miR-140-5p通过调节自噬影响结直肠癌细胞对奥沙利铂的敏感性
2022-01-03延飞飞李宏武
延飞飞 李宏武
中国医科大学附属第四医院普外科(沈阳110032)
结直肠癌(colorectal cancer,CRC)是全球第三大癌症致死的主要原因之一[1]。奥沙利铂是治疗CRC 最常用的药物之一[2]。通过诱导链内加合物的形成,奥沙利铂可以抑制细胞周期的进展,促进增殖细胞的死亡[3]。然而,新生和获得性奥沙利铂耐药显著降低了该药物治疗CRC 的疗效[4]。因此,深入研究CRC 耐药的分子机制,提高临床治疗效果具有重要意义。
微小核糖核酸(microRNAs,miRNAs)是小的、非编码的单链RNA 分子[5]。据报道,miRNAs 的失调有助于多种癌症的发生,包括CRC[6]。CRC 患者血浆外泌体中miR-23b的表达水平较正常健康人明显降低,其低表达与肿瘤分化程度、浸润深度、淋巴结转移、TNM 分期、脉管浸润、神经侵犯显著相关[7]。此外,许多miRNA 已经被确定通过调控靶基因参与CRC 的化疗耐药[8]。先前的研究表明CRC患者血清miR-140-5p较对照组明显下调,并与TNM 分期和淋巴结转移等临床病理参数显著相关[9]。KOUSTAS 等[10]研究表明miR-140-5p 的下调与CRC 的晚期和较低的总生存率显著相关。miR-140-5p通过抑制细胞增殖、迁移和侵袭而起到抑癌作用。因此,miR-140-5p可能在CRC中起到抑癌作用,以作为一种新的预后标志物和治疗靶点。然而,miR-140-5p在CRC耐药中的研究鲜有报道。
细胞自噬通过降解细胞器来使其适应缺氧或营养缺乏等肿瘤微环境,研究表明自噬在CRC 的进展中起着至关重要的作用[11]。因此,探讨CRC中自噬调节机制对于新型治疗是必不可少。当前研究表明自噬在参与CRC 细胞耐药中发挥重要作用[12]。已有研究发现,多个miRNA在CRC自噬的调节中起着重要作用,这种调节可以影响肿瘤的进展。miR-140-5p 在CRC 干细胞中的异位表达导致自噬破坏,抑制肿瘤干细胞生长和球形形成[13]。但是目前关于miR-140-5p 调控自噬在CRC 细胞奥沙利铂耐药中的相关作用尚未明确。
本文旨在研究miR-140-5p 对CRC 细胞奥沙利铂的敏感性,并探索miR-140-5p 是否通过调节自噬促进CRC 细胞奥沙利铂敏感性的影响,为临床诊治提供一定的策略。
1 材料与方法
1.1 组织来源和细胞培养收集中国医科大学附属第四医院2018年12月至2019年12月之间手术切除的30 例CRC 组织标本。本研究取得医院伦理委员会批准,并经患者签署知情同意书后进行。3种人CRC细胞系(HT-29,RKO,HCT-116)和1种人回盲肠腺癌细胞系(HCT-8)及正常结直肠上皮细胞FHC 均购置于中国生命科学院上海细胞研究所(中国,上海)。细胞培养用RPMI-1640 添加10%胎牛血清(Hyclone 公司,美国),在5%CO2、37 ℃培养箱中常规培养。
1.2 RNA 提取及qPCR 检测使用Trizol 试剂(Invitrogen 公司,美国)用于裂解细胞并收集RNA,后加入1/5 体积氯仿离心,取上清加入等体积异丙醇过夜。使用Power SYBR Green(TaKaRa 公司,大连)做real-Time PCR 分析。miR-140-5p 上游引物:5′-TCATGATGGAATTGGAGCCTT-3′,下游引物:5′-GCAGGGTCCGAGGTATTC-3′;β-Actin 上游引物:5′-TCAAGATCATTGCTCCTCCTG-3′,下游引物:5′-CTG-CTTGCTGATCCACATCTG-3′。实时PCR 和数据收集基于ABI 7500 平台。miR-140-5p 相对于β-Actin 的表达采用2-ΔΔCt法计算并标准化。
1.3 质粒的构建及细胞转染miR-140-5p 过表达质粒购自上海吉凯基因公司,选用miR-NC 空白载体作为阴性对照。将CRC 细胞株HCT 116 按每孔5 × 105个种于6 孔板中,待细胞达到80%融合时,用miR-140-5p mimics 以及相应的阴性对照miR-NC 通过Lipofectamine 3000(Invitrogen 公司,上海)转染细胞,48 h 后收获细胞用于RT-qPCR 检测,72 h 后用于western blot 检测。
1.4 CCK-8 法测定细胞增殖能力取对数生长期细胞接种于96 孔板中(5 × 103个/孔),各组细胞经相应处理后继续培养24 h,分别加入10%CCK-8培养基,置于培养箱中孵育2 h。最后,用酶标仪测定450 nm 波长下各样本的OD值。
1.5 Western blot 法检测蛋白表达收集不同处理因素下的细胞,用预冷却的磷酸盐缓冲液(phosphate buffered saline,PBS)液将其冲洗两次。然后快速加入细胞裂解缓冲液以使细胞裂解。收集蛋白质后,用标准曲线法检测蛋白质浓度,定完蛋白浓度后各样品加入5×上样缓冲液煮沸5 min。配好胶后加入相同量的蛋白质,之后进行十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate polyacrylamide gel electrophoresis,SDS-PAGE)。随后,在50 V 恒定电压的情况下转膜120 min,将蛋白质转移到聚偏二氟乙烯膜(polyvinylidene fluoride,PVDF)膜上,然后用5%干燥的脱脂乳密封PVDF 膜2 h,用特定抗体孵育一抗、二抗,进而检测特定蛋白(LC3、β-actin 抗体购置于Cell Signaling Technology 公司)的表达,β-Actin 作为对照。
1.6 免疫荧光实验检测自噬小体将爬片放入6 孔板中,将稳定表达的HCT 116 细胞接种于6 孔板中,待细胞贴壁且生长到80%左右,分别转染miR-140-5p mimics 以及miR-NC,于培养箱培养24 h 之后每孔加入预冷的4%多聚甲醛1 mL 室温固定20 min,漂洗三次后每孔加入0.5% TritonX-100 室温下孵育10 min。漂洗三次后加入2%BSA 室温下封闭30 min。之后加入LC3 一抗,孵育过夜后避光加入荧光二抗(绿色标记),滴加DAPI 复染核(蓝色标记)。以miR-NC作为阴性对照,在荧光显微镜下检测各组CRC细胞内LC3荧光斑点的分布与数量。
1.7 克隆形成实验取对数生长的CRC 细胞,用0.25%胰蛋白消化并吹打成单个细胞,把细胞悬浮在10%胎牛血清的RPMI-1640 培养液中备用。以每孔100 个细胞密度接种6 孔板中,待24 h 细胞贴壁后分别给予不同的处理方式,置37 ℃、5% CO2的湿润的环境下培养1 ~2 周,经常观察,当在显微镜下出现团状克隆时,终止培养,弃去上清,用PBS 小心浸洗2 次,每孔加入4%多聚甲醛固定15 min,之后用结晶紫染色10 min,然后用流水缓慢洗去染色液,干燥后进行图片采集。
1.8 统计分析SPSS 软件(SPSS 22.0)用于进行结果统计。Image J 进行蛋白和荧光定量分析,GraphPad Prism7.0 和Photoshop 和用于图像处理。实验数据定量资料以均数±标准差表示,选用独立样本t检验进行统计学分析。P<0.05 则认为差异有统计学意义。
2.2 过表达miR-140-5p促进CRC细胞对奥沙利铂的敏感性为检测过表达miR-140-5p对CRC细胞对奥沙利铂的敏感性,用miR-NC和miR-140-5p mimics质粒转染CRC细胞系HCT-116,构建miR-140-5p过表达细胞。RT-qPCR 用于分析miR-140-5p 表达。miR-NC 的表达标准化为1,miR-140-5p mimics转染组的miR-140-5p 相对高表达(P=0.000 2,图2A)。之后利用CCK-8 实验检测miR-NC 和miR-140-5p mimics 转染组在不同浓度奥沙利铂处理24 h 下的细胞增殖情况。结果显示,在5 μmol/mL[(78.767±3.365)vs.(60.800±4.300)]、10 μmol/mL[(63.900±4.011)vs.(37.600 ± 5.251)]、15 μmol/mL[(52.433±6.104)vs.(28.567±2.909)]、20 μmol/mL[(50.800±3.351)vs.(21.200±4.518)]浓度的奥沙利铂处理24 h 下,过表达miR-140-5p 促进CRC 细胞系HCT-116对奥沙利铂的敏感性(P<0.000 1,图2B)。
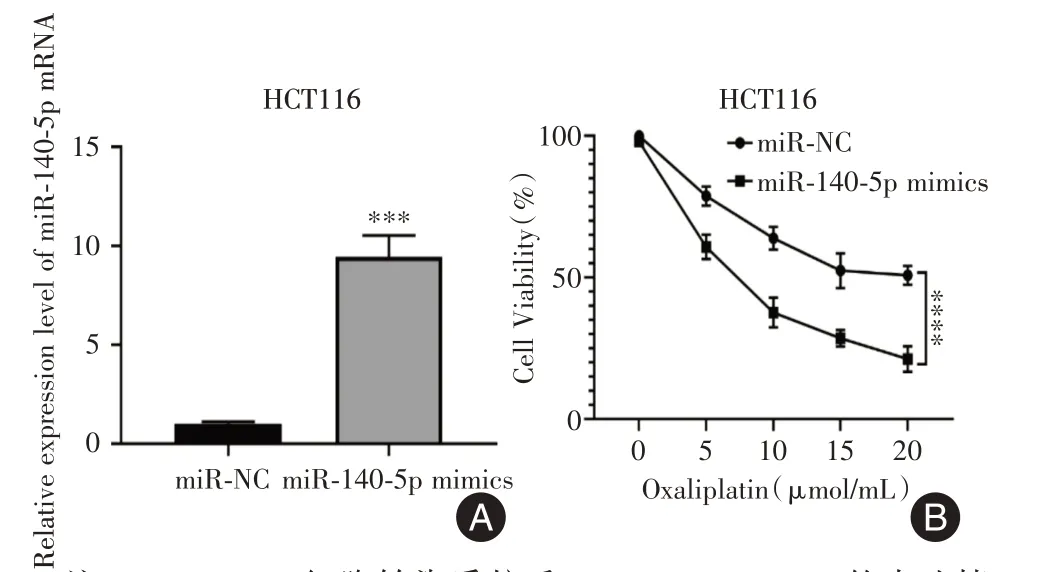
图2 不同浓度奥沙利铂处理下,过表达miR-140-5p 对结肠癌细胞活性的影响Fig.2 Effect of overexpression of miR-140-5p on the cell viability of CRC cells treated with different concentrations of oxaliplatin
2 结果
2.1 miR-140-5p 在CRC 组织中及细胞系中的表达RT-qPCR 用于检测30 例CRC 组织及相应癌旁组织样本中miR-140-5p 表达,癌旁组织表达标准化为1,CRC 组织中的miR-140-5p 的平均表达水平低于癌旁组织(0.343±0.107),差异有统计学意义(P=0.0015,图1A)。RT-qPCR 用于分析细胞中miR-140-5p 的表达,正常结肠上皮细胞(FHC)的表达标准化为1,结果表明在HCT-8、HT-29、RKO、HCT-116 中miR-140-5p 的表达低于FHC 细胞,其中HCT-116 细胞系差异有统计学意义(P<0.05),用于后续实验(图1B)。
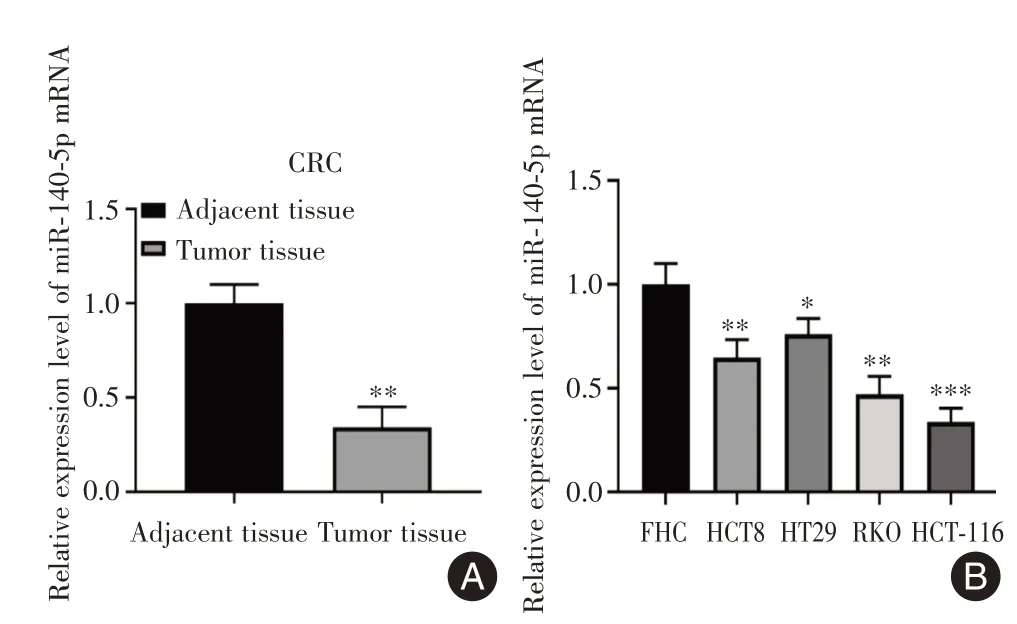
图1 miR-140-5p 在CRC 组织中及细胞系中的表达Fig.1 Expression of miR-140-5p in CRC tissues and cell lines
2.3 过表达miR-140-5p 抑制CRC 细胞的自噬由于自噬在CRC 的发生发展中起到关键的调控因素,检测miR-140-5p 是否能调节CRC 细胞的自噬。Western blot 检测LC3 蛋白的表达,与miR-NC相比,转染miR-140-5p mimics 后LC3 II/I 表达水平相对降低(P=0.008,图3A、B)。为进一步证实miR-140-5p 对自噬水平的影响,免疫荧光进行检测与western blot 结果一致,miR-140-5p 过表达组LC3 荧光表达减弱(图3C)。综上,过表达miR-140-5p 抑制CRC 细胞自噬的表达水平。
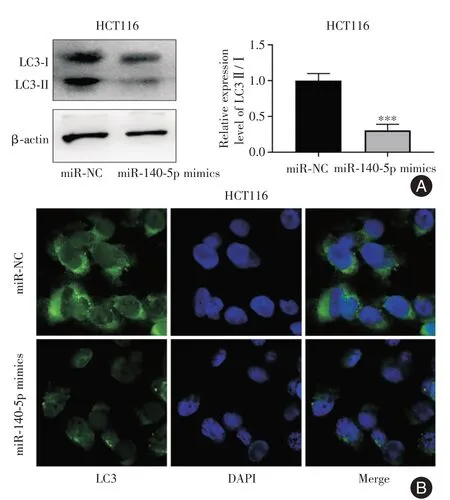
图3 过表达miR-140-5p 对CRC 细胞自噬的影响Fig.3 Effect of overexpression of miR-140-5p on autophagy of CRC cells
2.4 过表达miR-140-5p 合并雷帕霉素减弱CRC细胞对奥沙利铂的敏感性为验证miR-140-5p 是否通过自噬对奥沙利铂的敏感性产生作用,转染miR-140-5p mimics 并使用自噬激活剂雷帕霉素(rapamycin,Rapa)共同处理,免疫荧光检测发现,与过表达miR-140-5p相比,过表达miR-140-5p并处理Rapa 组的LC3 荧光表达增强(图4A)。CCK-8 结果表明在不同浓度的奥沙利铂处理下,miR-140-5p 加Rapa 组能够逆转miR-140-5p 组导致的增殖减少情况(P= 0.001 9, 图4B)。此外,选取在奥沙利铂10 μmol/L 进行克隆形成实验,发现与对照组相比,奥沙利铂处理下细胞克隆水平降低(P<0.000 1);过表达miR-140-5p加Rapa组的细胞菌落相比过表达miR-140-5p 组更多(P= 0.003 9,图4C)。以上结果表明,miR-140-5p 能调控自噬从而影响CRC 细胞对奥沙利铂的敏感性。
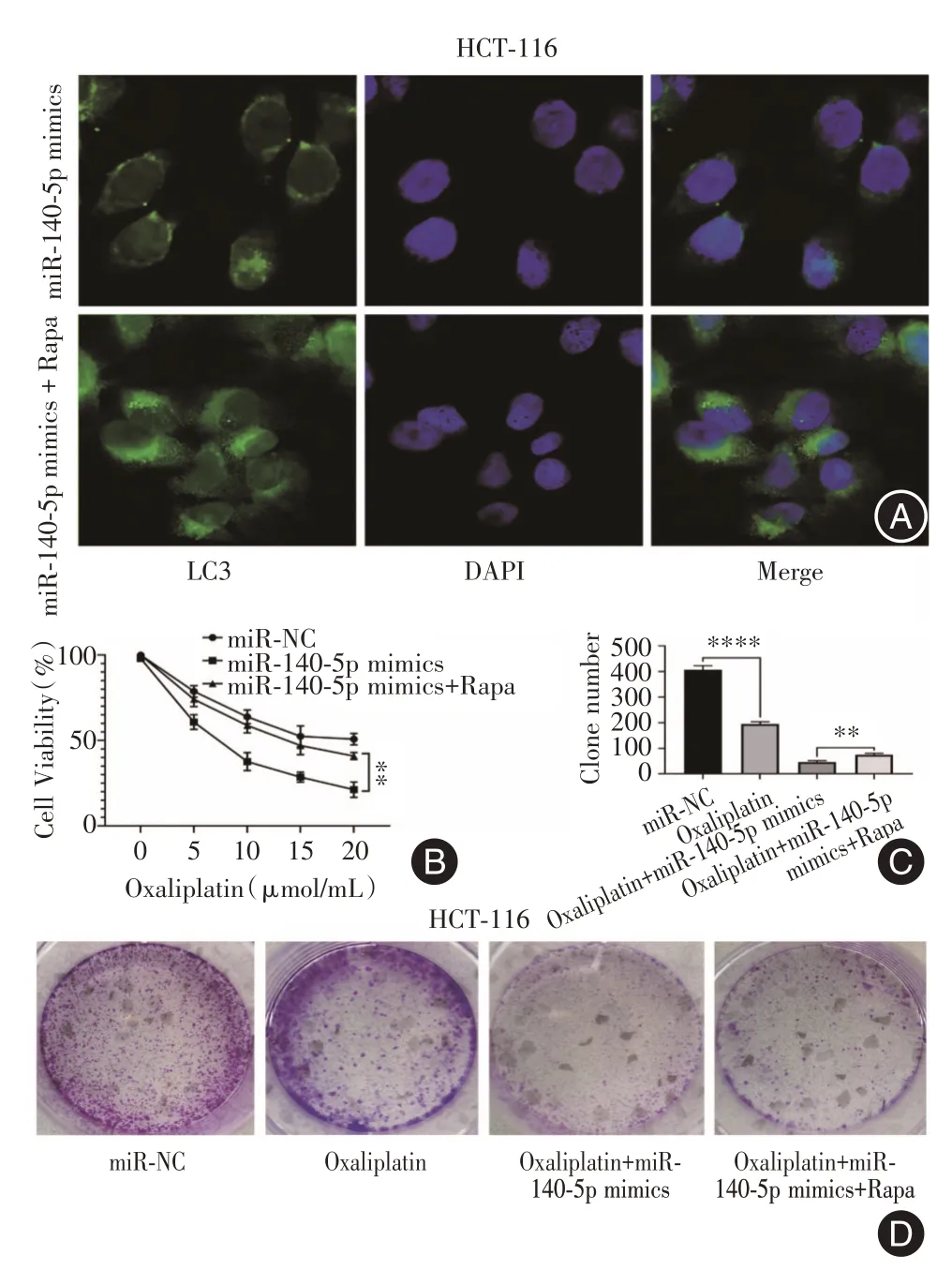
图4 过表达miR-140-5p 并促进自噬后,CRC 细胞对奥沙利铂的敏感性Fig.4 Sensitivity of CRC cells to oxaliplatin after overexpression of miR-140-5p and promotion of autophagy
3 讨论
在癌症治疗中,治疗耐药性是一个重大的挑战。奥沙利铂耐药是目前CRC 化疗的主要障碍之一。尽管肿瘤治疗取得了很大进展,但由于化疗耐药机制的复杂性,大多数临床有效的化疗诱导剂都是无效的[1]。
近年来,miRNA 的异常表达与CRC 化疗耐药的发生有关[12]。miR-302a 在西妥昔单抗耐药的CRC 细胞中的表达降低,过表达miR-302a 在功能上抑制了CRC 细胞的迁移、侵袭和转移,且miR-302a 的高表达恢复了CRC 细胞对野生型KRAS 和BRAF 基因的化疗反应性[13]。miR-140-5p 是被认为在CRC 的发展中起到抑癌的作用,其可通过直接靶向SOX4 抑制结直肠癌细胞增殖和侵袭[14]。此外,miR-140-5p 可抑制CRC 的进展和转移[15]。但miR-140-5p 调控CRC 耐药的分子机制尚未有研究报道。本研究结果表明,在CRC细胞中miR-140-5p 呈现低表达状态,而过表达miR-140-5p 后促进了CRC 细胞奥沙利铂的敏感性,本研究结果与前所述一致,这说明miR-140-5p 在CRC 中主要发挥抑癌的作用。然而miR-140-5p对CRC化疗药物敏感性的研究目前国内外尚无报道,本研究表明miR-140-5p 可以促进CRC 细胞对奥沙利铂的敏感性。
越来越多的文献表明,自噬可能有助于癌症发展。自噬激活可促进癌细胞存活(保护性自噬)或导致癌细胞死亡(细胞毒性/非保护性自噬)[16-17]。有研究表明阿帕替尼治疗CRC 细胞时可以诱导保护性自噬,而阿帕替尼与自噬抑制剂氯喹联合应用在体外和体内均具有最显著的抗肿瘤作用[18]。miR-489在大多数乳腺癌细胞和多个耐药的乳腺癌细胞中均处于下调状态,其可以通过抑制自噬来促进乳腺癌细胞对阿霉素的敏感性[19]。而miR-140-5p 在CRC 中的调控自噬的分子机制尚不清楚。本研究表明过表达miR-140-5p 可以抑制CRC 细胞的自噬并提高对奥沙利铂的敏感性,而过表达miR-140-5p 并使用自噬激活剂雷帕霉素时却降低了对奥沙利铂的敏感性,那么miR-140-5p参与调控CRC细胞自噬的哪个阶段,还有待进一步的探究。
综上所述,miR-140-5p 在CRC 细胞中低表达,其过表达通过抑制自噬促进了CRC 细胞对奥沙利铂的敏感性,不仅为CRC的临床诊治提供了有力的理论基础,还为CRC的靶向治疗提供了潜在的分子靶点。
