不同光温条件对翠碧一号烤烟碳氮代谢的影响
2022-01-21王月敏柯玉琴李文卿李春英钟月仙郑朝元魏晓玲
王月敏,柯玉琴,李文卿*,李春英,钟月仙,郑朝元,魏晓玲
(1.福建省烟草专卖局 烟草科学研究所,福建 福州 350013;2.福建农林大学 生命科学学院,福建 福州 350002;3.广东省汕头市农业科学研究所,广东 汕头 515021;4.福建农林大学 资源与环境学院/国际镁营养研究所,福建 福州 350002)
光照和温度是植物生长过程中重要的影响因素。光是光合作用的能量来源,也是形成叶绿素的必要条件,还调节碳同化中许多酶的活性[1]。光照强度对植物光合作用及生长代谢起着至关重要的作用[2]。烟株生长发育对环境温度敏感,温度不仅改变烟叶生长发育及组织形态,对物质代谢及色素积累也有重要影响[3]。生育期内温度过高,会破坏烟叶的叶绿素形成,影响光合作用,导致代谢失调,对烟株生长发育及烟叶的产品质量造成不利影响[4-8]。烟草是喜光作物,充足的日照有利于烟株生长,但是烟株生长期内温度过高时,叶绿素受到破坏,影响光合作用,烟叶品质降低,温度和光照只有保持一定平衡才能协调发挥作用,日照充足而不强烈对烟叶生长更为有利[4,9]。烟株处于高温和强光环境条件下时,首先发生变化的是其内在的生理过程,其中碳氮代谢酶的活性在适应逆境的物质和能量转化过程中起着重要作用。不同植物在环境压力下,碳氮代谢酶活性呈现不同趋势,一般来说,高温强光下植物体内与碳代谢相关的SS、SPS酶活性升高。NR、GS、GOGAT酶活性与氮代谢息息相关,NR对温度十分敏感,大多数植物的NR在环境压力下活性降低。GS/GOGAT循环是植物体内铵态氮同化的主要过程,产生的谷氨酰胺和谷氨酸是植物体内很多含氮物质的前体,此循环酶活性在不同环境因素和品种下呈现不同趋势[10-14]。高温条件容易引起植物体内蛋白质的分解,导致游离氨基酸含量的增加[15]。
福建烟区属于亚热带季风气候,主要种植春烟,随着生育期的推迟,烟株对应的温度和光照逐渐增强,到上部叶成熟采收时往往遇上高温和强光天气。高温和强光会对烟叶的成熟产生诸多不利的影响,尤其是在生长后期,高温强光的气候条件容易导致中上部烟叶出现“高温逼熟”的现象。通过光强与温度调控来研究烟株光合色素含量、碳氮代谢酶活性及代谢产物含量,不仅可以揭示烟株物质生产、分配以及生理代谢过程,而且可以反映烟株逆境适应能力,对烟叶品质形成的机理进行解析,以及对烤烟生产过程中如何合理调节生育期具有重要的指导意义。目前研究较多的是温度或光照单一胁迫对烤烟生长发育及生理代谢的影响,对于高温强光交互胁迫对成熟期烤烟生理代谢的研究鲜见报道。本试验通过设置不同光温处理,探讨了高温强光对成熟期烟叶的生理过程的影响,以期为烤烟的生产措施制定提供参考。
1 材料与方法
1.1 试验地点与供试材料
选用当地主栽烟草品种翠碧一号为试验材料,由福建省烟草专卖局烟草农业科学研究所提供。试验于2018年在福建省烟草专卖局烟草科学研究所宦溪科研基地进行。
1.2 试验处理
采用盆栽土培,花盆上口直径28 cm,下底直径22 cm,高25 cm,每盆装15.0 kg土壤。土壤为壤土,pH值为5.11,碱解氮含量为102.4 mg/kg,速效磷含量为16.8 mg/kg,速效钾含量为94.8 mg/kg,有机质含量为16.2 g/kg。每盆施用烟草专用肥20 g(N∶P2O5∶K2O=12%∶7%∶22%)。烟苗长到七叶一心时移栽,在室外露天种植;烟株打顶后1周移入植物生长箱(武汉瑞华仪器设备有限责任公司)进行高温和强光处理,处理时间为48 h。试验共设4个处理,每个处理重复4次。
T1:25 ℃遮光,光照强度为1600 μmol/(m2·s);在T2处理烟株测定的叶片上,用硫酸纸将倒2~倒3叶遮住半片叶片(硫酸纸遮光率为30%),用于取样测定生理指标;
T2:25 ℃强光,光照强度为1600 μmol/(m2·s);
T3:35 ℃遮光,光照强度为1600 μmol/(m2·s);在T4处理烟株测定的叶片上,用硫酸纸将倒2~倒3叶遮住半片叶片,用于取样测定生理指标;
T4:35 ℃强光,光照强度为1600 μmol/(m2·s)。
1.3 测定方法
烟株在植物生长箱中处理48 h后,取出烟株,分别取各处理倒2~倒3叶,剪碎混匀后用于测定各项生理指标。光合参数采用CIRAS-II型(由英国PP-system公司生产)便携式光合仪测定;叶绿素荧光参数采用Hansatech 公司生产的Pocket PEA便携式叶绿素荧光计测定;叶绿素含量参照张宪政[16]的丙酮浸提法进行测定;二磷酸核酮糖羧化酶(RuBPCase)活性应用上海酶联生物科技有限公司生产的试剂盒采用酶联免疫分析(ELISA)法进行测定;硝酸还原酶(NR)活性测定采用活体法[17];谷氨酰胺合成酶(GS)活性参照邹琦[17]的方法测定;蔗糖磷酸合成酶(SPS)和蔗糖合成酶(SS)活性参照刘以前等[18]的方法测定;蔗糖、还原糖、果糖、可溶性糖、淀粉等含量的测定方法参照《现代植物生理学实验指南》[19]。
烟叶中蛋白质水解氨基酸含量的测定方法:准确称取叶片干粉30 mg于1.5 mL离心管中,加入1 mL 80%甲醇,超声溶解30 min,以12000 r/min 4 ℃离心20 min,吸取上清液,用0.22 μm滤头过滤,用高效液相色谱仪进行测定,仪器型号为Waters XEVO-TQS。
2 结果与分析
2.1 不同处理对叶片光合色素含量的影响
叶绿体色素是植物体内进行光合作用的重要色素。从表1可见,35 ℃强光处理的Chla和Chl含量极显著低于其它3个处理,其它3个处理差异不显著。35 ℃遮光处理Chlb含量最高,显著高于35 ℃强光处理和25 ℃强光处理;其次是25 ℃遮光处理,其Chlb含量极显著高于35 ℃强光处理。不同光照处理的Chlx.c含量在25 ℃温度时差异不显著;25 ℃温度的两个光处理Chlx.c含量极显著高于35 ℃两个光处理;在25 ℃条件下,遮光导致Chl和Chlx.c含量下降;在35 ℃高温下遮光导致Chl和Chlx.c含量上升。说明在25 ℃适宜温度条件下,短期强光处理会激发Chl和Chlx.c色素含量增加,而高温处理显著导致烟株叶片总叶绿素和类胡萝卜素的大量降解。

表1 不同光温处理对叶片光合色素含量的影响
2.2 不同处理对碳代谢酶活性的影响
2.2.1 不同处理对叶片蔗糖合成酶活性的影响 从图1可见:25 ℃温度条件下,遮光导致叶片SS酶活性极显著下降;35 ℃温度条件下,遮光导致叶片SS酶活性极显著上升。遮光条件下,35 ℃温度处理的叶片SS酶活性极显著高于25 ℃温度处理;强光条件下,35 ℃温度处理的叶片SS酶活性极显著低于25 ℃温度处理。说明25 ℃适宜温度条件下,短期强光处理导致叶片SS酶活性显著上升,促进叶片蔗糖的合成;35 ℃温度条件下强光处理则明显抑制了叶片SS酶活性;遮光条件下升温导致SS酶活性上升,强光条件下升温导致SS酶活性下降。

图1 不同光温处理对打顶烟株SS活性的影响
2.2.2 不同处理对叶片蔗糖磷酸合成酶活性的影响 从图2可见:25 ℃温度条件下,遮光导致叶片SPS酶活性极显著下降;35 ℃温度条件下,遮光导致叶片SPS酶活性极显著上升。在相同光条件下,35 ℃温度处理的叶片SPS酶活性均显著高于25 ℃温度处理。说明25 ℃适宜温度条件下,短期强光处理导致叶片SPS酶活性显著上升,促进叶片蔗糖的合成;35 ℃温度条件下强光处理则极显著抑制了叶片SPS酶活性;两种光条件下升温均导致SPS酶活性上升。

图2 不同光温处理对打顶烟株SPS活性的影响
2.2.3 不同处理对叶片淀粉合成酶活性的影响 从图3可见:25 ℃温度条件下,遮光导致叶片淀粉合成酶活性极显著下降;35 ℃温度条件下,遮光导致叶片淀粉合成酶活性极显著上升。在相同光条件下,35 ℃温度处理的叶片淀粉合成酶活性均极显著高于25 ℃温度处理。说明25 ℃适宜温度条件下,短期强光处理导致叶片淀粉合成酶活性显著上升,促进叶片淀粉的合成;35 ℃温度条件下强光处理则极显著抑制了叶片淀粉合成酶活性;两种光条件下升温均导致淀粉合成酶活性上升。

图3 不同光温处理对烟草淀粉酶活性的影响
2.3 不同处理对叶片碳代谢物质的影响
从表2可见,无论25 ℃还是35 ℃温度条件,强光条件下烟叶中蔗糖、还原糖、可溶性糖和淀粉等的含量均显著或极显著高于遮光处理;果糖含量在25 ℃温度条件下,强光处理显著高于遮光处理,35 ℃温度条件下,强光处理高于遮光处理,但差异不显著。相同光照条件下,35 ℃温度处理的蔗糖、还原糖和可溶性糖等的含量均显著或极显著高于25 ℃温度处理;果糖含量仅在遮光条件下,35 ℃处理显著高于25 ℃处理,强光条件下随温度上升果糖含量上升但差异不显著;相同光处理条件下淀粉含量随温度上升表现为显著下降。说明不同温度条件下,增强光照均可促进烟株糖类物质和淀粉的合成积累,相同光照条件下从25 ℃升到35 ℃可以促进糖类物质含量的增加,但导致淀粉含量的下降。
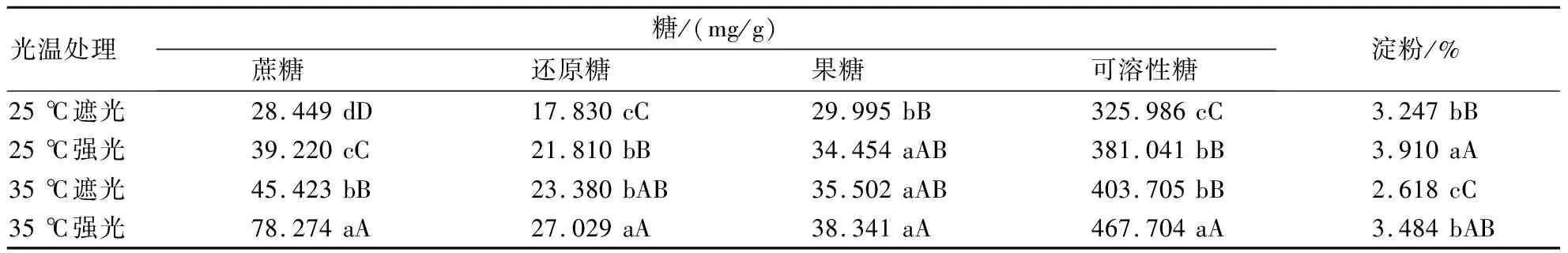
表2 不同光温处理对叶片糖和淀粉含量的影响
2.4 不同处理对叶片氮代谢酶活性的影响
2.4.1 不同处理对叶片NR活性的影响 从图4可见,25 ℃强光处理的烟叶硝酸还原酶活性极显著高于其它3个处理,其它3个处理间NR活性差异不显著;但在35 ℃温度条件下,NR活性明显低于25 ℃遮光处理。说明25 ℃适宜温度条件下,短时间强光处理显著促进了烟株叶片NR活性上升,温度升至35 ℃时,NR活性有所下降,且光照强度对其影响不显著。

图4 不同光温处理对打顶烟株NR活性的影响
2.4.2 不同处理对叶片GS活性的影响 从图5可见,35 ℃强光处理的谷氨酰胺合成酶活性极显著高于其它3个处理,其它3个处理之间GS活性较为接近,差异不显著。说明GS活性相对比较稳定,仅在高温强光条件下表现为显著上升。

图5 不同光温处理对打顶烟株GS活性的影响
2.4.3 不同处理对叶片GOGAT活性的影响 从图6可见,在相同温度条件下,强光处理叶片的谷氨酸合成酶活性极显著高于遮光处理;强光条件下,35 ℃温度处理GOGAT活性极显著高于25 ℃温度处理,遮光条件下35 ℃温度处理GOGAT活性比25 ℃处理有所上升,但差异不显著。说明光照强度对GOGAT活性的影响较为显著,而仅在强光条件下温度变化对GOGAT活性的影响较为显著。

图6 不同光温处理对打顶烟株GOGAT活性的影响
2.5 不同处理对烟株叶片蛋白质水解氨基酸含量的影响
从表3可见,3-磷酸甘油酸衍生氨基酸主要有丝氨酸和甘氨酸两种。其中,在25 ℃条件下生长的烟叶中丝氨酸含量显著高于35 ℃条件下的;相同温度条件下,25 ℃时,强光条件下生长的烟叶丝氨酸含量显著高于遮光下的,35 ℃温度下表现相反。在25 ℃条件下甘氨酸含量未测出;在35 ℃条件下,强光条件下烟叶的甘氨碳含量高于遮光下的。

表3 不同光温条件对3-磷酸甘油酸衍生氨基酸含量的影响
从表4可见,在两种温度条件下,强光处理的苯丙氨酸、酪氨酸和色氨酸等磷酸烯醇式丙酮酸衍生氨基酸含量略高于遮光处理,但25 ℃条件下差异不显著,35 ℃条件下差异达到极显著水平。35 ℃处理烟株苯丙氨酸、酪氨酸和色氨酸含量极显著高于25 ℃处理。

表4 不同光温条件对磷酸烯醇式丙酮酸衍生氨基酸含量的影响
从表5可见,25 ℃条件下,与遮光处理相比,强光处理烟株的缬氨酸、亮氨酸和丙氨酸等丙酮酸衍生氨基酸含量有较明显增加,但差异不显著;35 ℃处理的烟株缬氨酸、亮氨酸和丙氨酸含量极显著高于25 ℃处理,且强光处理极显著高于遮光处理。

表5 不同光温条件对丙酮酸衍生氨基酸含量的影响
从表6可见,α-酮戊二酸衍生氨基酸包含脯氨酸、谷氨酰胺、谷氨酸、组氨酸和精氨酸等,在相同光强条件下,谷氨酰胺、谷氨酸、组氨酸和精氨酸等氨基酸含量随着温度的升高均表现为显著增加;在25 ℃温度条件下,谷氨酰胺、谷氨酸、组氨酸和精氨酸等氨基酸含量随光强增加而增加,其中谷氨酰胺和谷氨酸达到极显著水平;35 ℃温度条件下,谷氨酰胺、谷氨酸和精氨酸等氨基酸含量随光强增加而显著增加,组氨酸表现显著降低。在25 ℃温度条件下,遮光处理烟株叶片的脯氨酸含量显著高于不遮光处理;35 ℃温度处理表现相反,且均极显著低于25 ℃温度处理的。

表6 不同光温条件对α-酮戊二酸衍生氨基酸含量的影响
从表7可见,草酰乙酸衍生氨基酸包含苏氨酸、天冬氨酸、赖氨酸、甲硫氨酸、异亮氨酸和天冬酰胺等6种。25 ℃温度处理烟株叶片的苏氨酸、赖氨酸和异亮氨酸含量极显著低于35 ℃温度处理,且强光处理与遮光处理差异不显著;但35 ℃温度处理时,强光处理的烟叶苏氨酸、赖氨酸和异亮氨酸含量极显著高于遮光处理。不同处理烟株叶片的天冬氨酸含量差异不显著。25 ℃温度处理烟株叶片的甲硫氨酸含量随光强的增加而增加,但差异不显著;35 ℃温度遮光处理烟株叶片的甲硫氨酸含量最高,极显著高于其它3个处理,35 ℃温度强光处理烟株叶片的甲硫氨酸含量与25 ℃温度处理差异不显著。25 ℃温度处理烟株叶片的天冬酰胺含量随光强的增加表现为显著减少;35 ℃温度处理烟株叶片的天冬酰胺含量随光强的增加表现为极显著增加;35 ℃温度遮光处理的天冬氨酸含量高于25 ℃遮光处理,但差异不显著,35 ℃温度强光处理烟株叶片的天冬氨酸含量极显著高于其它3个处理。

表7 不同光温条件对草酰乙酸衍生氨基酸含量的影响
3 讨论
光合色素在光能的吸收、传递和转换过程中起着重要作用[1]。植物在生长期间,光合色素不断合成与分解,含量也不断发生变化。叶绿素是在系列酶促反应中合成的,合成的最适温度为26~30 ℃,高温下叶绿素合成的酶促反应受到抑制,并且降解量更大,降解、转化更充分,叶绿素含量降低[1,20]。叶绿素合成的最后阶段一般为需光反应,光照不足会使叶绿素合成受到影响,因此温度与光照是影响植物体内叶绿素含量的重要因素[1,16-21]。本文研究中,25 ℃短期强光处理下,Chl含量显著增加,说明适宜温度下,光照充足会促进叶绿素的合成,叶绿素含量增加,有利于光能的充分利用;而高温处理时,Chl和Chlx.c的含量均显著降低,说明高温导致烟株叶片中的总叶绿素和类胡萝卜素大量降解。35 ℃强光处理时,叶绿体色素含量均显著低于其他处理,在高温强光交互胁迫下,烟株叶绿体结构遭到破坏,叶绿素的合成受到抑制并加速分解,其含量显著降低,不利于烟株生长和品质的形成。
碳代谢是植物体内基本的代谢活动,直接影响烟株的生长发育、产量以及品质[22-23]。光合作用与碳代谢密切相关,两者呈正相关,光温条件影响碳代谢酶的活性[24-25]。在高等植物体内,参与糖代谢主要为SS及SPS两种酶:SS既控制蔗糖分解又可催化蔗糖合成,调节蔗糖分解与合成的可逆反应;SPS主要催化蔗糖的合成,两者活性的变化直接影响植物体内糖的合成[25-27]。淀粉合成酶是植物体内控制淀粉合成的关键酶[28]。当这3种酶的活性高时,植物新陈代谢快,光合作用能力强,有利于合成更多含碳物质,促进有机物的积累[29]。本文研究表明,在25 ℃条件下,短期强光处理SS、SPS及淀粉合成酶活性显著上升; 35 ℃强光条件下,3种酶活性则显著下降;在两种光条件下,升温均导致SPS及淀粉合成酶活性上升,说明在试验条件下,25 ℃短期强光及升温有利于促进蔗糖及淀粉的合成。从烟叶中蔗糖、还原糖、可溶性糖和淀粉等的含量看,相同温度下,光照增强,烟株的糖类物质及淀粉含量增加,这与SS、SPS及淀粉合成酶活性规律相吻合;相同光照条件下,升温促进糖类物质的积累,这与SS及SPS酶活性规律一致,说明适宜温度下短期强光及升温均有利于烟叶内糖类物质及淀粉的积累,这与任雷在甜瓜上的研究结果一致[10]。
氮代谢是植物体内重要生命活动,植物体内的蛋白质和氨基酸大部分是通过氮代谢合成的,氮代谢相关酶活性以及氮代谢产物含量也可作为衡量植物抗逆性的指标[14]。NR、GS及GOGAT是植物体内氨氮和硝氮同化的关键酶,NR催化NO3-还原成NO2-,而GS和GOGAT是同化铵态氮的关键酶[12]。氨基酸对烟叶的氮代谢和烟草香味物质的形成具有重要意义,氨基酸是合成蛋白质的重要原料,适量的氨基酸有利于提高烟叶的香气量和丰满度,氨基酸含量过低则烟气寡淡无味且丰满度差,过高则燃吸过程中会产生氨气,导致烟气刺激辛辣,不利于烟叶品质的形成[1,30-31]。本试验中,适宜温度下短时间强光处理显著促进了烟株叶片中NR活性上升,其他处理差异不大;高温强光处理的GS、GOGAT活性极显著高于其它处理,与此同时大多数水解氨基酸含量在高温强光条件下显著高于其他3个处理,这可能是由于水溶蛋白及游离氨基酸总量的增加,其原因可能是高温强光胁迫下,氮代谢相关酶活性提高,诱导逆境蛋白的合成,胁迫的增加进一步导致蛋白质大量水解,致使游离氨基酸总量增加,增加的游离氨基酸可能在调节渗透压、维持细胞水势及细胞结构、消除物质毒害及储存氮素方面起着重要作用,氮同化的加强需要大量的碳架,叶片中的碳流会从合成糖类的方向转为合成氨基酸[32-33]。高温强光环境显著提高烟株氮代谢能力:一方面,由于高温强光条件下氮代谢相关酶活性增加[34-35];另一方面,为了减少环境胁迫对细胞膜的伤害,很多有害物质也会转化成含氮物质,提高植物氮代谢能力[7,13]。氮同化作用增强,含氮物质增加,不利于中上部烟叶品质的形成。
福建种植春烟,在烤烟生长后期常遇上高温和强光天气。从本研究看,高温和强光导致烟株碳的合成代谢和氮的同化代谢均加强。更多的糖类物质积累可能导致叶片增厚甚至粗筋爆叶;而氮代谢的加强可能导致烟叶中碱性物质的增加,影响烟气平衡,最终均会导致烟叶感官评吸质量的下降。因此,适时早栽和适时成熟采收对福建的烤烟生产极为重要;另外,本研究高温强光处理时间仅为48 h,进一步延长高温和强光的处理时间会对烟株产生哪些不利影响还有待进一步研究。
4 结论
在高温强光胁迫下,烟株叶片的叶绿体结构遭到破坏,叶绿素合成受到抑制,分解加快,含量显著下降,烟株光合作用过程受到抑制;烟株体内代谢过程受到影响,SS酶活性显著下降,蔗糖、还原糖、果糖、可溶性糖含量均高于其余3个处理,说明高温强光处理下,SS催化蔗糖合成与分解的反应过程向合成蔗糖方向进行,烟株糖类物质积累增加。在高温强光胁迫下,NR酶活性低于25.0 ℃强光处理,与其余两组处理差别不大,GS、GOGAT酶活性极显著高于其它处理,与此同时大多数游离氨基酸含量显著高于其他3个处理,说明高温强光环境显著提高烟株的代谢能力,含氮物质合成增多,不利于烟叶品质的形成。
