亚东鲑幼鱼饲料中适宜淀粉种类与水平的研究
2022-01-20党江雨高擘为李小勤冷向军
党江雨 高擘为 徐 禛 杨 航 李小勤 冷向军
(1. 上海海洋大学水产科学国家级教学示范中心, 上海 201306; 2. 上海海洋大学农业农村部鱼类营养与环境生态研究中心,上海 201306; 3. 水产动物遗传育种中心上海市协同创新中心, 上海 201306)
糖类, 即碳水化合物, 是食物中最廉价的能量来源, 在很多粮食作物中含量丰富[1]。水生动物为变温动物, 相较于陆生动物, 体内代谢用于维持体温的比例低, 并不需要太多碳水化合物, 故其需求量较低。研究表明鱼类饲料中添加一定量的碳水化合物, 不仅可以减少饲料中蛋白质和脂肪的添加量, 节省饲料成本, 还可以提高鱼类的生长性能[2,3]。淀粉因其来源广泛, 黏合性好[4], 是水产饲料中使用最多的糖源。但不同来源的淀粉, 分子结构不同,其支链和直链淀粉的比例存在差异[5], 对利用率产生较大影响[6—8], 进而影响水产动物的生长性能。关于鱼类对不同来源淀粉利用的相关研究在虹鳟(Oncorhynchus mykiss)[9,10]、大西洋鲑(Salmo salar)[11]、大口黑鲈(Micropterus salmoides)[12]和达氏鲟(Acipenser dabryanus)[13]等鱼类中已有报道。由于鱼类对糖的耐受能力有限, 饲料中过多的糖会对鱼类造成生长减缓[14]、肝脏损伤[15]、血糖升高[16]和免疫损伤[17]等负面影响。
亚东鲑, 与褐鳟(Salmo trutta)同物异名, 属于鲑形目(Salmoniformes)鲑科(Salmonidae)鲑亚科(Salmoninae), 原产自欧洲、非洲北部和西亚一些地区[18]。现已分布于30多个国家和地区[19], 国内仅天然分布于西藏海拔3000 m的亚东河中[20], 为外来引进物种。因为其在亚东河已形成自然种群, 故命名为亚东鲑, 是一种肉质鲜美, 营养丰富的特色养殖鱼类。目前, 国外关于褐鳟糖类营养研究的报道较少,Viaplana-Marín等[21]发现, 低蛋白质/高碳水化合物饲料组较高蛋白质/低碳水化合物饲料组褐鳟的蛋白质合成速率没有显著差异, 但蛋白质降解速率增加, 使得蛋白质沉积率降低; 另外, 也有一些关于褐鳟饲料脂肪来源[22—24]及脂肪替代[25,26]对其造成的影响报道。国内关于褐鳟的营养需求研究仅见褐鳟幼鱼[27]和成鱼[28]的蛋白质和脂肪需求的报道, 尚未见到有关碳水化合物需求的报道。
本实验以亚东鲑为研究对象, 分别以玉米淀粉和木薯淀粉为糖源, 设置不同的添加水平, 考察对生长、体组成、消化酶活性及肝脏生化指标和肝脏组织学的影响, 确定适宜淀粉种类与水平, 为开发高效亚东鲑饲料提供理论依据, 推动西藏当地亚东鲑养殖业的进一步发展。
1 材料与方法
1.1 实验设计与实验饲料
实验采用2×3双因素设计, 分别以玉米淀粉和木薯淀粉为糖源, 设置3种淀粉水平(5%、10%和15%), 共6种等氮等脂饲料。实验饲料配方含量见表 1。使用膨润土和纤维素来平衡各组饲料组成。饲料原料购于浙江粤海饲料有限公司。各主要饲料原料被粉碎过60目筛后, 按配方比例称取进行混合, 用单螺杆挤压机(SLP-45,中国水产科学研究院渔业机械仪器研究所)制成粒径2 mm的沉性颗粒饲料, 自然风干, 密封储藏于阴凉干燥处备用。
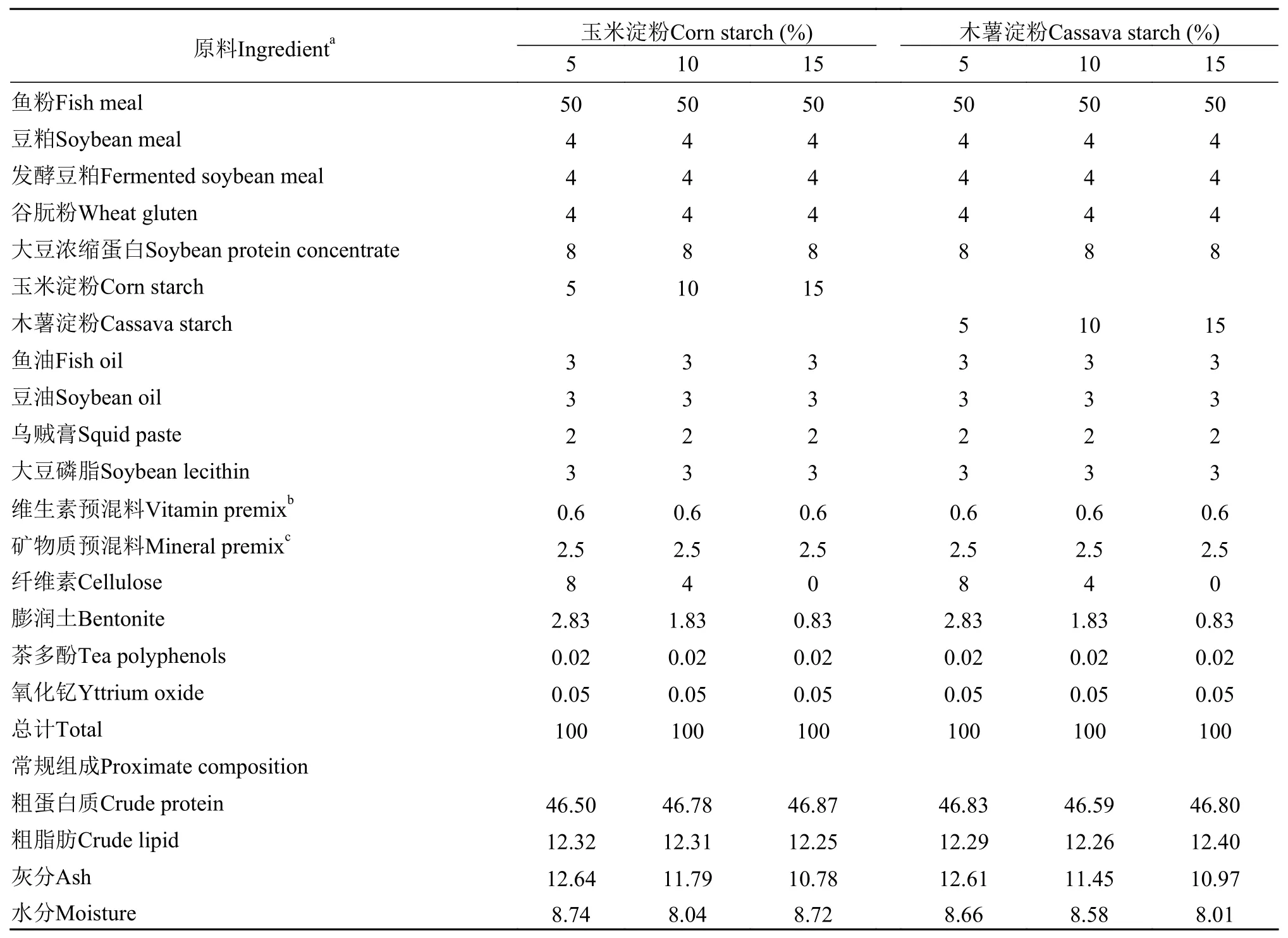
表 1 实验饲料配方组成及营养成分含量Tab. 1 Ingredients and proximate composition of experimental diets (%)
1.2 实验鱼和饲养管理
养殖实验在西藏亚东县春丕村亚东鲑繁育基地进行, 实验鱼由繁育基地提供。实验鱼用商品饲料(INICIO Plus, 粗蛋白58%, 粗脂肪15%)暂养驯化2周后, 选取720尾体格健壮、规格均一的亚东鲑(0.50±0.03) g, 随机分配到18个塑料框中(长0.55 m,宽0.30 m, 高0.20 m, 水体体积0.0165 m3), 每个实验组3个重复, 每个重复40尾鱼。养殖模式为流水养殖, 水流量为12 L/min。养殖期间, 每天在(10:00、14:00和18.00)3个时间点进行投喂, 每天投喂量约为鱼体重的2.0%—3.0%, 同时随着水温和摄食情况适当调整, 各框保持一致的投饲量, 每次投喂以亚东鲑不浮上水面摄食和水面下不抢食为宜。养殖期间每天检测流水水质, 其基本指标如下: 溶氧>6 mg/L, 水温7.5—12.5℃, pH 7.5—8.0, 氨氮<0.1 mg/L,亚硝酸盐<0.1 mg/L, 并每周采用虹吸法吸污2次。养殖实验持续84d。
1.3 样品采集
养殖实验结束后, 将亚东鲑饥饿24h, 统计每框亚东鲑尾数并称重, 计算增重率(WGR)、饲料系数(FCR)和成活率。每框随机取3尾亚东鲑幼鱼, 测量其体重和体长, 然后解剖取内脏称重, 计算肥满度(CF)和脏体指数(HSI)。另取肝脏和肠道(全肠),-20℃冷冻保存, 用于肝脏生化指标和胃、肠消化酶活性测定。同时从每个框中随机取6尾, 用于全鱼体成分分析, 并计算蛋白质和脂肪沉积率。另取6尾鱼的肝脏, 测定肝糖原含量。再取3尾鱼肝脏置于波恩固定液中固定, 用于肝脏组织学观察。
1.4 测定指标与方法
生长性能与形体指标成活率(%)=100×终尾数(尾)/初尾数(尾);
增重率(%)=100×[末体重(g)-初体重(g)]/初体重(g);
饲料系数=采食量(g)/[末体重(g)-初体重(g)];
脏体比=100×内脏重(g)/体重(g);
肥满度(g/cm3)=100×体重(g)/体长(cm)3;
蛋白质沉积率(%)=100×鱼体蛋白质贮积量/摄入饲料的蛋白质总量;
脂肪沉积率(%)=100×鱼体脂肪贮积量/摄入饲料的脂肪总量。
全鱼与饲料组成全鱼与饲料的水分、粗蛋白含量测定分别采用105℃常压干燥法和凯氏定氮法(2300自动凯氏定氮仪, FOSS, 瑞典), 粗脂肪含量测定采用氯仿-甲醇抽提法, 粗灰分测定采用550℃高温灼烧法。
胃肠道消化酶测定取胃、肠道(全肠)样本于4℃解冻, 稀释匀浆, 4℃离心(3000 r/min, 10min), 保留上清液, 采用试剂盒测定消化酶活性。胃蛋白酶和淀粉酶活性采用南京建成生物工程研究所提供的试剂盒测定。肠道蛋白酶活性采用福林酚法测定。
肝脏生化指标及肝糖原含量测定取肝脏样本于4℃解冻, 稀释匀浆, 4℃离心(3000 r/min,10min), 保留上清液, 采用南京建成生物工程研究所试剂盒测定肝脏谷草转氨酶(GOT)、谷丙转氨酶(GPT)、总胆固醇(TCHO)和甘油三酯(TG)活性。
将肝脏样本于4℃解冻后, 称重, 将肝脏与碱液以1∶3的体积混合, 在沸水浴中加热20 min, 得到水解液, 采用南京建成生物工程研究所试剂盒测定肝糖原含量。
肝脏组织切片石蜡切片HE染色: 将亚东鲑肝脏样品从固定液中取出, 用不同浓度的酒精、酒精和二甲苯混合液、二甲苯逐级脱水, 之后进行浸蜡、包埋、切片、染色等工作。染色后装片, 待干燥后在电子显微镜下观察其组织形态。
1.5 数据处理
采用SPSS25.0软件进行单因子方差分析和双因素方差分析, 结果用“平均值±标准误”表示, 其中差异显著者进行Duncan多重比较, 差异显著水平为P<0.05。
2 结果
2.1 生长性能和形体指标
在养殖过程中, 各组存活率都在96%以上(表 2)。随着饲料中玉米淀粉和木薯淀粉水平的提高, 增重率先上升后下降, 饲料系数则先下降后上升(P<0.05),并在10%水平组达到最值, 其中10%木薯淀粉组增重率最高(518.81%), 饲料系数最低(1.32), 15%淀粉组的增重率显著低于10%和5%组(P<0.05)。在同一淀粉水平下, 2种淀粉组的增重率和饲料系数没有显著差异(P>0.05)。饲料淀粉水平对增重率、饲料系数存在显著影响(P<0.05), 淀粉种类及其交互作用没有显著影响(P>0.05), 而饲料淀粉种类和水平对成活率、脏体比和肥满度均无显著影响(P>0.05)。
2.2 全鱼组成和营养物质沉积率
各组在全鱼水分、粗蛋白质、粗脂肪和粗灰分含量上均无显著差异(P>0.05)。饲料淀粉水平对蛋白质沉积率存在显著影响(P<0.05), 淀粉种类和水平对脂肪沉积率存在显著影响(P<0.05), 但其二者交互作用对其影响不显著(P>0.05)。15%水平组蛋白质沉积率显著低于其他水平(P<0.05); 而脂肪沉积率则随着淀粉水平的升高先上升后下降, 10%木薯淀粉组显著高于玉米淀粉组(P<0.05; 表 3)。
2.3 胃肠道消化酶
在表 4中, 淀粉种类和水平及其交互作用对胃蛋白酶和胃淀粉酶活性没有显著影响(P>0.05), 但淀粉水平显著影响肠淀粉酶和肠蛋白酶活性(P<0.05)。随着饲料中淀粉水平的提高, 肠淀粉酶和肠蛋白酶呈现上升趋势, 5%淀粉水平组的肠淀粉酶和肠蛋白酶均显著低于其他水平(P<0.05)。
2.4 肝脏生化指标及肝糖原
由表 5可见, 淀粉种类和水平及其交互作用对肝脏GPT、GOT活性和TCHO、TG含量均没有显著影响(P>0.05)。淀粉水平对肝糖原含量存在显著影响(P<0.05), 随着饲料中淀粉水平的提高, 肝糖原含量呈上升趋势, 15%淀粉水平组显著高于其他水平(P<0.05)。
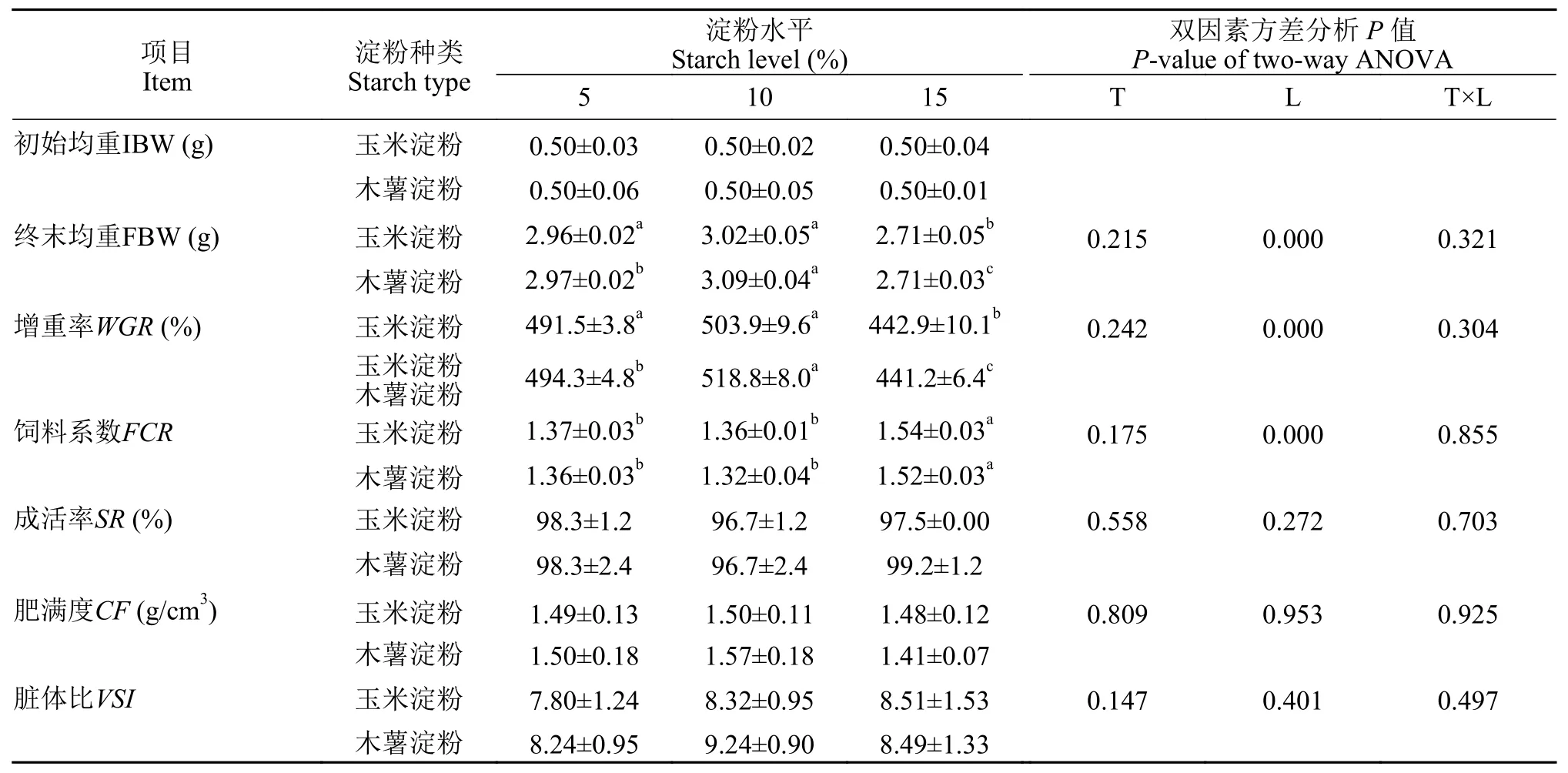
表 2 饲料淀粉种类和水平对亚东鲑幼鱼生长性能和形体指标的影响Tab. 2 Effects of dietary starch on growth and morphological indices of brown trout juvenile
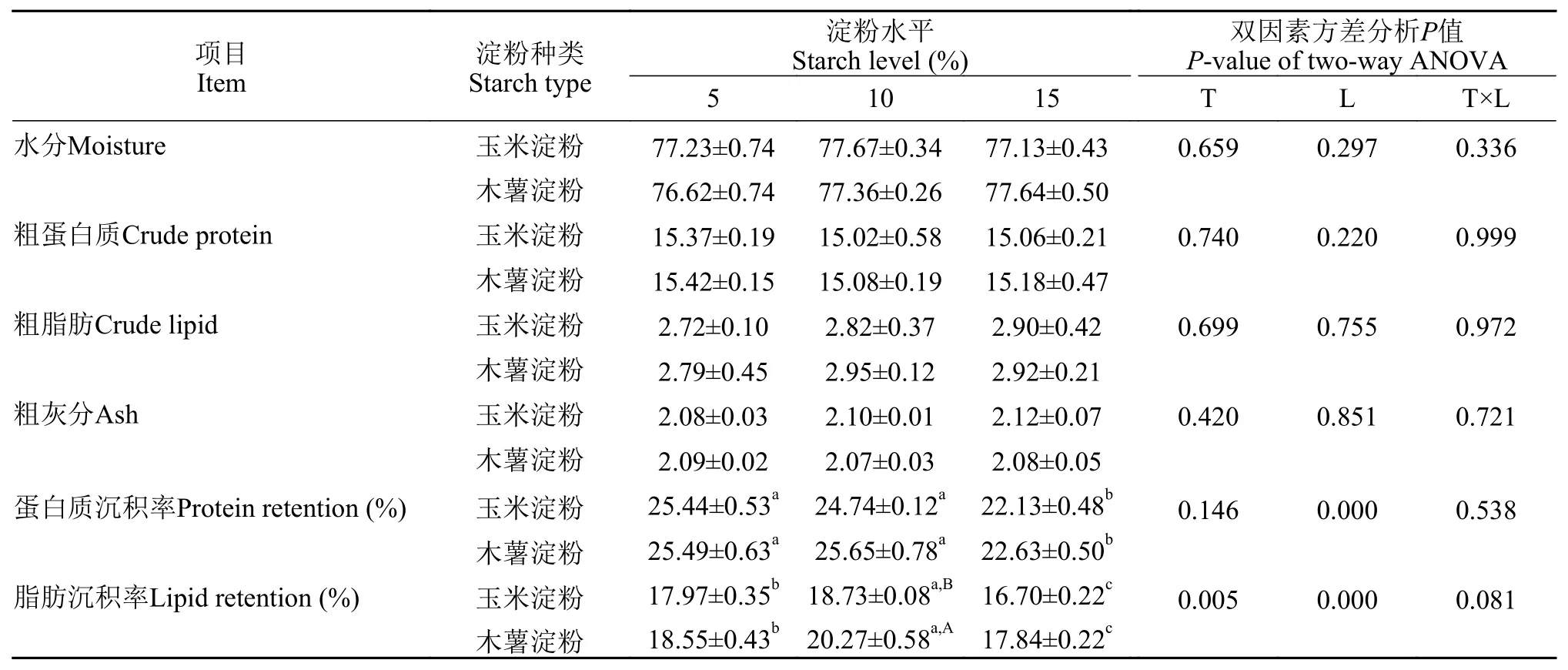
表 3 饲料淀粉种类和水平对亚东鲑幼鱼全鱼组成和营养物质沉积率的影响Tab. 3 Effects of dietary starch on body composition and nutrient retention of brown trout juvenile (%)
2.5 肝脏组织学
如图 1所示, 随着木薯淀粉和玉米淀粉水平的提高, 肝细胞开始肿胀, 出现细胞核位移和空泡化现象, 其中以15%淀粉组较为明显。
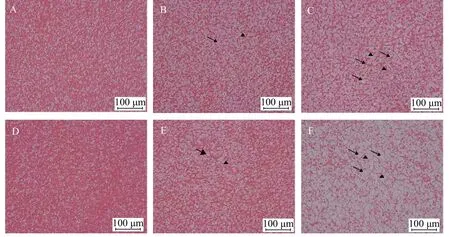
图 1 亚东鲑幼鱼肝脏组织切片Fig. 1 Tissue slices of brown trout juvenile liver (200×)
3 讨论
3.1 饲料淀粉种类和水平对亚东鲑幼鱼生长性能的影响
尽管鱼类对碳水化合物没有特别的需求, 但一定量的碳水化合物当做能量被添加进鱼类饲料中时, 可以减少蛋白质和脂肪作为能量物质的分解,从而使更多的蛋白质和脂肪用于鱼体的生长, 提高饲料利用的效率[1,29]。在大西洋鲑(Salmo salar)[30]和鲈(Lateolabrax japonicus)[31]中的研究表明, 饲料中淀粉水平为9%和12%时, 鱼体生长性能得到了显著提升。在本实验中, 适量的淀粉添加水平也改善了亚东鲑幼鱼的生长性能, 根据增重率和饲料系数,亚东鲑幼鱼饲料中淀粉的适宜水平为10%, 与虹鳟(Oncorhynchus mykiss)[32]对碳水化合物的需要量(12%)基本一致。而尼罗罗非鱼(Oreochromis niloticus×O. aureus)[33]和草鱼(Ctenopharyngodon idellus)[34]饲料中碳水化合物的最适添加量可达到22%和30%以上, 远远高于亚东鲑, 这是因为亚东鲑属于肉食性冷水鱼类, 较杂食性和草食性鱼类利用糖的能力差[2], 对碳水化合物的需要量相对较低。
此外, 淀粉的来源也会对鱼类生长产生影响。淀粉由直链淀粉和支链淀粉两种结构淀粉组成, 支链淀粉比直链淀粉更容易被酶水解, 所以两者的比例会影响淀粉的消化利用, 从而影响水产动物生长[11]。Li等[35]和刘子科等[12]的研究表明, 在大口黑鲈摄入直链淀粉比例高的淀粉源后, 其直链淀粉在加工过程中形成难以被消化的抗性淀粉, 使实验鱼的餐后血糖水平较低, 从而表现出对直链淀粉比例高的淀粉源有较好的利用效果, 而虹鳟对支链淀粉比例高的淀粉源的利用程度更高[36]。所以, 不同鱼种对不同比例的直链淀粉和支链淀粉的淀粉源利用程度不同。本实验中10%木薯淀粉组的增重率高于玉米淀粉, 但差异不显著, 表明亚东鲑可能对支链淀粉比例高的淀粉源的利用程度高, 这有待进一步研究。
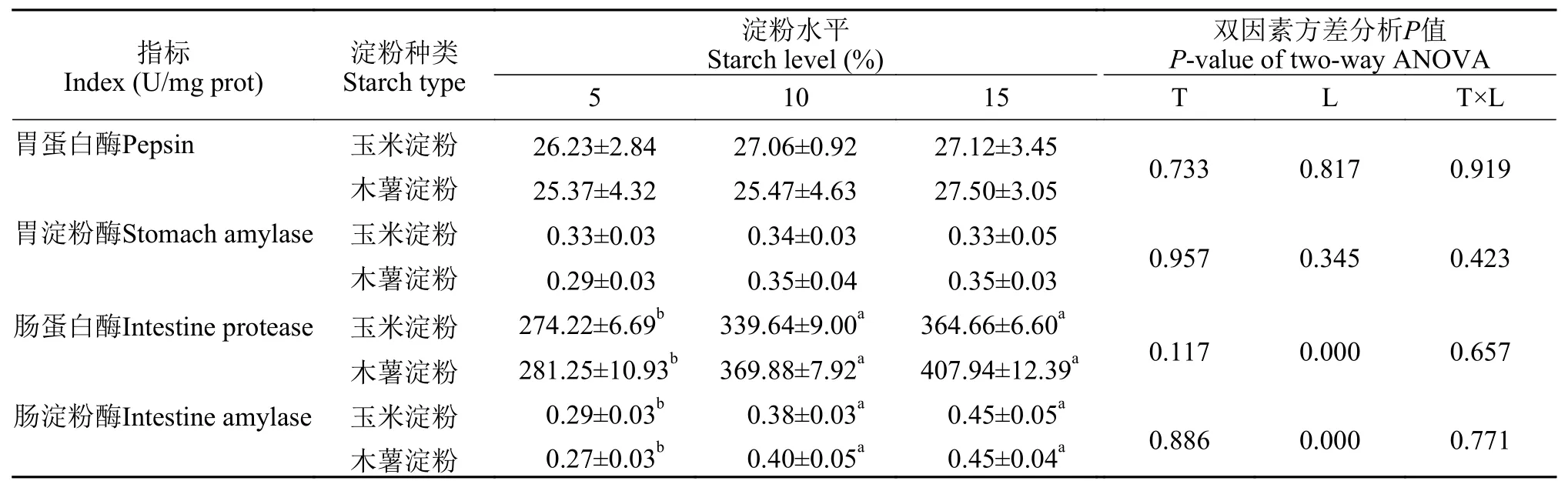
表 4 饲料淀粉种类和水平对亚东鲑幼鱼胃肠道消化酶的影响Tab. 4 Effects of dietary starch on digestive enzyme activities of brown trout juvenile
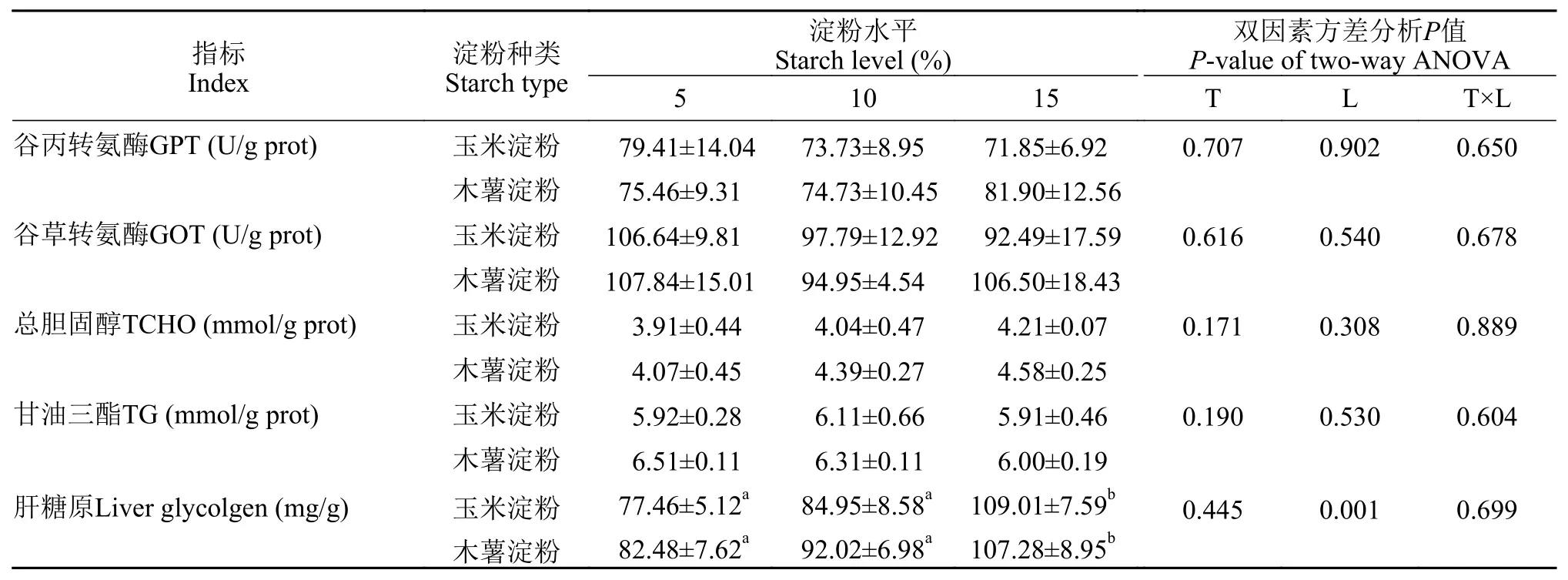
表 5 饲料淀粉种类和水平对亚东鲑幼鱼肝脏生化指标及肝糖原的影响Tab. 5 Effects of dietary starch on liver biochemical indices and liver glycogen of brown trout juvenile
3.2 饲料淀粉种类和水平对亚东鲑幼鱼全鱼组成的影响
在大黄鱼[37]中的研究显示, 随着淀粉水平的提高, 全鱼脂肪含量上升。但在赤点石斑鱼(Epinephelus akaara)[38]和黄鳍鲷 (Sparus latus)[39]的研究中,随着碳水化合物水平的提高, 鱼体粗脂肪含量下降。本实验中亚东鲑全鱼组成不受淀粉种类与水平的影响, 与在大菱鲆(Scophthalmus maximusLinnaeus)[40]上的报道类似。这可能是因为不同种鱼类对淀粉利用能力的不同导致的。
虽然本实验中全鱼组成没有显著差异, 但淀粉水平显著影响着蛋白质沉积率和脂肪沉积率: 15%淀粉组的蛋白质沉积率显著低于其他两个水平组,10%淀粉水平组的脂肪沉积率显著高于其他2个水平, 这主要是由增重的差异所决定的。此外, 木薯淀粉10%组的脂肪沉积率显著高于玉米淀粉10%组, 可能亚东鲑幼鱼能更好地利用支链淀粉。
3.3 饲料淀粉种类和水平对亚东鲑幼鱼胃肠道消化酶的影响
消化酶的活性可在一定程度上衡量鱼类对饲料营养成分消化、吸收和利用程度。在本实验饲料中, 淀粉种类和水平对亚东鲑胃蛋白酶及胃淀粉酶活性无显著影响, 而肠蛋白酶及肠淀粉酶随着淀粉水平的提高而增加。在军曹鱼(Rachycentron canadum)[4]、黑鲷(Acanthopagrus schlegelii)[41]和洛氏鱥(Rhynchocypris lagowskiiDybowski)[42]中也有过类似的报道。淀粉酶活性增加, 可能是鱼类为了适应高碳水化合物而做出的适应性调整[43]。
3.4 饲料淀粉种类和水平对亚东鲑幼鱼肝脏生化指标影响
GOT和GPT是反映肝脏功能的重要指标, 甘油三酯和胆固醇则反映鱼类机体脂肪代谢情况。在大口黑鲈[44]的研究中, 21.74%碳水化合物水平组血清GOT和GPT活性显著高于5.93%、8.71%和11.96%碳水化合物水平组, 在翘嘴红鲌(Erythroculter ilishaeformisBleeker)[45]中也有类似报道; 高水平的碳水化合物也导致了翘嘴红鲌[46]和鲤(Cyprinuscarpio)[47]血清甘油三酯和胆固醇含量显著提高。在本实验中, 由于鱼体规格小, 无法采血测定血液生理生化指标, 故只测了肝脏部分生化指标,结果表明, 淀粉种类及水平对甘油三酯和胆固醇含量及转氨酶活性并无显著影响, 说明亚东鲑转化糖类为脂肪的能力较弱, 并没有导致脂肪在肝脏的积累; 肝脏GOT和GPT活性在各组没有显著差异, 但由于没有检测其在血液中的变化, 尚不足以据此评判肝脏的健康状况, 需结合肝脏组织学等加以综合评判。
3.5 饲料淀粉种类和水平对亚东鲑幼鱼肝脏组织学影响
在鱼类摄入淀粉后, 首先会被α-淀粉酶水解为麦芽糖, 进一步水解释放出葡萄糖, 葡萄糖在生糖作用下合成糖原储存起来。所以摄入含高水平糖类的饲料会造成肉食性鱼类肝脏糖原升高, 不利于鱼类的正常代谢, 并导致生长的下降[48]。在本实验中, 肝糖原含量随着淀粉水平的增加显著提高, 与Lygren等[49]、刘浩等[50]和Moreira等[51]在大西洋鲑、斜带石斑鱼(Epinephelus coioides)和欧洲狼鲈(Dicentrarchus labrax)中的报道一致。
Pereira等[52]在虹鳟上的研究表明, 肝细胞同时存在糖原合成和糖原分解的过程, 鱼类长期摄入高糖饲料, 导致肝糖原的积累, 会使鱼类产生营养胁迫。异育银鲫(Carassiusauratus gibelio)[53]和吉富罗非鱼(Oreochromis niloticus)[54]在摄入高糖饲料后,肝脏细胞脂滴和糖原颗粒大量出现, 说明鱼类无法及时将糖分解代谢, 对肝脏细胞造成了一定程度损伤。在本实验中, 随着饲料淀粉水平增加, 肝脏细胞空泡化现象逐渐加重, 并且存在细胞核移位等现象, 这在松浦镜鲤(Cyprinus carpio specularis)[55]、大口黑鲈[56]和南方鲇(Silurus meridionalis)[57]的研究结果中也有类似现象出现, 这可能是由于肝脏负担过重, 鱼类对糖代谢负担过大所致。
4 结论
在本实验条件下, 以生长性能、饲料利用、胃肠道消化酶、肝脏生化指标和组织学等指标为依据, 亚东鲑幼鱼饲料中淀粉的适宜添加水平为10%,且木薯淀粉优于玉米淀粉。
