,其空间特征既不同于内陆的山体(生境岛屿),也不同于小型无居民海岛(海洋岛屿),形成了特殊的生境和植被类型。然而随着平潭综合实验区的设立,海坛岛正经历快速的城市化进程,自然栖息地的改变和外来物种的引进威胁着海坛岛的生物多样性,因此在目前国家倡导的海岛植被保护与修复背景下,研究该区域植物分布格局及其形成机制具有重要意义。海坛岛植被覆盖率较高的区域多为山体,有鉴于此,本研究以海坛岛20座山体为研究对象,通过探讨海坛岛20座山体面积、海拔、周长、岸线长度等空间特征对物种丰富度格局的影响,并进一步分析空间特征对不同生活型、不同分布区类型植物组成、不同山体间物种相似性的作用。在城市发展与人为扰动加剧的背景下,以期为海坛岛及大型海岛生态安全建设和植物资源合理利用提供理论依据。
1 材料和方法
1.1 研究区概况
海坛岛是福建省第一大岛平潭岛的主岛,陆地面积324 km2,25°15′~25°45′N,119°32′~120°10′E[24]。海坛岛最高峰为海拔434 m的君山,岛上以丘陵、台地、平原为主,其山体地势陡峻,常见基岩裸露,且多沿海岸线分布[19]。土壤主要以砖红壤性土、盐土、风沙土为主。受热带气旋的影响,海坛岛年平均风速为5.0 m/s,7级以上大风日长达125 d,风速极值出现在7-9月台风期间[23]。岛上植被优势种明显,乔木层优势种主要为台湾相思(Acaciaconfusa)、木麻黄(Casuarinaequisetifolia)、黑松(Pinusthunbergii)、楝(Meliaazedarach)和桑(Morusalba)等。灌木层优势种主要为黑面神(Breyniafruticosa)、胡颓子(Elaeagnuspungens)、硕苞蔷薇(Rosabracteata)、算盘子(Glochidionpuberum)和滨柃(Euryaemarginata)等。草本层优势种主要为鬼针草(Bidenspilosa)、狗牙根(Cynodondactylon)、火炭母(Polygonumchinense)、白茅(Imperatacylindrica)和海边月见草(Oenotheradrummondii)等。
1.2 调查方法
选取海坛岛上植被保存较为完好的山体,于2018-2021年对海坛岛君山等20个山体进行实地调查(图1),采用样线和样方2种方法调查物种丰富度,样线法调查于每年的夏季进行,并于当年的秋季和次年的春季对调查的山体各进行一次补充调查,每座山体的调查次数共3次。样线法采用环线样线和十字形样线相结合的方法[25],每座山体布置1条环形样线和2条构成十字形的样线,每座山体3条样线,20座山体共调查60条样线,样线宽度10 m范围内调查植物种类。对山体典型阔叶林、针叶林和灌草丛以标准样方法设置样方,调查样方内物种数。森林样方按20 m×20 m设置,灌草丛样方按10 m×10 m设置,共设置102个森林样方和61个灌草丛样方。现场拍摄照片并采集标本,标本存放于福建农林大学园林学院植物标本室。参考《福建植物志》[26]《中国植物志》[27]对植物进行鉴定,存疑物种请专家鉴定,完成植物名录。
物种丰富度(species richness, SR)以物种数为准则,属种系数为植物属总数与种总数的比值。依据《中国植物志》[27]界定种子植物名录并参考吴征镒等[28]对种子植物地理成分进行分析。地理成分分析中热带成分属总数与温带成分属总数的比值为R/T。根据《福建植物志》对植物生境的描述加以区分筛选具有滨海特色的植物(地理分布上通常为滨海地区和海岛)。生活型依据Raunkiaer[29]的划分原则来区分,参考王梅峒[30]的生活型分类方式将高位芽植物进一步细分为大型高位芽植物、中型高位芽植物、小型高位芽植物、矮小型高位芽植物。
1.3 空间特征参数获取
采用Google Earth的“添加-多边形”工具勾画20个山体范围的图形,将数据保存为.kmz文件导入Acr GIS 10.3中转换为shp格式,导入DEM高程数据提取等值线,获得每座山体的最高点海拔。矢量数据导入Acr GIS 10.3使用“计算几何”工具获取各个山体的面积(误差±5 m2)、周长、岸线长度(表示山体临海一侧的海岸线长度)、距最近居民点最短距离等参数采用Google Earth的“标尺”工具获取。通过以上参数计算如下参数:植被覆盖率为山体植被覆盖面积与A的比值,周长面积比(perimeter area ratio, PAR)表征岛屿的边缘效应[31],形状指数(shape index,SI)表征山体形状的复杂程度,其公式为SI=Per/[2×(π×A)0.5][32]。20座山体的空间特征参数见表1。文中应用的8个参数中英文及英文缩略词如下:最高点海拔(maximum elevation,ME)、面积(area, A)、周长(perimeter, Per)、岸线长度(coastline length, CL)、距最近居民点最短距离(distance to nearest settlement, DNS)、植被覆盖率(vegetation coverage,VC)、周长面积比(perimeter area ratio, PAR)、形状指数(shape index, SI)。
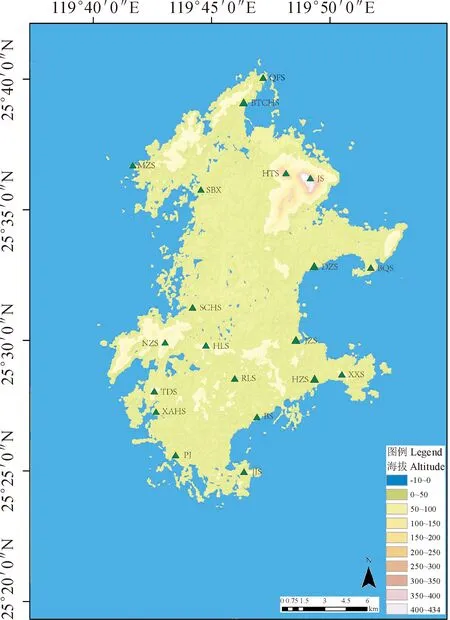
BS.北山;XXS.小雄山;HZS.猴仔山;JZS.金仔山;BTCHS.伯塘村后山;QFS.青峰山;SCHS.砂场后山;TDS.天大山;JJS.将军山;DZS.帝仔山;BQS.白犬山;HLS.后楼山;JS.君山;RLS.瑞岭山;XAHS.下澳后山;PJ.坪礁;NZS.牛寨山;SBX.蛇鼻下;MZS.猫子山;HTS.虎头山。下同图1 海坛岛山体调查地点分布图BS. Mount. Bei; XXS. Mount. Xiao Xiong; HZS. Mount. Hou Zi; JZS. Mount. Jin Zai; BTCHS. Mount. Bo Tang Village; QFS. Mount. Qing feng; SCHS. Mount. Sha Chang; TDS. Mount.Tian Da; JJS. Mount. Jiang Jun; DZS. Mount. Di Zi; BQS. Mount. Bai Quan; HLS. Mount. Hou Lou; JS. Mount. Jun; RLS. Mount. Rui Ling; XAHS. Mount. Xia Ao; PJ. Mount. Ping; NZS. Mount. Niu Zhai; SBX. Mount. She Bi; MZS. Mount. Mao Zi; HTS. Mount. Hu Tou. The same belowFig.1 Distribution map of investigated mountains on Haitan Island

表1 海坛岛20个山体的基本信息
1.4 数据处理分析
运用多元线性回归分析拟合物种丰富度与空间特征的多元回归模型,运用基于赤池信息准则(akaike information criterion, AIC)的逐步回归方法,筛选出最小AIC值的最佳回归模型,并通过方差分解阐明海坛岛山体物种丰富度与空间特征关系的权重。
除了物种丰富度的格局之外,本研究还对不同的植物类群(不同生活型、属的不同区系地理成分类型)使用Spearman相关性分析植物类群与空间特征的关系。为有效反映进化历史不同的植物组成格局及影响因素的差异,参考《中国种子植物区系地理》[28]将种子植物按照属的分布区类型划分为世界广布、热带(2-7区系成分)和温带(8-14区系成分)3种区系成分。应用多元回归、逐步回归与方差分解来解释空间特征对不同生长型、不同属的区系地理成分类型的影响。
采用Jaccard相似性系数计算山体间物种组成相似性,表征不同山体间的Beta多样性[33]。其公式为:
其中,j代表不同山体间共有的物种数,a、b分别代表不同山体包含的物种数。山体间两两计算空间特征比值与Jaccard相似性系数,采用Spearman相关性分析山体间植物Beta多样性的影响因素。
数据分析主要采用R 4.1.0,使用psych包和correlation包进行Spearman相关性分析、packfor包进行逐步回归、relaimpo包进行方差分解分析重要性,用ggplot2包作图[34]。
2 结果与分析
2.1 种子植物组成及植物类群分析
2.1.1 种子植物组成分析海坛岛20个山体共记录种子植物541种(含种下分类),隶属于110科381属,海坛岛山体间物种丰富度差异较大,最多的君山达308种,最少的金仔山仅29种。其中,种类最多的前3个科分别是菊科(Compositae)、禾本科(Gramineae)与豆科(Leguminosae)。种间频度差异较大,苦郎树(Clerodendruminerme)、补血草(Limoniumsinense)、珊瑚菜(Glehnialittoralis)等均仅见于1个山体,而台湾相思(Acaciaconfusa)、鬼针草(Bidenspilosa)、鸡矢藤(Paederiafoetida)则均出现在20个山体中。海坛岛20个山体中属种系数最大的为小雄山(98.70%),属种系数较小的是君山(81.49%)。受海岛特殊自然条件的影响,海坛岛山体具有丰富的海岛特色植物,其中肾叶打碗花(Calystegiasoldanella)、滨海珍珠菜(Lysimachiamauritiana)、厚藤(Ipomoeapescaprae)、单叶蔓荆(Vitexrotundifolia)等主要分布在滨海砂生生境。海滨藜(Atriplexmaximowicziana)、南方碱蓬(Suaedaaustralis)等主要分布于盐生湿地生境。滨柃(Euryaemarginata)、补血草(Limoniumsinense)、滨海前胡(Peucedanumjaponicum)等主要分布于基岩海岸。细枝叶下珠(Phyllanthusleptoclados)、海岸扁担杆(Grewiapiscatorum)等主要分布于海岸林生境。
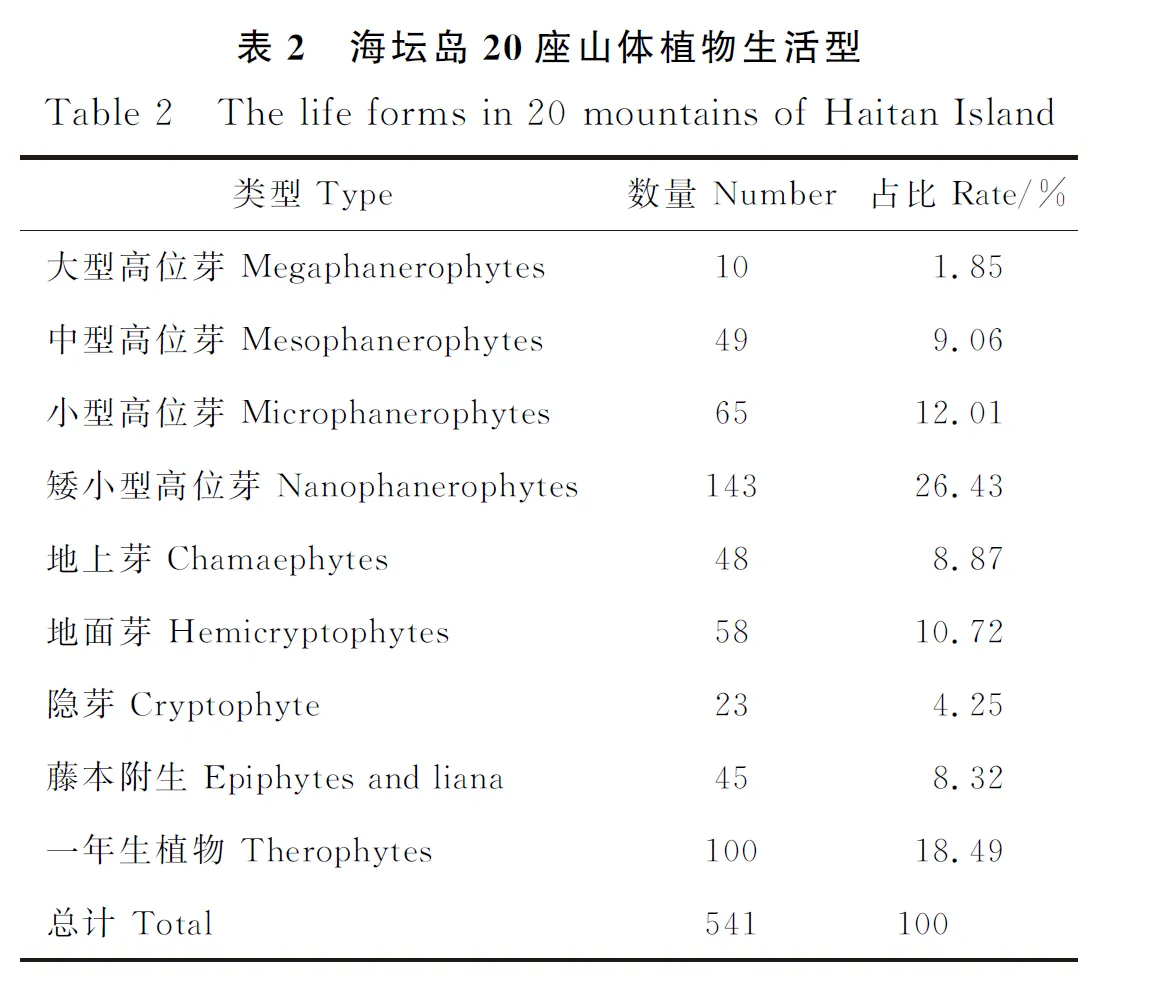
2.1.2 植物生活型类型分析根据海坛岛20座山体总的植物名录,分析山体植物生活型类型(表2)。从表1可知,海坛岛山体植物生活型以矮小型高位芽植物为主,占总物种数的26.43%。矮小型高位芽主要为蔷薇科(Rosaceae)、鼠李科(Rhamnaceae)、马鞭草科(Verbenaceae)、山茶科(Theaceae)等。一年生植物次之,占总物种数的18.43%,主要为禾本科、菊科、莎草科(Cyperaceae)、旋花科(Convolvulaceae)等。而大型高位芽占比最低,仅占比1.85%,主要为松科(Pinaceae)。
2.1.3 属的区系地理成分类型分析根据海坛岛20座山体总的植物名录,分析山体属的区系地理成分类型(表3)。从表3可知,种子植物381属可分15个分布区类型。海坛岛20个山体种子植物中世界分布有49属(世界广布性因不具地区代表性,而不纳入总属数比例计算,下同)。分布中心为热带的有218属,占总属数的65.67%,主要为榕属(Ficus)、木防己属(Cocculus)、脚骨脆属(Casearia)、打碗花属(Calystegia)、栀子属(Gardenia)、黄花稔属(Sida)、紫珠属(Callicarpa)等。分布中心为温带的有112属,占总属数的33.73%,主要为蒿属(Artemisia)、胡颓子属(Elaeagnus)、桑属(Morus)、接骨木属(Sambucus)、忍冬属(Lonicera)、蔷薇属(Rosa)等。中国特有分布仅2属,占总属数的0.6%,分别为杉木属(Cunninghamia)和枳属(Poncirus)。海坛岛20个山体种子植物属的R/T值为1.94。
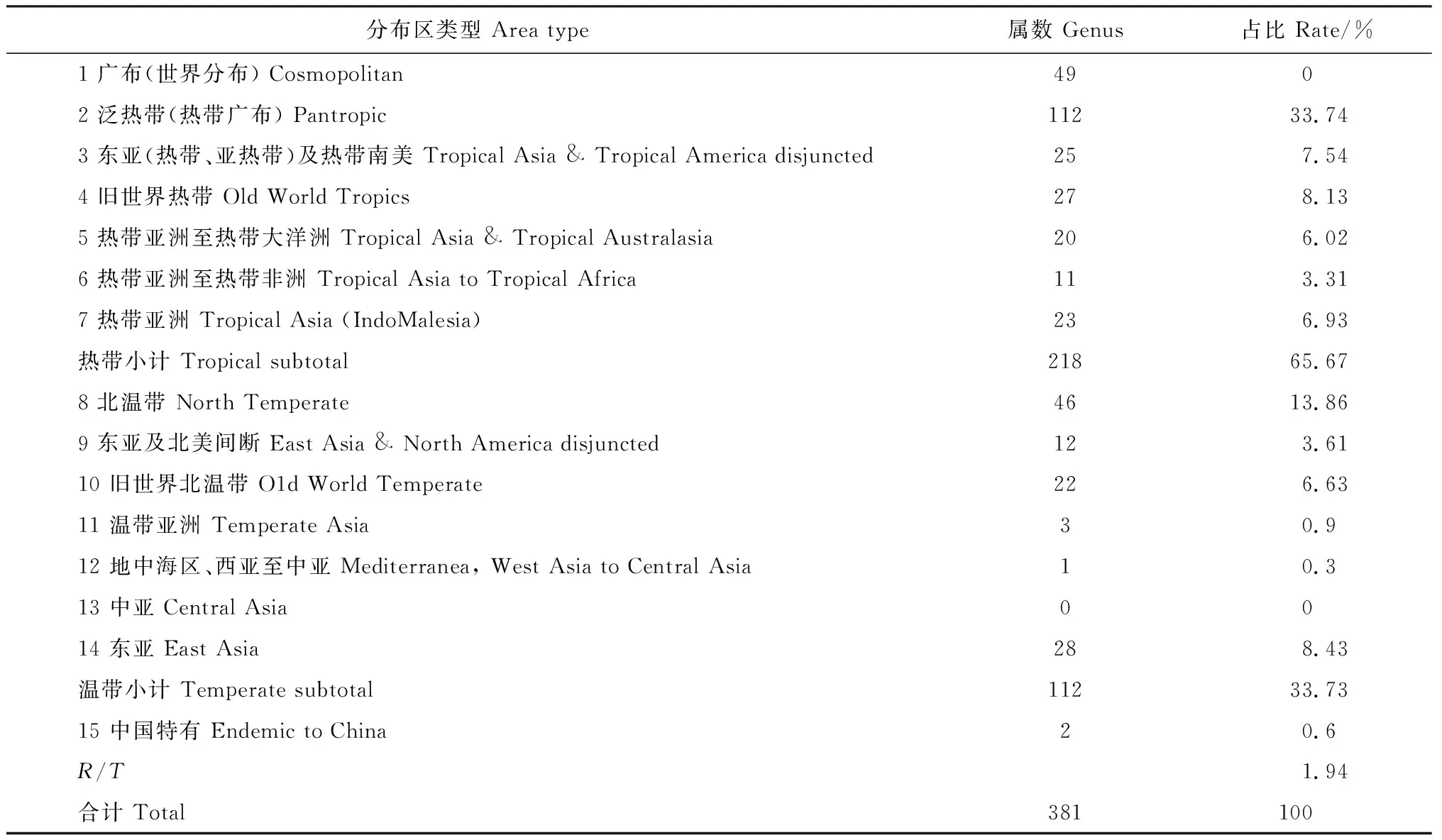
表3 海坛岛的山体种子植物属的区系地理成分
2.2 物种丰富度与空间特征的关系
以每座山体的物种丰富度(物种数)及对应的空间特征参数进行多元线性回归分析,通过逐步回归判定适合总物种丰富度的模型,以探究与空间特征之间的关系,得到的最优模型为:SR=0.32ME+253.45CV-3.63PAR+41.094SI,AIC值为181.76,调整后的R2为0.92。通过方差分解可知,海拔对总物种丰富度具有较高的解释率,重要性达38.11%,而形状指数仅占总物种丰富度重要性的2.05%,具有较低的解释率。可知海拔、植被覆盖率、周长面积比、形状指数是物种丰富度格局的主要影响因素。
2.3 生活型组成、属区系地理成分组成与空间特征的关系
以每座山体的不同生活型物种丰富度及对应的空间特征参数进行逐步回归分析,并经方差分解分析(图2,A)。海拔是显著影响大型高位芽物种丰富度的因子,通过逐步回归可知大型高位芽的最优模型为0.579312+0.017033ME,调整后的R2为0.73,海拔是大型高位芽的唯一影响因素,因此无法进行下一步的方差分解分析。由上可知,在20座山体中,随海拔的升高,大型高位芽的物种丰富度逐步增加(P<0.05)。
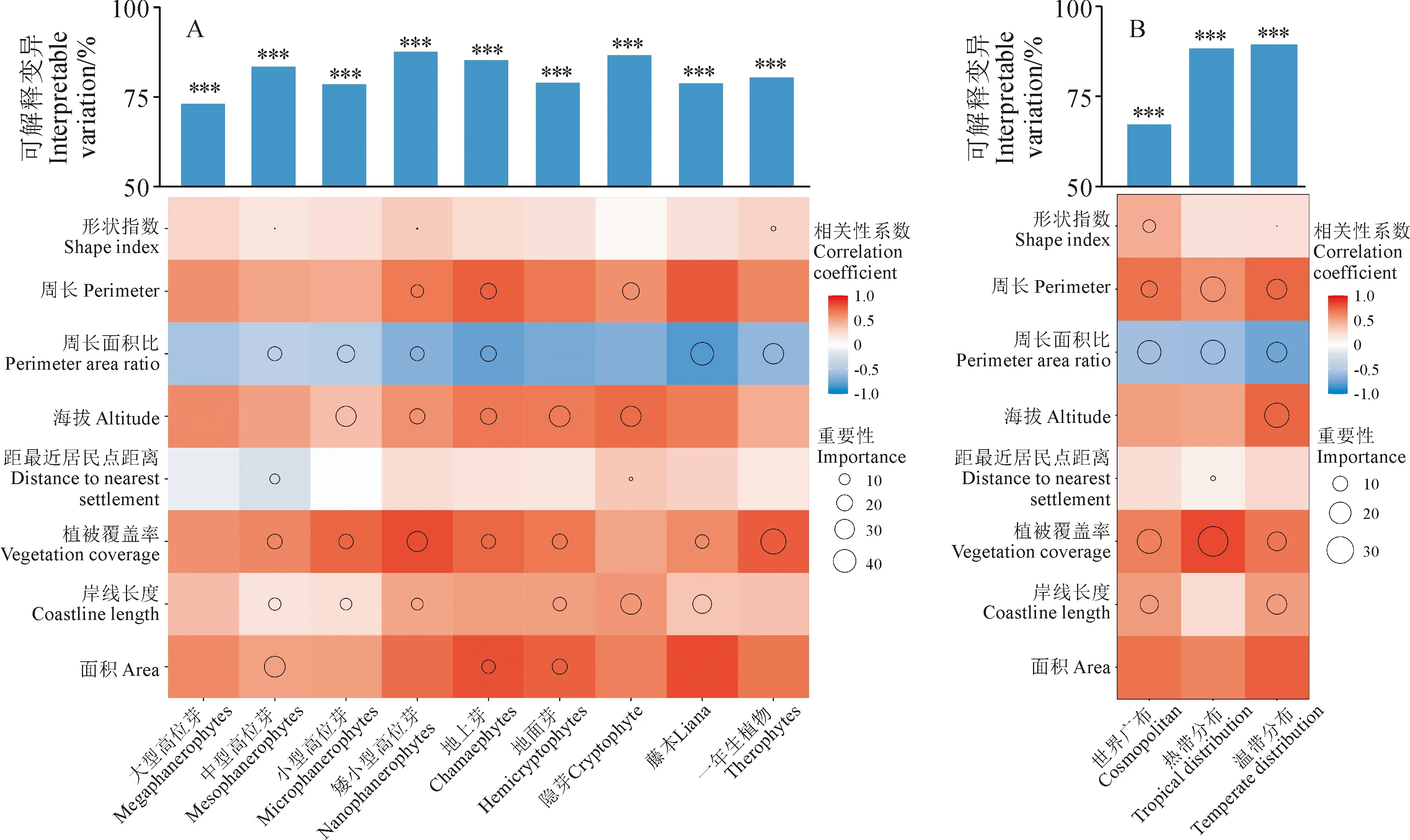
A表示空间特征对生活型的影响;B表示空间特征对属的区系地理成分的影响。颜色表示Spearman相关性,红色表示正相关,蓝色表示负相关,颜色越深表示相关性越强。柱状图为空间特征对因变量变异的总解释率,***P<0.001;圆圈的大小表示方差分解后各变量的重要性图2 空间特征对生活型和区系地理成分的影响A shows the effects of spatial characteristics on life forms; B shows the effects of spatial characteristics on areal types of genera. The color indicates the Spearman correlation, the red indicates the positive correlation, the blue indicates the negative correlation, and the darker the color, the stronger the correlation. The histogram shows the interpretable variation rate of spatial characteristics to dependent variable, ***P<0.001; The size of the circle indicates the importance of each variable after variance decompositionFig.2 Effects of spatial characteristics on life forms and areal types
面积、周长面积比、距居民点最近距离、岸线长度、植被覆盖率、形状指数显著影响中型高位芽物种丰富度,其总解释率为78%。随面积、距居民点最近距离、岸线长度、植被覆盖率、形状指数的增大,中型高位芽物种丰富度显著增加(P<0.05),随周长面积比的增大,中型高位芽物种丰富度显著降低(P<0.05)。经方差分解分析,面积对中型高位芽具有最大的解释率,重要性达33.72%,其次是植被覆盖率和周长面积比,重要性分别为17.18%和15.56%,而形状指数的重要性仅为1.67%。
以每座山体的不同分布区类型的属数及对应的空间特征参数进行逐步回归分析,并经方差分解分析(图2,B)。周长、周长面积比、岸线长度、植被覆盖率、形状指数显著影响世界广布类型的属数,总解释率为75%。随周长、岸线长度、植被覆盖率、形状指数的增大,世界广布类型的属数逐渐增多(P<0.05),随周长面积比的增大,世界广布类型的属数逐渐减少(P<0.05)。经方差分解分析,植被覆盖率的重要性最大,达24.55%,其次是周长面积比和岸线长度,重要性分别为23.10%和14.34%,而形状指数仅占7.69%的重要性。
2.4 山体间物种组成相似性与空间特征的关系
根据Jaccard相似性指数(表4)可知海坛岛20个山体中,君山与虎头山的物种组成相似性最高,Jaccard指数为0.83;物种组成相似性最低的为金仔山与君山、金仔山与虎头山,Jaccard指数为0.07。
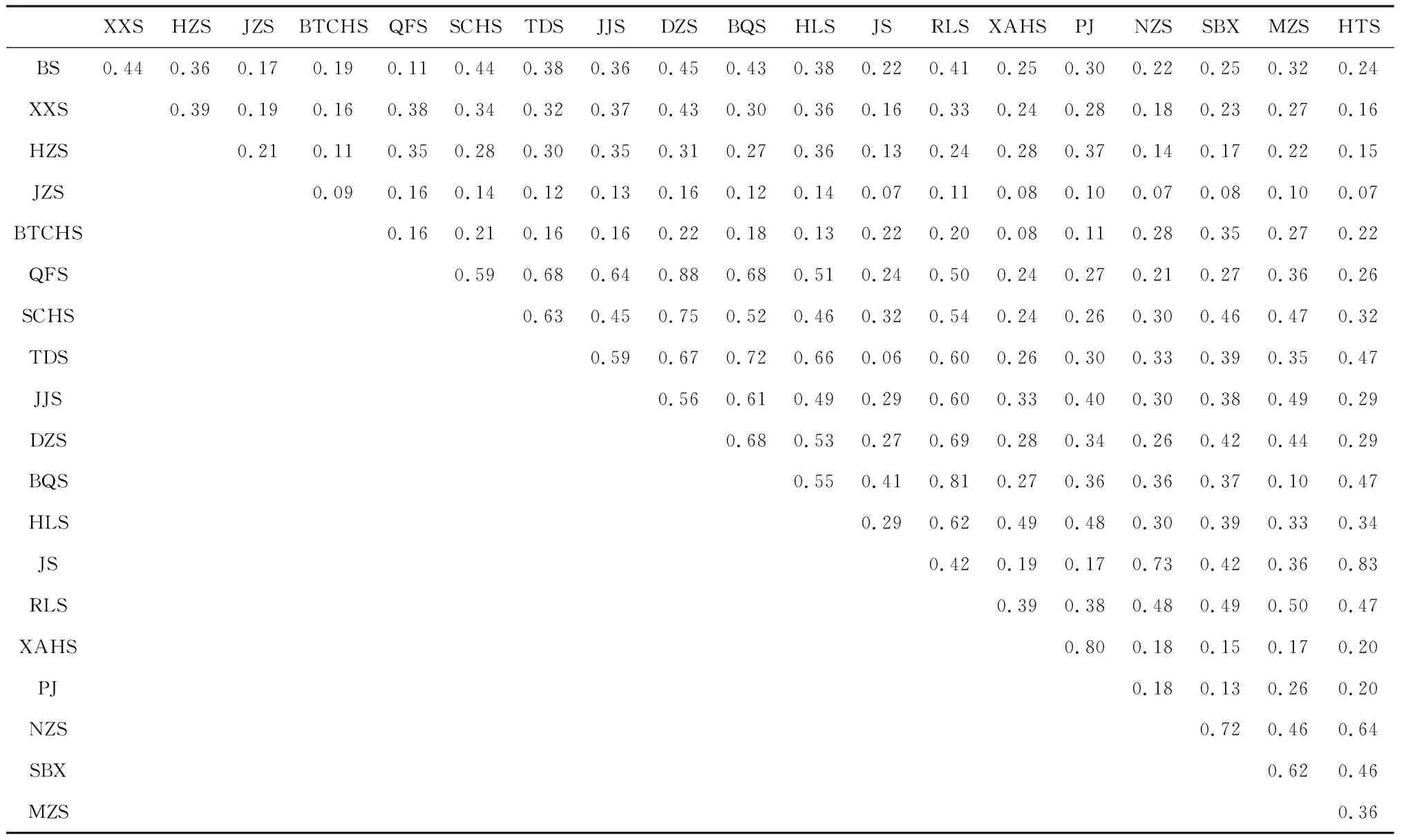
表4 海坛岛20座山体之间物种组成的Jaccard相似性系数
使用Spearman相关性分析解释山体空间特征对山体间物种组成相似性的影响(表5),Cj与空间特征参数之间的相关分析表明,Cj与面积比、海拔比存在极显著正相关关系(P<0.001),而与周长面积比比值之间存在显著负相关关系(P<0.01)。

3 讨 论
3.1 海坛岛山体植物生活型的特点
海坛岛20个山体属种系数平均值为92.02%,说明该地区单种属较多,表明海岛山体生境恶劣。同属植物起源与进化趋势通常近似[30],往往因为生态位较一致而产生强烈的竞争,导致海岛山体单种属较多。生活型是植物对环境条件的长期适应[35],海坛岛山体植物生活型以矮小型高位芽、一年生植物为主。海坛岛山体现有植物群落占比最大的是台湾相思群系和木麻黄群系等,属于20世纪飞机播种后构建形成的半自然群落,矮小型高位芽植物主要在以台湾相思、木麻黄等大型高位芽为建群种的林下分布。鉴于此,矮小型高位芽植物生长受干燥气候、强风、盐雾和土壤贫瘠的影响远低于大中型高位芽[36],由此可推测,适宜的生境是矮小型高位芽在海坛岛山体占比高的原因。而一年生植物由于体积小、生命周期短、扩散能力强,与体型较大的物种相比,面临更低的灭绝风险[37],因此在海坛岛山体占比较高。
以往的研究表明随着植物生态序列的正向演替,亚热带常绿阔叶林生活型表现为高位芽植物占比为主,而一年生植物相对较少[38]。经卡方检验,海坛岛山体(高位芽植物49.35%、一年生植物18.48%)与邻近大陆山体梅花山(高位芽植物81.20%、一年生植物2.80%)、武夷山(高位芽植物74.00%、一年生植物4.00%)[39]、南靖六斗山[40](高位芽植物83.30%、一年生植物3.70%)之间存在极显著差异(P<0.001)。海坛岛山体高位芽植物比例下降,而一年生植物比例上升,植被表现出明显的次生特征,这可能是因为随着原生植被的破坏和次生植被的恢复以及海岛恶劣的生境(强风、盐雾以及土壤贫瘠)影响,高位芽植物的生态位较少,而一年生植物由于生活史短、迁徙能力较强[41],具有更多样化的生态适应性[39],受上述环境的影响较小,得以快速繁殖占据新的生态位。
3.2 海坛岛山体植物属分布区类型的特点
植物属的区系地理成分具有相对稳定的特点,因此属的地理成分更能清晰地反映植物的演化历程与发育环境[42]。中国亚热带地区归属于东亚区系,植物区系以泛热带分布和北温带分布为主[43]。R/T值越大,表明该地区植物地理成分热带性质越显著,海坛岛山体的R/T值为1.94,这说明热带分布植物是构成海坛岛20个山体种子植物的主要成分,温带分布植物是该地区植物区系的补充,符合其气候特征与所处亚热带地理特点。海坛岛山体属的分布区类型(热带成分65.67%、温带成分33.73%)与同纬度大陆山体福建梁野山[44](热带成分60.82%、温带成分36.72%)、福建梅花山[39](热带成分64.74%、温带成分31.41%)之间无显著差异(P>0.05),对比发现海坛岛山体植物地理成分与同纬度山体类似,具有由中亚热带向南亚热带过渡的特点。
3.3 空间特征对海坛岛山体植物组成的影响
种面积关系和岛屿生物地理学理论的研究认为,面积越大,能分化出更多的生境和新物种,物种丰富度将越高[45]。本研究对20座山体物种丰富度与8个空间特征参数的逐步回归表明,随山体面积的增大,20座山体物种丰富度提高没有达到显著水平,可能存在以下原因:
(1) 可能存在“小岛屿”效应,小岛屿效应指的是,当岛屿面积小于某个特定阈值,岛屿物种丰富度不受面积影响,其不确定性的倾向增加[1]。本研究中,研究的20个山体面积差异较大,面积最大的君山达1 587 hm2,而面积最小的金仔山仅为3.4 hm2,同时面积小于20 hm2的山体共8座,其物种丰富度差别较大,如面积接近的伯塘后山(11.42 hm2)和下澳后山(11.54 hm2),物种丰富度分别为92和62。
(2) 君山与牛寨山面积差异较大,君山的面积为牛寨山6.01倍,但君山和牛寨山的物种丰富度分别为308和227,君山的物种丰富度仅为牛寨山的1.36倍,这可能是20座山体面积多元线性回归没有达到显著水平的原因。根据物种丰富度分布格局的种库假说[46],假设20座山体能生长的潜在种库物种数为541种,而20座山体中有些山体,如君山植被次生性质明显,还未形成稳定的植物群落,在群落发育过程中,通过持续的增加物种数,将使物种数和面积的线性关系达到显著水平。
(3) 种面积关系和岛屿生物地理学理论都认为,面积增大导致物种丰富度增加的原因是,更大的面积将包含更多类型的异质生境,而随着异质生境类型的增多,新物种的出现概率也随之增加[47]。本研究中,植被覆盖率和山体面积因为共线性关系,逐步回归分析中能代表异质生境的可能是山体的植被覆盖率,逐步回归分析表明,随山体植被覆盖率的提高,20座山体物种丰富度增加达到极显著水平,同时,随山体植被覆盖率的提高,20座山体不同生活型(中型高位芽、小型高位芽、矮小型高位芽、地上芽、地面芽、藤本和一年生植物)物种丰富度增加达到显著水平或极显著水平。在海岛山体中,用植被覆盖率代替山体面积来代表生境多样性更为合适。因为海坛岛山体主要特征为大面积的裸露基岩和沙滩,这些生境分布的植物较少,同时君山、牛寨山这些面积相对较大的山体,邻近村庄,并有水泥道路的布设,这些因素使用面积来表示植物生长生境多样性不太准确。
逐步回归分析表明,20座山体随着周长面积比的增大,物种丰富度逐渐变小(P<0.05),不同生活型(小型高位芽、矮小型高位芽、地上芽、藤本和一年生植物)植物的物种数量逐渐减少(P<0.05),世界分布、热带分布和温带分布的属数也逐渐减少(P<0.05)。周长面积比参数是指示山体边缘效应的一个指标,根据逐步回归分析结果可推测,海坛岛20座山体边缘效应对物种丰富度的影响较小,对山体物种丰富度影响大的主要部分是山体内的植物群落。边缘效应对物种丰富度影响小的另一个原因可能是,大部分山体临海一面为物种分布较少的悬崖峭壁、沙滩或滩涂。同理,随周长面积比比值的增大,山体间Beta多样性减少可能也是因为山体临界处很多是无法生长植物的恶劣生境。
海拔梯度将引起各种环境因子的梯度变化,是物种丰富度变化的一个重要因子[48],内陆已有的研究表明,在云南丽江地区,受中域效应的影响,在海拔梯度近4 500 m范围内,随海拔的升高,物种丰富度呈现先增加后减少的单峰变化趋势[49-50],本研究的逐步回归分析表明,海坛岛20座山体,随海拔升高,物种丰富度呈现直线上升的趋势(P<0.05),不同生活型(大型高位芽、小型高位芽、矮小型高位芽、地上芽、地面芽和隐芽)植物的物种数量呈直线增加趋势(P<0.05)。出现不一致的原因可能是本研究的山体海拔梯度范围小,最高海拔为君山的434 m,最低海拔为北山的15 m,受中域效应影响的程度较小[51]。逐步回归分析表明,随海拔上升,温带分布类型的属增加,由于山体植物的属种系数高,因此,随着海拔的升高,温带分布类型的种类可能增多,这是高海拔物种丰富度更高的一个原因。逐步回归分析表明山体形状指数也是影响物种丰富度的一个因子,但方差分解分析表明其重要性较低,因此二者间的关系需进一步研究。类似地,还有一些因子对不同生活型植物物种丰富度的相关性也需进一步研究。
4 结论与建议
海坛岛20座山体植物以具有较强生态适应性的矮小型高位芽和一年生植物为优势,分布区类型具有明显的热带性质。海拔和周长面积比是海坛岛山体总的物种丰富度与不同类群植物物种丰富度的的主导因素。在本研究中,小岛屿效应、生境异质性等因素可能是导致海坛岛山体物种丰富度与植物组成受面积效应影响较小的原因。海坛岛山体间Beta多样性主要受面积比、海拔比以及周长面积比的影响。本研究进一步解释了空间特征参数在植物组成及物种丰富度分布格局中的影响机制,今后的研究可引入土壤理化性质、温度、降水等因素,或将有助于更全面深入地理解植物组成及物种丰富度分布格局。
此外,海坛岛山体具有丰富的滨海特色植物,如滨海珍珠菜(Lysimachiamauritiana)、海滨藜(Atriplexmaximowicziana)、滨柃(Euryaemarginata)、肾叶打碗花(Calystegiasoldanella)等,这些植物既适应海岛的气候与土壤,又具有滨海特色与观赏价值,可以考虑将其引种驯化为园林植物,建设海坛岛成为生态环境佳,兼具特色的观光型海岛。
