MKK7-mediated phosphorylation of JNKs regulates the proliferation and apoptosis of human spermatogonial stem cells
2021-12-24ZengHuiHuangChuanHuangXiRenJiWenJunZhouXueFengLuoQianLiuYuLinTangFeiGongWenBingZhu
Zeng-Hui Huang, Chuan Huang, Xi-Ren Ji, Wen-Jun Zhou, Xue-Feng Luo, Qian Liu, Yu-Lin Tang, Fei Gong,Wen-Bing Zhu
Zeng-Hui Huang, Xi-Ren Ji, Wen-Jun Zhou, Xue-Feng Luo, Qian Liu, Yu-Lin Tang, Fei Gong, Wen-Bing Zhu, Institute of Reproductive and Stem Cell Engineering, Central South University, Changsha 410008, Hunan Province, China
Zeng-Hui Huang, Department of Reproductive Center, Reproductive and Genetic Hospital of CITIC-Xiangya, Changsha 410008, Hunan Province, China
Chuan Huang, Department of Sperm Bank, Reproductive and Genetic Hospital of CITICXiangya, Changsha 410008, Hunan Province, China
Abstract BACKGROUND Human spermatogonial stem cells (SSCs) are the basis of spermatogenesis. However, little is known about the developmental regulatory mechanisms of SSC due to sample origin and species differences.AIM To investigates the mechanisms involved in the proliferation of human SSC.METHODS The expression of mitogen-activated protein kinase kinase 7 (MKK7) in human testis was identified using immunohistochemistry and western blotting (WB). MKK7 was knocked down using small interfering RNA, and cell proliferation and apoptosis were detected by WB, EdU, cell counting kit-8 and fluorescenceactivated cell sorting. After bioinformatic analysis, the interaction of MKK7 with c-Jun N-terminal kinases ( JNKs ) was verified by protein co-immunoprecipitation and WB. The phosphorylation of JNKs was inhibited by SP600125, and the phenotypic changes were detected by WB, cell counting kit-8 and fluorescenceactivated cell sorting.RESULTS MKK7 is mainly expressed in human SSCs, and MKK7 knockdown inhibits SSC proliferation and promotes their apoptosis. MKK7 mediated the phosphorylation of JNKs, and after inhibiting the phosphorylation of JNKs, the phenotypic changes of the cells were similar to those after MKK7 downregulation. The expression of MKK7 was significantly downregulated in patients with abnormal spermatogenesis, suggesting that abnormal MKK7 may be associated with spermatogenesis impairment.CONCLUSION MKK7 regulates the proliferation and apoptosis of human SSC by mediating the phosphorylation of JNKs.
Key Words: MKK7; Spermatogonial stem cell; Proliferation; Apoptosis; JNKs
INTRODUCTION
Infertility is an important global health problem that affects approximately 15% of couples of childbearing age[1]. For about 50% of such couples, male factors are a contributing cause[2]. There are many possible types of male subfertility, including idiopathic, acquired and congenital[3]. Although most male infertility problems can be solved through assisted reproductive technology, there are still some patients who are unable to have genetic offspring, especially those with non-obstructive azoospermia (NOA)[4]. Thus, it is necessary to find other effective treatments, such as spermatogonial stem cell (SSC) transplantation andin vitrospermatogenesis.
SSCs are present on the basement membrane of the seminiferous tubules of the testis, and these cells self-renew and differentiate for life-long spermatogenesis[5]. Intricate molecular and cellular interactions form the niche for SSC development[6]. SSC self-renewal is promoted by the secretion of glial cell line-derived neurotrophic factor (GDNF) from the niche[7].In vitro, GDNF and fibroblast growth factor 2 (FGF2) were observed to function synergistically to promote the growth of undifferentiated spermatogonia[8]. However, the function of basic FGF in SSC maintenancein vivoremains poorly defined[9]. Common molecules are activated by GDNF and FGF2 through Src family kinases, resulting in Ras activation[10]. Glucocorticoid-induced leucine zipper protein antagonizes Ras and stimulates the protein kinase B and MAPK/ERK kinase pathways[11]. When active protein kinase B or mitogen-activated protein kinase kinase 1 (a downstream molecule of the MAPK/ERK kinase pathway) were overexpressed, SSCs transfected with protein kinase B or mitogen-activated protein kinase kinase 1 could proliferate only in the presence of FGF2 or GDNF[12,13], respectively. Maintenance of the self-renewing state requires these pathways to upregulate certain genes, including ETS variant transcription factor 5, LIM homeobox 1, BCL6B transcription repressor and Nanos C2HC-type zinc finger 2[14].
Although studies have revealed a number of regulators of SSC self-renewal in mice, the regulatory mechanisms of SSCs are significantly different in humans and rodents[15]; these results cannot be repeated in human SSCs. In mouse, stemness occurs within the single spermatogonia; by contrast, the chains and pairs of spermatogonia are believed to represent committed progenitors[16,17]. In humans, Adark spermatogonia are regarded as the actual stem cells and slowly produce Apale spermatogonia, which have enough self-renewal capacity to maintain the stem cell pool[18-20]. Moreover, in mouse, the seminiferous epithelium cycle usually comprises 12 stages; however, only 6 stages have been distinguished in humans[21,22]. Notably, mouse and human SSCs have some common biomarkers; however, others differ considerably. For example, octamer-binding protein 4, a marker of mouse SSCs, is absent in humans[23,24]. Likewise, GDNF family receptor alpha 1 (GFRA1) positive spermatogonia are rare in mouse seminiferous tubules but abundant in humans[15].
Mitogen-activated protein kinase kinase 7 (MKK7) is an activator of c-Jun Nterminal kinase (JNK). MKK7 directly activates JNK1 and JNK2 (stressactivated protein kinases)viaphosphorylation[25-27]. JNK signaling has an important function in self-renewal, apoptosis and cell proliferation and was reported to be associated with reactive oxygen species-mediated mouse SSC self-renewal[28]. In the present study, we found that MKK7 was mainly expressed in human SSCs [GFRA1+/proliferating cell nuclear antigen (PCNA)+/KIT proto-oncogene, receptor tyrosine kinase (KIT)-] with proliferative activity. Deletion of MKK7 promoted human SSC apoptosis and impaired their proliferation. We further confirmed that MKK7 regulates SSC proliferation by inducing JNK1/JNK2/JNK3 phosphorylation. Additionally, MKK7 levels were confirmed to be decreased in patients with NOA compared with those in patients with obstructive azoospermia with normal spermatogenesis, particularly in patients with spermatogonial or spermatocyte maturation arrest. Our results reveal the molecular mechanisms responsible for human SSC proliferation and apoptosis and provide the basis for further research into the etiology, molecular diagnosis and therapy for male infertility.
MATERIALS AND METHODS
Collection of human testes samples
The Ethics Committee of the Reproductive & Genetic Hospital of CITICXiangya, Basic Medical Science School, Central South University approved and supervised the present study (approval No.: LL-SC-2020-028). The samples used in the experiments were provided by donors who supplied informed consent. All testis samples were derived from patients undergoing microdissection testicular sperm extraction, and patients with spermatogenic failure because of known hereditary factors, such as Klinefelter syndrome and Y chromosome microdeletions, were excluded. We collected a total of 16 testicular biopsies weighing 30-50 mg and classified them according to the results of hematoxylin-eosin staining, including normal, spermatogenic failure and Sertoli cell only syndrome. Testis tissue samples were rinsed thrice in Dulbecco’s modified Eagle’s medium (DMEM) comprising 10 mL/L streptomycin and penicillin and then fixed with 40 g/L paraformaldehyde or stored in liquid nitrogen.
Culture of a human SSC line
The method described by Houet al[29] was used to establish a human SSC line, which was a gift from He ZP, Hunan Normal University, Changsha. Human SSCs were cultured in DMEM/F12 (Gibco, Grand Island, NY, United States) containing 100 mL/L fetal bovine serum (Gibco) and 100 unit/mL streptomycin and penicillin (Invitrogen, Carlsbad, CA, United States) at 34 °C in a 50 mL/L CO2incubator. Every 4 d, the cells were passaged using 0.53 mmol/L EDTA with 0.5 g/L trypsin (Invitrogen).
Extraction of RNA, reverse transcription polymerase chain reaction and quantitative real-time polymerase chain reaction
RNAiso Plus reagent (Takara, Kusatsu, Japan) was used to extract total RNA from cells following the supplier’s protocol. Total RNA quality and concentration were measured using a Nanodrop instrument (Thermo Scientific, Waltham, MA, United States). The total RNA was converted to cDNAviareverse transcription PCR using a First Strand cDNA Synthesis Kit (Thermo Scientific) and following a previously described method[30]. The cDNA was then subjected to quantitative real-time PCR (qPCR) using SYBR Premix Ex Taq II (Takara) and comprising the following reaction conditions: 95 °C for 5 min; followed by 32 cycles of denaturation at 95 °C for 30 s, annealing at 52-60 °C for 45 s (see Supplementary Table 1 for the specific annealing temperatures) and elongation at 72 °C for 45 s. The negative control comprised RNA without reverse transcription but with qPCR. An Applied Biosystems ABI Prism 7700 system (Applied Biosystems, Foster City, CA, United States) was used to perform qPCR on triplicate samples. The amplicons were separatedviaelectrophoresis through 20 g/L agarose gels with ethidium bromide visualization. On the recorded images, chemiluminescence was used to analyze the band intensities (Chemi-Doc XRS, Bio-Rad, Hercules, CA, United States). The data were normalized to the expression of beta actin using the comparative cycle threshold method to calculate the relative mRNA expression level for each sample[31]. The designed primer sets for qPCR are listed in Supplemen tary Table 1.
Immunocytochemistry, immunohistochemistry and immunofluorescence
The human SSCs were rinsed thrice using cold phosphate buffer saline (PBS) (Gibco) and subjected to fixation for 15 min in 40 g/L paraformaldehyde. After a further three rinses with PBS, the cells were permeabilized for 10 min using 2.5 mL/L Triton X-100 (Sigma, St. Louis, MO, United States). The cells were blocked for 1 h at room temperature in 5% bovine serum albumin before being incubated with primary antibodies overnight at 4 °C. Supplementary Table 2 provides the detailed information regarding the antibodies used. After extensive washes with PBS, Alexa Fluor 488-conjugated immunoglobulin (Ig) G or Alexa Fluor 594-conjugated IgG were used as the secondary antibodies, and the nuclei were stained using 4,6-diamidino-2-phenylindole. Images were captured using a fluorescence microscope (Carl Zeiss, Oberkochen, Germany).
Xylene was used to deparaffinize testis sections, which were then rehydrated using a graded series of ethanol concentrations. Antigens were then retrieved by heating the samples at 98 °C for 18 min in 0.01 mol/L sodium citrate buffer. For immunohistochemistry but not for immunofluorescence, endogenous peroxidase activity was blocked using 30 mL/L H2O2(Zsbio, Beijing, China). The sections were then subjected to permeabilization for 15 min using 2.5 mL/L Triton X-100 (Sigma). The sections were blocked at room temperature for 1 h using 50 mL/L bovine serum albumin for 1 h and then incubated with primary antibodies overnight at 4 °C. Following extensive washing using PBS for immunohistochemistry, the sections were incubated at room temperature for 1 h with horseradish peroxidaseconjugated secondary antibodies, and then chromogen detection was carried out using a 3,3’-diaminobenzidine chromogen kit (Dako, Glostrup, Denmark). The sections were finally stained with hematoxylin. For immunofluorescence, the sections were incubated at room temperature for 1 h with Alexa Fluorconjugated secondary antibody, and the nuclei were stained with 4,6-diamidino-2-phenylindole. The images were captured under a Zeiss microscope.
Western blotting and immunoprecipitation
Testis samples and human SSCs were ground and lysed on ice for 30 min using radioimmunoprecipitation assay lysis buffer (Thermo Scientific) and then subjected to centrifugation at 12000gto produce clear lysates. A BCA kit (Thermo Scientific) was then used to determine the protein concentration in the lysates. Primary antibodies or control rabbit IgG were added to cell lysates and incubated at 4 °C overnight. The next day the supernatants were added with Protein G magnetic beads and incubated at 4 °C for 2 h. The samples were washed three times with washing buffer, the beads were pelleted magnetically, and then resuspended and boiled at 95 °C for 5 min. For each sample, 30 micrograms of total protein extracts were subjected to SDS-PAGE (Bio-Rad), and western blotting was performed following a previously published protocol[30]. Supplementary Table 2 details the antibodies used and their dilution ratios. Chemiluminescence (Bio-Rad) was used to visualize the intensities of the immunoreactive protein bands.
Small interfering RNA transfection
RiboBio (Guangzhou, China) synthesized the small interfering RNA (siRNA) sequences targeting humanMKK7mRNA, which are listed in Supplementary Table 3. Scrambled siRNAs were used as negative controls. Lipofectamine 3000 (Life Technologies, Carlsbad, CA, United States) was used to transfect the control-siRNA orMKK7-siRNAs (both 100 nmol/L) into human SSCs, following the manufacturer’s instructions. At 48 h after transfection, the cells were harvested to evaluate the changes in the expression levels of genes and proteins.
Cell counting kit-8 assay
After siRNA transfection, human SSC proliferation was detected using a cell counting kit-8 (CCK-8) assay (Dojindo, Kumamoto, Japan) following the manufacturer’s protocol. Briefly, 100 mL/L CCK-8 reagent replaced the cell culture medium, and the cells were incubated for 3 h. A microplate reader (Thermo Scientific) was then used to measure the absorbance at 450 nm.
EdU incorporation assay
Human SSCs were seeded at 5000 cells/well in a 96-well plate; each well contained DMEM/F12 medium containing 50 μmol/L EdU (RiboBio). The cells were cultured for 12 h, washed using DMEM, and subjected to fixation using 40 g/L paraformaldehyde. Cell neutralization was achieved using 2 mg/mL glycine, followed by permeabilization for 10 min using 5 mL/L Triton X-100 at room temperature. Apollo staining reaction buffer was then used to reveal the EdU staining. Cell nuclei were stained using Hoechst 33342. A fluorescence microscope (Zeiss) was used to capture images, and the positive rate of EdU staining was calculated by counting at least 500 cells.
Staining with Annexin V/propidium iodide and flow cytometry
Human SSCs were transfected with siRNAs for 48 h and then assessed for their apoptotic percentage. Cells were digested with trypsin, washed twice using cold PBS, and collected by centrifugation. Cells (at least 106) were resuspended in Annexin V Binding Buffer (BD Biosciences, Franklin Lakes, NJ, United States) following the manufacturer’s instructions. Then, 5 μL of Annexin Vconjugated Allophycocyanin and 10 μL of propidium iodide solution were added to the cells and incubated at room temperature for 15 min in the dark. The cells were then subjected to C6 flow cytometry analysis (BD Biosciences).
Terminal deoxynulceotidyl transferase nick-end-labeling assay
An In Situ Cell Death Detection Kit (Roche, Mannheim, Germany) was used to determine the percentage of apoptosis among human SSCs, following the manufacturer's instructions. Proteinase K (20 mg/mL) was used to treat the cells for 15 min at room temperature. Thereafter, the cells were incubated with fluorescein isothiocyanate-12-deoxyuridine 5’-triphosphate labeling/terminal deoxynucleotidyl transferase enzyme buffer in the dark for 1 h. Cell nuclei were then labeled using 4,6-diamidino-2-phenylindole. The negative control cells were treated with PBS without the terminal deoxynucleotidyl transferase enzyme. At least 500 cells were evaluated per sample under a Zeiss fluorescence microscope.
Statistical analysis
All the statistical review of this study was performed by a biomedical statistician. GraphPad Prism 8.0 (GraphPad software, La Jolla, CA, United States) was used for the descriptive and statistical analyses. All experiments were performed at least in triplicate, and all data are presented as the mean ± SD. At-test was used to calculate the statistical difference between two groups, withP< 0.05 being considered as statistically significant.
RESULTS
In humans, MKK7 is mainly expressed in SSCs
To explore MKK7 function, we first examined its expression in the adult human testis. The MKK7 protein level was measured in testis samples from patients with obstructive azoospermia with normal spermatogenesis using western blotting (Figure 1A). We further analyzed the localization of MKK7 in testicular tissue. MKK7 was mainly found in the cytoplasm of cells close to the seminiferous tubule basement membrane (Figure 1B), indicating that it might be expressed in SSCs. The results of double immunofluorescence staining showed that most MKK7-expressing cells were DEADbox helicase 4-positive germ cells, and about 76% of MKK7 positive germ cells expressed GFRA1. Interestingly, about 90% of MKK7-expressing cells were PCNA positive (Figure 1C). Taken together, the results suggested that MKK7 was mainly expressed in SSCs and might be involved in cell proliferation.
MKK7 knockdown inhibits human SSC proliferation
We then used a human SSC cell line to explore the role of MKK7 in SSC proliferation. First, quantitative real time PCR was used to confirm the identity of the human SSC line. The results indicated that the cells expressed many markers of human SSCs, includingGFRA1, Thy-1 cell surface antigen, ubiquitin C-terminal hydrolase L1 and DEAD-box helicase 4, while a hallmark of human Sertoli cells (encoding SRY-box transcription factor 9) was not detected (Supplementary Figure 1A). The results of immunofluorescence also showed that almost all cells expressed SSC markers, such as GFRA1, ubiquitin C-terminal hydrolase L1 and Thy-1 cell surface antigen (Supplementary Figure 1B). These results suggested that the SSC line had similar properties to primary SSCs.
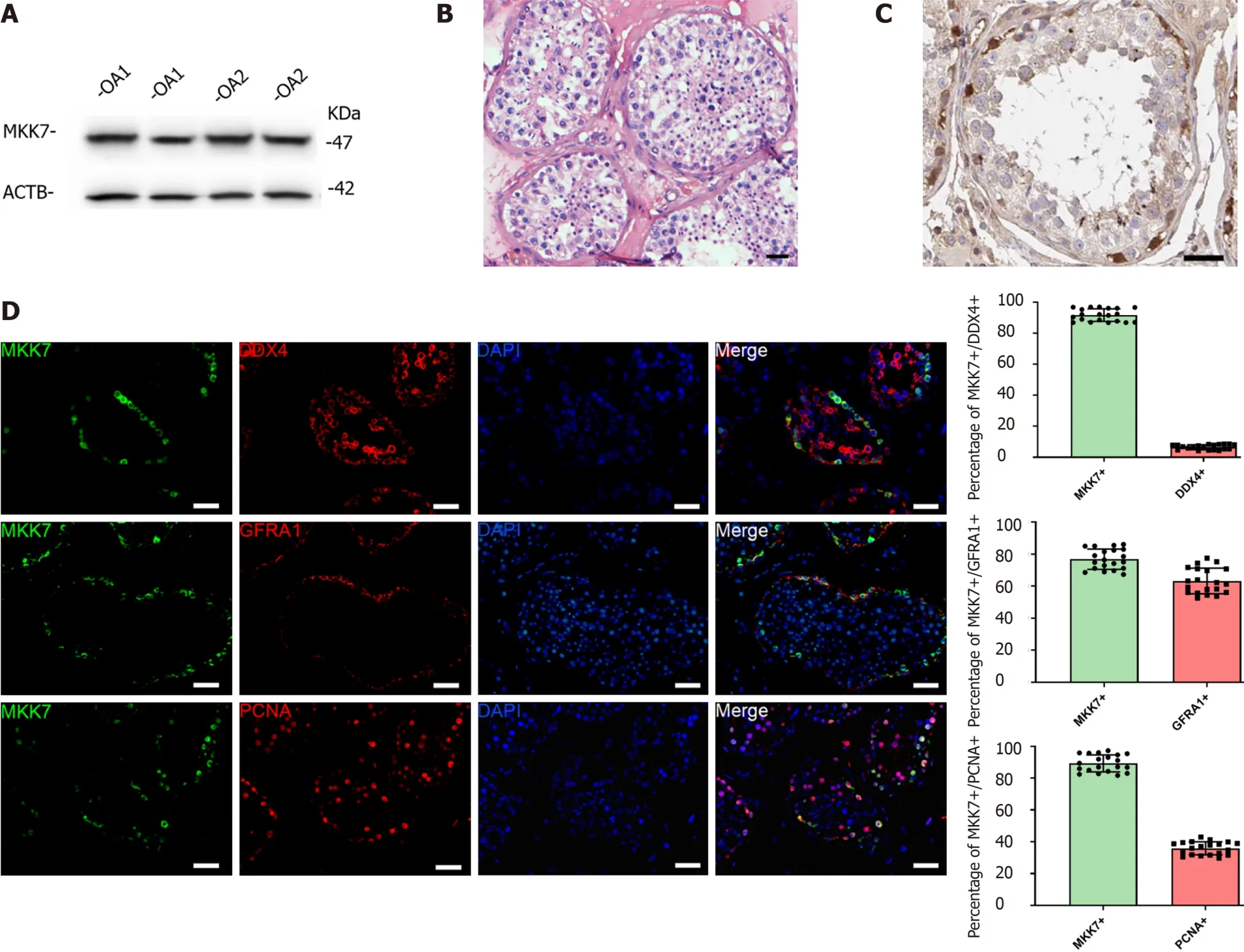
Figure 1 Mitogen-activated protein kinase kinase 7 was mainly expressed in human spermatogonial stem cells.
To explore the function of MKK7 in SSCs, siRNA-triggered knockdown ofMKK7was performed in the human SSC line. We examined the knockdown efficiency using quantitative real time PCR (Figure 2A) and western blotting (Figure 2B and C), which showed that MKK7 expression was attenuated byMKK7-siRNA1,MKK7-siRNA2 andMKK7-siRNA3.MKK7-siRNA3 showed the best knockdown effect. We then investigated the proliferative ability ofMKK7siRNA3-transfected SSCs using the CCK-8 assay (Figure 2D). We observed thatMKK7knockdown repressed SSC proliferation at 2 d to 5 d after transduction. We also found that the level of PCNA (a cell proliferation hallmark) was significantly reduced (Figure 2E and F). Likewise, at 48 h after transfection, we examined cellular DNA synthesis using an EdU assay. The proportion of EdU-positive cells was decreased compared with that in the control siRNA transfected cells (35.73% ± 0.64%vs22.05% ± 1.58%,P< 0.05) (Figure 2G and H).
MKK7 deficiency promoted human SSC apoptosis
Next, Annexin V/propidium iodide staining and flow cytometry were used to further examine cell apoptosis.MKK7silencing increased both early and late apoptosis of the human SSC line compared to that in the cells transfected with the controlsiRNA (early apoptosis: 5.17% ± 0.37%vs0.62% ± 0.19%,P< 0.05; late apoptosis: 4.58% ± 0.40%vs0.84% ± 0.09%,P< 0.05) (Figure 3A-C). Similarly, the results of terminal deoxynucleotidyl transferase nick-end-labeling (TUNEL) assay displayed that the proportion of TUNEL positive cells increased afterMKK7silencing compared with that in the control siRNAtransfected cells (14.34% ± 1.83%vs6.01% ± 0.95%,P< 0.05) (Figure 3D and E). Taken together, the results suggested that MKK7 promotes DNA synthesis and cell proliferation, whereasMKK7knockdown causes apoptosis.
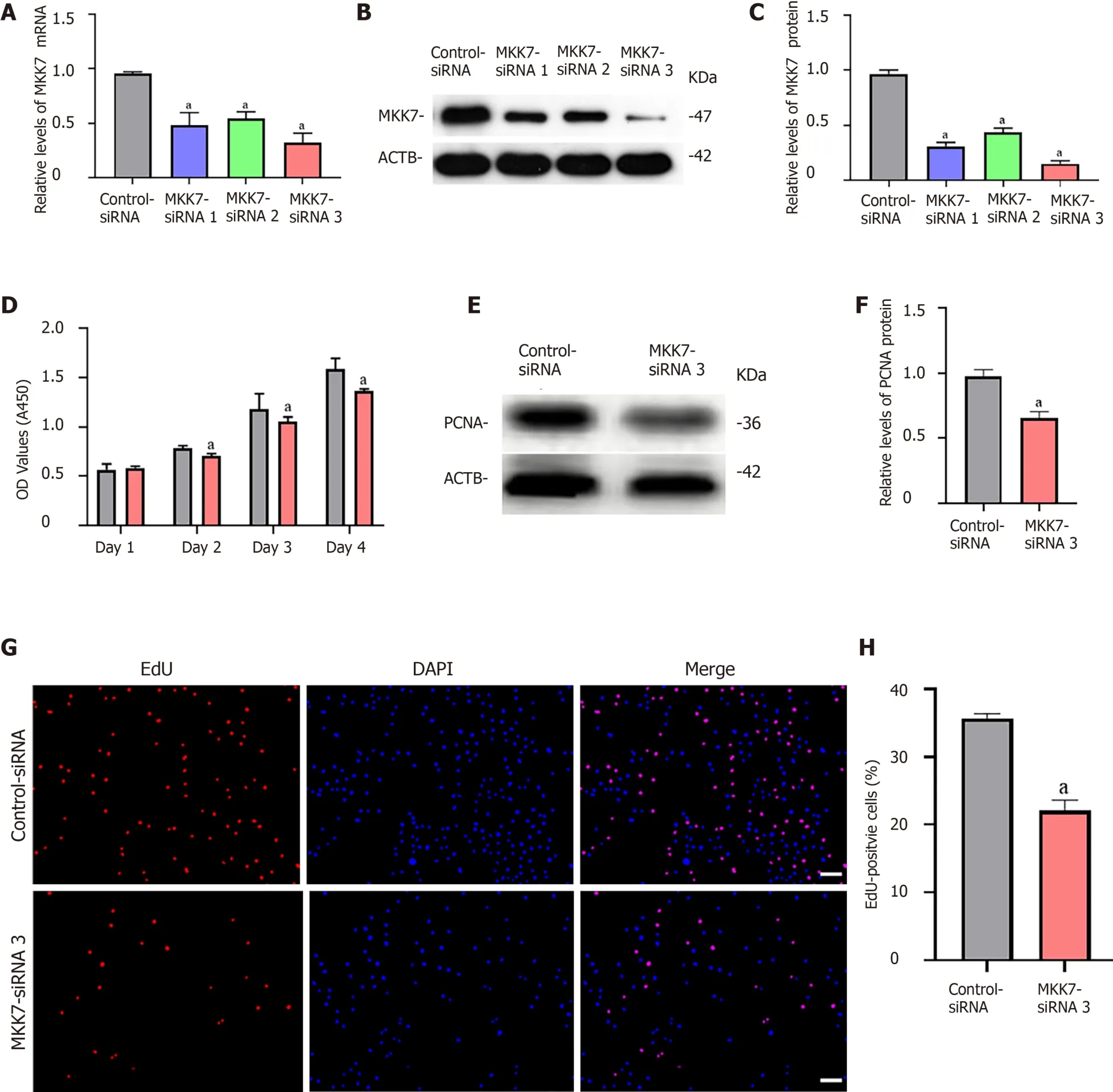
Figure 2 The influence of mitogen-activated protein kinase kinase 7 knockdown on human spermatogonial stem cell proliferation.
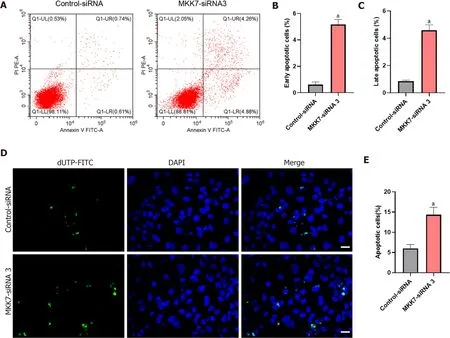
Figure 3 The influence of mitogen-activated protein kinase kinase 7 knockdown on the apoptosis of human spermatogonial stem cells.
MKK7 promotes the phosphorylation of JNKs
We further investigated the targets of MKK7. We predicted the targets of MKK7 using bioinformatic analysis, (GeneMANIA, HitPredict and String). Comprehensive prediction using the three databases identified JNK1, JNK2 and mitogen-activated protein kinase kinase kinase 7 as MKK7interacting proteins (Figure 4A). Mitogenactivated protein kinase kinase kinase 7 was reported as an upstream activator of the MKK/JNK signal transduction pathway, actingviaphosphorylation of MKK7[32], and MKK7 was reported to activate JNKs by phosphorylation[26]; therefore, JNKs might be candidate downstream targets of MKK7 in SSCs. To examine whether there was a physical interaction between MKK7 and JNKs, an immunoprecipitation assay followed by western blotting was carried out (Figure 4B), which showed that MKK7 could bind directly to JNKs in the human SSC line. By contrast, the control IgG did not pull down JNKs. We further detected the levels of JNKs and phosphorylated JNKs in human SSCs transfected withMKK7-siRNA3 (Figure 4C). We found that the levels of phosphorylated JNKs were reduced significantly compared with those in cells transfected with the control siRNA; however, the total level of JNKs did not change significantly. This result indicated that MKK7 could bind directly to JNKs to promote their phosphorylation.
Inhibition of JNKs decreases proliferation and promotes apoptosis of human SSC
To verify whether MKK7 affects SSC proliferation through phosphorylation of JNKs, we blocked the function of JNKs using SP600125, a selective inhibitor of JNK phosphorylation. We detected the level of phosphorylated JNKs after SP600125 treatment for 24 h, which showed that the levels of phosphorylated JNKs were notably reduced compared with those in the untreated control group (Figure 5A and B). In addition, PCNA protein levels decreased significantly (Figure 5C and D). Then, we examined the proliferation of human SSCs using the CCK-8 assay. SP600125 treatment significantly inhibited cell proliferation compared with that in the control group (Figure 5E). In addition, the proportion of EdU-positive cells decreased after culture with SP600125 for 24 h (34.17% ± 1.56%vs21.84% ± 1.62%,P< 0.05) (Figure 5F and G). We further analyzed the apoptosis of human SSCs using the TUNEL assay, which showed that TUNEL positive cells increased significantly (5.13% ± 0.34%vs12.18% ± 1.03%,P< 0.05) after SP600125 treatment (Figure 5H and I). Likewise, SP600125 treatment caused an increase in early apoptosis but not late apoptosis of SSCs according to Annexin V/propidium iodide staining and flow cytometry [1.80% ± 0.25% (control)vs4.61% ± 0.45% (SP600125),P< 0.05] (Figure 5J-L). Taken together, the results indicated that SP600125-mediated inhibition of JNK phosphorylation in SSCs resulted in impaired cell proliferation and increased cell apoptosis.
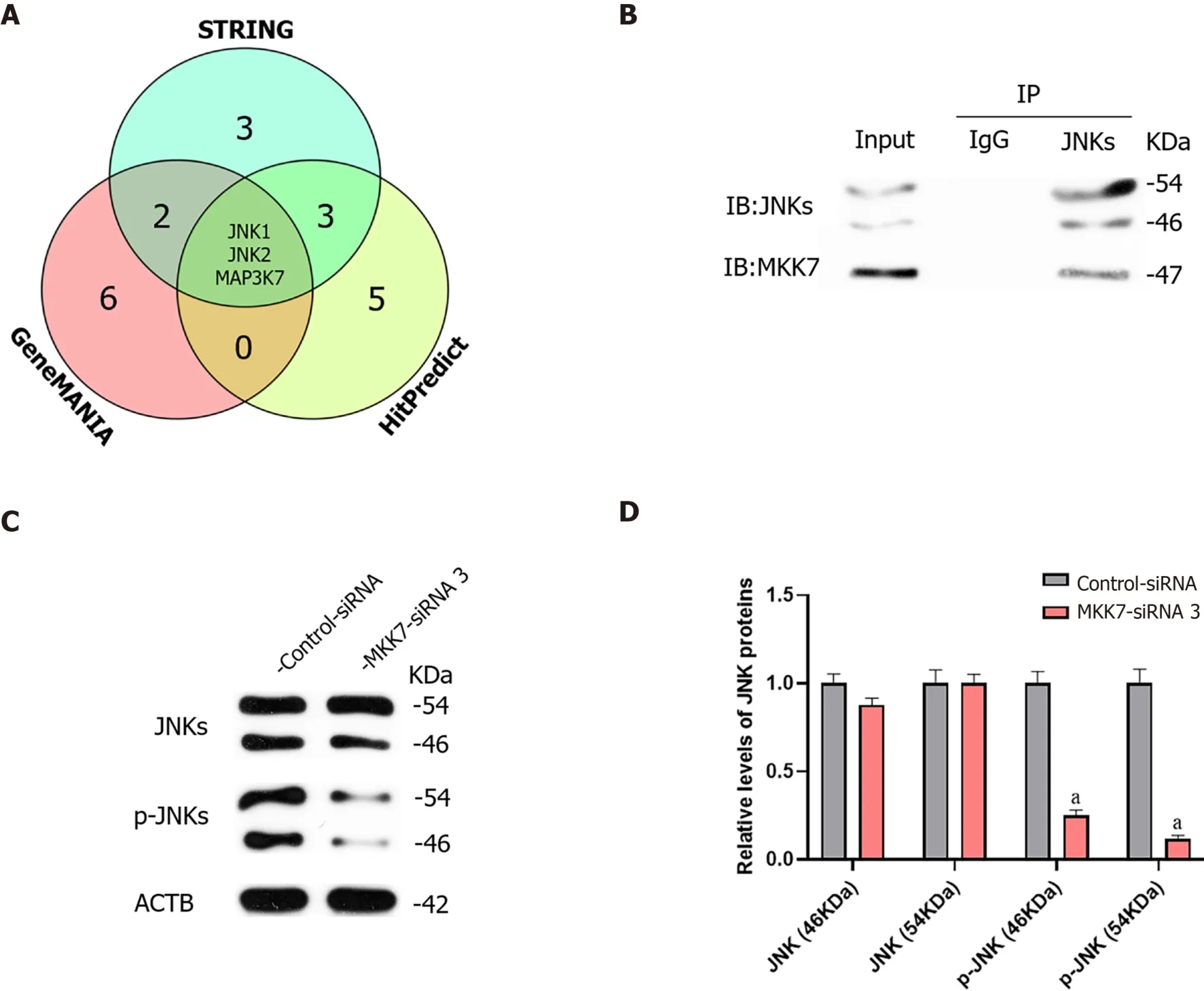
Figure 4 Mitogen-activated protein kinase kinase 7 affected the phosphorylation of c-Jun N-terminal kinases.
MKK7 downregulation correlates with NOA
NOA is a serious male infertility disease in clinical practice. According to the results of testicular pathological tissue, NOA can be divided into spermatogonia maturation arrest, spermatocyte maturation arrest, spermatid maturation arrest, hypospermatogenesis and Sertoli cell only syndrome. To investigate whether MKK7 is associated with impaired spermatogenesis in adults, we examined MKK7 expression patterns in eight testes. According to the results of hematoxylin-eosin staining, we confirmed the spermatogenesis status of the testis (Supplementary Figure 2A-H). We examined the positive proportion and localization changes of MKK7 (red) in SSCs using double immunohistochemistry with ubiquitin C-terminal hydrolase L1 (green). The results indicated that the percentage of MKK7-expressing SSC was significantly reduced in spermatogonia maturation arrest and spermatocyte maturation arrest samples compared to samples with normal spermatogenesis (Figure 6A and B), and the fluorescence intensity appeared to decrease in hypospermatogenesis samples, but there were no translocations of MKK7 protein observed in NOA samples. We further detected the relative levels of MKK7 protein by using western blots. The results displayed that the expression of MKK7 was significantly downregulated in all samples with impaired spermatogenesis compared to the normal group (Figure 6C and D). Those results implied that the aberrant expression of MKK7 was correlated with spermatogenesis disorder, especially the maturation of spermatogonia and spermatocytes.
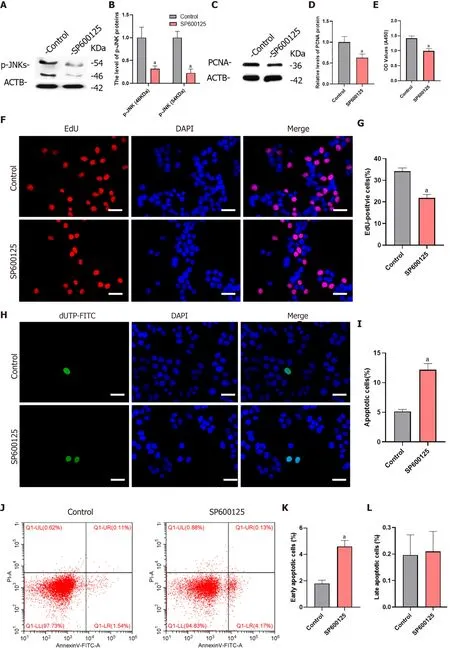
Figure 5 Inhibition of c-Jun N-terminal kinase 1 phosphorylation inhibited proliferation and promoted apoptosis of human spermatogonial stem cells.
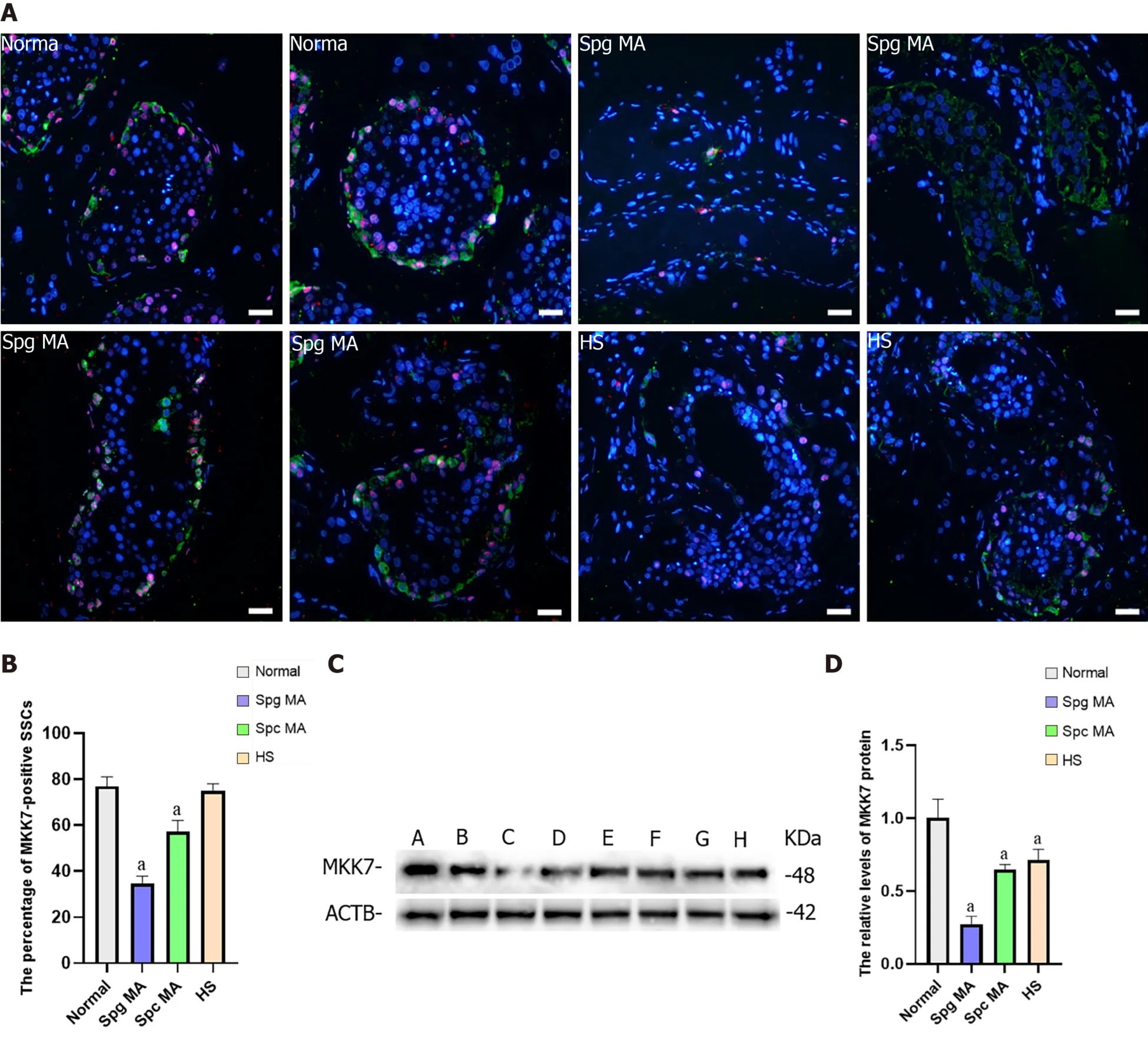
Figure 6 The expression of mitogen-activated protein kinase kinase 7 in the testis of obstructive azoospermia patients and nonobstructive azoospermia patients.
DISCUSSION
Human SSCs are the origin of spermatogenesis[33]; however, research on human SSCs are hampered because of limited sources and scarce numbers[34,35]. Much is known about the mechanism of SSC regulation in rodents, and spermatogenesis has been reconstructed through SSC transplantation[36]. However, these results are not entirely applicable to primates because many biological processes of rodent SSCs are different from those of primates[15,37]. For example, the spermatogenic epithelial cycle in mice can usually be divided into 12 stages; however, humans generally have 6 stages[21,22]. Furthermore, octamer-binding protein 4, a biomarker of mouse SSCs, is not expressed in human SSCs[23,24]. Therefore, research on the mechanisms of human SSCs are hindered by these discrepancies. Human primary SSCs cannot be culturedin vitrofor a long time because of the weak proliferation of human SSCin vitro, and the culture system of mice SSCs is not suitable for human SSC. In addition, the proliferation of human SSCsin vitrois weak[34]. To establish the human SSC line, the human SV40 Large T antigen was overexpressed in human primary GPR125-positive undifferentiated spermatogonia[29,38]. These cells possess similar biological properties to human SSCs and possess an unlimited proliferative capacity, which assisted us in studying the molecular mechanisms of SSCs.
MKK7 is involved in the phosphorylation of JNKs and activates their downstream pathways[39]. However, the promotion of SSC proliferation by MKK7 was unexpected, given that its inhibition often enhances self-renewal division of other stem cells. In mouse embryonic stem cells, activation of c-Jun NH2-terminal kinase by CdCl2- or HgCl2requires MKK7[40], and it is also involved in the process of embryonic stem cell differentiation into cardiomyocytes[41]. This suggested that MKK7 regulation in SSCs is distinct from that in other stem cells. In our study, we found that MKK7 was primarily expressed in self-renewing SSCs (GFRA1+/PCNA+), which was consistent with the findings of single cell sequencing, in which MKK7 was mainly expressed in SSCs[42]. Therefore, we believe it may be associated with the proliferation or apoptosis of SSCs. We validated the functions of MKK7 in a human SSC cell line using siRNA, and the results showed that MKK7 deficiency inhibited proliferation and promoted apoptosis of SSCs. MKK7 has been reported to be involved in the apoptotic process in neural cells[43] and participates in the proliferation of hepatocytes and cancer cells[44,45]. The functional diversity of MKK7 in various cells might reflect different downstream targets. Our work provides more evidence for the functions of MKK7; however, the specific downstream effectors of MKK7 remain to be explored.
Previous reports and bioinformatic prediction suggested that MKK7 is involved in the phosphorylation modification of JNKs[39] and blocking JNKs using small molecule inhibitors similarly inhibited the proliferation of mouse SSCs[28]. The development of SSCs is regulated by many key growth factors, such as GDNF[46], FGF[12], epidermal growth factor[47] and LIF interleukin 6 family cytokine[8]. Other intracellular regulatory molecules, including promyelocytic leukemia zinc finger[48], Nanos C2HCtype zinc finger 2[49] and Spalt like transcription factor 4[50], have also been demonstrated to be involved in SSC fate determination. Although the phosphorylation of JNK1 and JNK2 were reported to increase after treatment with GDNF and FGF2 in mouse SSCs[28], whether MKK7 is involved in the signal transduction of these important regulatory factors or if other regulatory pathways participate in this process remains to be further investigated. It is also reported that cell division cycle 5 like[51] and TNF receptor associated factor 6[52] have direct interactions with MKK7, which suggests that MKK7 might also affect cell proliferation through other molecules. To explore this question, further investigations are needed, for example, protein coimmunoprecipitation combined with mass spectrometry and transcriptome sequencing.
Although the pathogenesis of NOA is largely unclear, our results displayed that MKK7 levels were reduced in the testis of patients with NOA compared with those in patients with obstructive azoospermia, especially in testis with spermatocyte maturation arrest and spermatogonia maturation arrest. A reduction was also observed in the proportion of MKK7-positive SSCs. Exploration of mutations in theMKK7gene in patients with NOA and the construction of point mutations or knockout mouse models should be used to confirm the functions of MKK7. In addition, the current study was carried out in a human SSC line, and the proliferation function of MKK7 was only tested in SSCs. Thus, we are unsure whether MKK7 is involved in the differentiation of progenitor cells, which also needs further studies in mouse models.
In conclusion, we revealed that MKK7 is expressed mainly in human SSCs and inhibits the apoptosis and enhances the proliferation of SSCsviaJNK phosphorylation. Although some questions remain, our study offers new clues regarding the pathways and genes involved in the determination of SSC fate in humans. The activation of MKK7 or JNK1 using small molecules might contribute to human SSC self-renewalin vitroand might identify molecular targets to diagnose and treat male infertility.
CONCLUSION
MKK7 regulates the proliferation and apoptosis of human SSC by mediating the phosphorylation of JNKs. Abnormal expression of MKK7 may impair human spermatogenesis.
ARTICLE HIGHLIGHTS
Research background
Human spermatogonial stem cells (SSCs) are the basis of spermatogenesis. However,little is known about the developmental regulatory mechanisms of SSC due to sample origin and species differences.
Research motivation
To investigates the mechanisms involved in the proliferation of human SSCs.
Research objectives
To investigate the functions and mechanisms of mitogen-activated protein kinase kinase 7 (MKK7) during proliferation and apoptosis in human SSCs.
Research methods
The expression of MKK7 in human testis was identified using immunohistochemistry and western blotting (WB). MKK7 was knocked down using small interfering RNA,and cell proliferation and apoptosis were detected by WB, EdU, cell counting kit-8 and fluorescence-activated cell sorting. After bioinformatic analysis, the interaction of MKK7 with c-Jun N-terminal kinases (JNKs) was verified by protein co-immunoprecipitation and WB. The phosphorylation of JNKs was inhibited by SP600125, and the phenotypic changes were detected by WB, cell counting kit-8 and fluorescenceactivated cell sorting.
Research results
MKK7 is mainly expressed in human SSCs, and MKK7 knockdown inhibits SSC proliferation and promotes their apoptosis. MKK7 mediated the phosphorylation of JNKs,and after inhibiting the phosphorylation of JNKs, the phenotypic changes of the cells were similar to those after MKK7 downregulation. The expression of MKK7 was significantly downregulated in patients with abnormal spermatogenesis, suggesting that abnormal MKK7 may be associated with spermatogenesis impairment.
Research conclusions
MKK7 regulates the proliferation and apoptosis of human SSC by mediating the phosphorylation of JNKs. Abnormal expression of MKK7 may impair human spermatogenesis.
Research perspec tives
This study intended to reveal the role and regulatory mechanism of MKK7 in the regulation of SSC development and spermatogenesis in humans, which can provide a scientific basis for the etiological interpretation and molecular diagnosis of male infertility. It also provided new molecular targets for the clinical treatment of male infertility and the development of contraceptives.
杂志排行

World Journal of Stem Cells的其它文章
- Priming strategies for controlling stem cell fate: Applications and challenges in dental tissue regeneration
- Epigenetic regulation of dental pulp stem cells and its potential in regenerative endodontics
- Effects of immune cells on mesenchymal stem cells during fracture healing
- Regulation of the mesenchymal stem cell fate by interleukin-17:Implications in osteogenic differentiation
- Why stem/progenitor cells lose their regenerative potential
- Nanofat: A therapeutic paradigm in regenerative medicine