豆象亚科昆虫性信息素研究进展
2021-12-21陈思雨李猷王秀荣武承旭
陈思雨 李猷 王秀荣 武承旭

摘 要:豆类作物是我国重要的经济作物,而豆象亚科(Bruchinae)中约4/5的种可蛀入豆荚内啃食豆粒,是其重要害虫。该类害虫因其钻蛀性为害的特点导致施用农药无法达到良好的防治效果,因此急需开发高效、无污染的防治技术。豆象亚科雌雄成虫均可产生和释放昆虫性信息素,对雌、雄成虫有较强的吸引力且会影响幼虫行为,这些性信息素组分构成相似,其中一些性信息素组分被豆象亚科昆虫种间共享,因而有望开发并应用于该类害虫的绿色防控。本文对国内外豆象亚科昆虫性信息素组分结构,合成及其影响因素,以及在应用等方面的研究开展综述,并对存在问题和未来研究进行展望,旨在为豆象亚科昆虫性信息素的研究和豆象亚科昆虫绿色防控提供参考。
关键词:豆象;性信息素;绿色防控
中图分类号:S7
文献标识码:A
文章编号:1008-0457(2021)06-0044-09
国际DOI编码:10.15958/j.cnki.sdnyswxb.2021.06.006
Abstract:Legume crops are important cash crops in China,and about 4/5 species of Bruchinae can feed legume grains in pods,which is an important pest of legume crops.It is urgent to develop high-effective and pollution-free control techniques because the habits and characteristics of borers affect the effect of spraying.Both male and female adults of Bruchinae could produce and release insect sex pheromones,which are attractive to both male and female adults,and affect larvae behavior.The composition of these pheromone components is similar,and some of them are shared among different species,it is expected to be developed and applied in the green control of this pest.In this paper,the structure,synthesis,influencing factors,and application of the sex pheromone components of Bruchinae insects at home and abroad are reviewed,as well as the existing problems and future research prospects,the purpose of this study are to provide a reference for the research and green control of the sex pheromone of the legume insects.
Keywords:Bruchinae;sex pheromone;green control
化學通讯是指生物个体之间,生物与环境间利用化学物质进行信息沟通的现象。昆虫之间及昆虫与寄主之间的信息化学物质往往由昆虫的某些特殊分泌器官和寄主挥发释放,被其他昆虫个体感受器接收,从而引起一定的行为或生理效应(如觅食、聚集、产卵等)[1]。其中,昆虫性信息素是由昆虫特殊腺体分泌,对同种异性个体具有强烈引诱作用,能够促进同种交配的极微量化学物质,而以性信息素为主导的辨识互动反应保证了同种昆虫之间通讯的畅通和异种之间的隔离[2]。
豆象亚科(Bruchinae)隶属鞘翅目(Coleoptera)叶甲总科(Chrysomeloidea)。目前,全世界已记载约60个属1700个种,其中多数取食豆科植物种子,是豆科植物的重要害虫[3]。例如绿豆象Callosobruchus chinensis、四纹豆象Callosobruchus maculatus、鹰嘴豆象Callosobruchus analis、西非花生豆象Callosobruchus subinnotatus、罗得西亚豆象Callosobruchus rhodesianus、菜豆象Acanthoscelides obtectus、巴西豆象Zabrotes subfasciatus、皂角豆象Megabruchidius dorsalis等,其中瘤背豆象属(Callosobruchus)中的绿豆象、四纹豆象、鹰嘴豆象、西非花生豆象、罗得西亚豆象,以及菜豆象和巴西豆象皆为我国进境植物检疫性有害生物[3]。豆象首先在豆荚上产卵,卵孵化为幼虫后蛀入豆荚内啃食豆粒,而且豆象会随豆子进入仓库,发生二次侵害,害虫的侵扰导致大量的定性和定量损害,表现为营养损失和发芽率低 [4-5]。由于豆象害虫钻蛀性危害的特点,利用农药进行防治的效果较差,且农药残留会污染环境,而利用昆虫性信息素防治害虫,具有无毒,高效,特异性强,无污染的优点[6]。
昆虫性信息素防治技术作为一种绿色行为调控技术,在鞘翅目、鳞翅目(主要是蛾类) 和双翅目等害虫的测报和防治中已经得到广泛应用[7]。Butenandt等[8]对家蚕性信息素进行分离提纯后,首次证明了化学通讯在昆虫界的存在,并科学地解释了化学通讯在生物间的化学联系。目前,对豆象昆虫信息素研究集中于性信息素,豆象昆虫性信息素的研究报道最早见于雄性菜豆象表皮化合物的提取与鉴定[9-10],本田博等[11]也研究了与绿豆象和四纹豆象的产卵行为和交配行为有关的生态化学物质。我国对豆象昆虫寄主挥发物和植物源杀虫剂的研究相对较多[12-14],而其性信息素的研究较少。
部分豆象性信息素虽已被分析鉴定,但用于防治或监测豆象害虫的性信息素的报道却很少。因此,本文针对国内外豆象昆虫性信息素组分结构,合成以及释放的影响因素,以及在应用等方面的研究开展综述,并对存在问题和未来研究进行展望,旨在为豆象害虫绿色防控提供参考。
1 豆象昆虫性信息素组分结构及功能
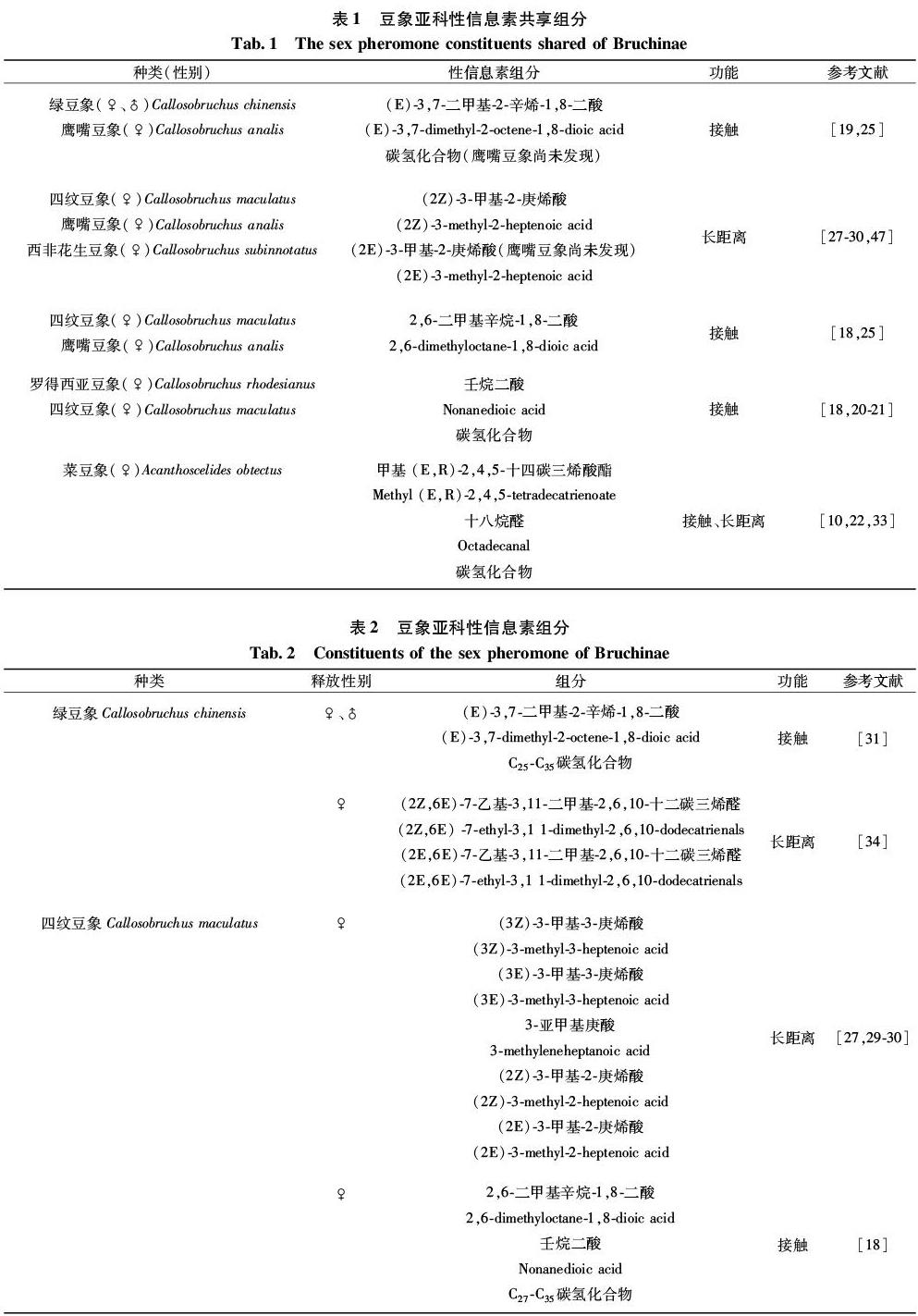
目前,国内外学者对豆象的性信息素研究多数局限于仓储危害严重的8种豆象,其中绿豆象、四纹豆象、鹰嘴豆象、罗得西亚豆象、西非花生豆象、菜豆象的性信息素已鉴定,另外2种巴西豆象、皂角豆象的性信息素尚未进行鉴定。按照性信息素的作用方式及范围,通常可将性信息素分为接触性信息素和长距离性信息素,接触性信息素有助于辨别伴侣和诱导求爱,而长距离性信息素挥发性高,有效的作用距离长,可以有效吸引距离较远的异性[15-16]。
1.1 组分结构
1.1.1 接触性信息素
豆象接触性信息素大部分由雌虫释放,一般具有C25-C35直链和甲基支链烃类,且含有C8-C19直链烷烃,碳链两端具有羧酸类官能团或碳链末端具有酮类、酯类、醛类官能团的化合物,化合物多数不含双键,因具有碳链较长的C25-C35碳氢化合物(表1,2),不易挥发,只有近距离吸引异性的效果[17-19]。绿豆象、四纹豆象、鹰嘴豆象的接触性信息素的组分相似,其主要化合物 (E)-3,7-二甲基-2-辛烯-1,8-二酸、2,6-二甲基辛烷-1,8-二酸均为单萜二羧酸,但绿豆象和四纹豆象的接触性信息素在混合物组成中更为相似,都由C25-C35直链和甲基支链烃类及C8主链和二甲基支链的单萜二羧酸构成[18,19]。罗得西亚豆象接触性信息素除了含有C27-C33碳氢化合物和二羧酸类化合物,还含有6,10,14-三甲基-2-十五烷酮和2-十九烷酮两种酮类化合物,其中壬烷二酸是与四纹豆象接触性信息素共有的组分[18,20-21]。菜豆象的接触性信息素除了含有C25-C31碳氢化合物外,还含有甲基 (E,R)-2,4,5-十四碳三烯酸酯和十八烷醛两种其他豆象接触性信息素中未发现的酯类和醛类化合物,与瘤背豆象属昆虫接触性信息素组分明显不同[10,22]。
具有相同分子式的化合物,立体异构体不同,化学性质和物理性质有所不同,例如生物体对于两个旋光異构体会做出偏好选择[23]。Yajima等[24]测定了绿豆象接触性信息素的立体异构体组成,根据合成性信息素样品得出旋光异构体R∶S为3.3~3.4∶1。虽然鹰嘴豆象与绿豆象、四纹豆象含有相同接触性信息素组分 (E)-3,7-二甲基-2-辛烯-1,8-二酸和2,6-二甲基辛烷-1,8-二酸,但化合物的立体异构体(2S,6R)-1∶ (S)-2=1.8∶1与绿豆象和四纹豆象不同[25],鹰嘴豆象的单萜二羧酸都表现出立体专一性,这与鹰嘴豆象雄虫通过识别雌虫表皮的接触性信息素只与同种雌虫进行交配的配偶识别特异性密切相关[18,26,20]。罗得西亚豆象雄性也可通过与其他同属物种主要接触性信息素化合物的分子立体结构差异实现只与同种雌性交配的配偶识别特异性[20]。然而绿豆象和四纹豆象的单萜二羧酸缺乏立体专一性,雄虫不能区分同种或异种雌虫[18,26]。
1.1.2 长距离性信息素
豆象的长距离性信息素主要由雌虫释放,主要为含有C7-C12直链烷烃,甲基或乙基分支,1~3个双键,碳链末端具有醛类、羧酸类、酯类官能团的化学结构(表1,2),能够吸引远距离的异性。(2Z)-3-甲基-2-庚烯酸是四纹豆象、鹰嘴豆象、西非花生豆象共有的组分,且三种豆象的长距离性信息素皆为含有C7直链烷烃,1个双键的短链脂肪酸的同源物[27-30]。绿豆象与罗得西亚豆象的长距离性信息素主要为醛类化合物,与其他瘤背豆象属的短链脂肪酸同系物不同的是绿豆象的性信息素在C7类倍半萜烯处有一个乙基[31];表明了绿豆象进一步的系统发育关系[32],菜豆象长距离性信息素主要为酯类化合物[33]。
豆象长距离性信息素通常由相同分子式化合物的不同立体异构体组成,如绿豆象的长距离性信息素由(2Z,6E)-7-乙基-3,11-二甲基-2,6,10-十二碳三烯醛及其E型异构体(2E,6E)-7-乙基-3,11-二甲基-2,6,10-十二碳三烯醛组成,通过气相色谱分析,估计信息素提取物中2Z,6E-和2E,6E-异构体的比例为1∶1[34],同时发现2Z异构体比其2E异构体对雄性绿豆象更具吸引力[31]。四纹豆象和西非花生豆象同样由相同分子式化合物的顺反异构体(2Z)-3-甲基-2-庚烯酸(鹰嘴豆象长距离性信息素组分)和(2E)-3-甲基-2-庚烯酸组成,推测瘤背豆象属(Callosobruchus)的其他种性信息素可能含有该化合物[28]。性信息素化合物旋光异构体的作用不尽相同,Shimomura等[35]通过Y型嗅觉仪选择试验发现雌性罗得西亚豆象的长距离性信息素(3S,6E)-7-乙基-3,11-二甲基-6,10-十二碳二烯醛的 (S)-对映异构体吸引雄性,而(R)-对映体和外消旋体不吸引雄性,表明(R)-对映体能抑制该化合物的活性。
1.2 组分的功能多样性
豆象性信息素组分不仅具有性引诱功能,也具有阻碍产卵、影响交配的功能,也可作为雄虫识别信号。豆象接触性信息素大部分由雌虫释放,而绿豆象的接触性信息素雌、雄虫皆可释放,但是只对雄性有影响[19],菜豆象的接触性信息素由雄虫释放,能够吸引雌性,是一类性引诱剂[9],其中组分甲基 (E,R)-2,4,5-十四碳三烯酸酯,被证明是一种阻碍素,菜豆象在豆类上释放该信息素,告知同种雌虫豆子上存在卵或幼虫,以此阻碍其产卵,使卵在豆类中的分布更加均匀,并减少幼虫之间的竞争[36-37],同时雄性在交配时将该化合物转移到雌性身上,导致求爱的雄性避开正在交配的雌性,因此推测该化合物还可作为雄虫识别信号[33,38]。豆象接触性信息素组分普遍含有C25-C35碳氢化合物,而且C25-C31碳氢化合物也被证明具有阻碍雌虫产卵的作用[22],同时碳氢化合物能显著增强豆象的交配行为,豆象表皮碳氢化合物也被作为配偶识别信号 [18]。
豆象性信息素在不同种间或昆虫间所具有的功能不尽相同。西非花生豆象与四纹豆象性信息素共享两种组分,但仅有西非花生豆象能夠引起四纹豆象雄虫反应,推测(3Z)-3-甲基-3-庚烯酸为四纹豆象性信息素的主要成分,则其可作为一种抑制西非花生豆象交配的拮抗剂[39]与菜豆象性信息素相似的化学组分反-11-十四碳烯乙酸酯在茶小卷叶蛾(Adoxophes honmai)的性信息素中也有发现,而且许多蛾类性信息素都含有C11-C16烯酸酯[8,40-41]。茶蚕(Andraca bipunctata)性信息素与菜豆象的接触性信息素都含有十八烷醛[8,42]。菜豆象的长距离性信息素(3Z,6E)-α-法呢烯也是吸引暗褐断眼天牛(Tetropium fuscum)雌雄虫的主要物质[43],(3Z,6E)-α-法呢烯和3E,6E)-α-法呢烯也是引诱植食性昆虫的植物挥发性信息化合物[44]。
2 豆象昆虫性信息素合成及应用
2.1 豆象昆虫性信息素合成
2.1.1合成方法
豆象昆虫接触性信息素主要为二羧酸类化合物。Bostedor等[45]确定了二羧酸类化合物可以来自法呢醇,并提出了法呢基焦磷酸转化成法呢醇,然后转化成法呢醇酸,最后转化成法呢醇衍生的二羧酸的代谢途径。
Chiluwal等[46]用不太复杂且经济可行的Z-选择方法(烯烃复分解反应,选择Z式结构烯烃)合成富含2Z,6E-异构体的绿豆象长距离性信息素。四纹豆象、鹰嘴豆象、西非花生豆象的长距离性信息素皆为短链脂肪酸的同源物,属同源半萜类化合物,一些同源二萜类化合物是通过甲羟戊酸酯和同源甲羟戊酸酯途径合成的,该信息素成分可能与其他同源半萜类化合物的生物合成途径相似[34]。罗得西亚豆象的长距离性信息素(3S,6E)-7-乙基-3,11-二甲基-6,10-十二碳二烯醛与核桃果峰斑螟(Acrobasis nuxvorella) 的性信息素组分(E,Z)-9,11-十二碳二烯醛相似,可以通过增加支链等方式达到合成目的[41]。
2.1.2影响因素
豆象性信息素的合成位置会影响化合物的活性,四纹豆象雌虫臀板的提取物较于腹部和胸部提取物能引起雄虫更强烈的反应[30]。豆象性信息素的不同合成物组合也会影响化合物的活性,四纹豆象长距离性信息素的5种已鉴定的二羧酸类化合物对四纹豆象雄虫均有活性,且2种或2种以上的组合具有相加效应,但在风洞实验中雄虫只对(3Z)-3-甲基-3-庚烯酸和(3E)-3-甲基-3-庚烯酸以及5种二羧酸类化合物的组合产生反应[30]。Shu等[47]对四纹豆象触角电位(EAG)和行为反应的研究显示,(3Z)-3-甲基-3-庚烯酸比(2Z)-3-甲基-2-庚烯酸更能引起雄虫反应,且以1∶3比例混合使用比单独使用更能引起雄虫反应,而与C27-C35碳氢化合物一起使用可显著增强雄性的交配行为[18]。同时,(3Z)-3-甲基-3-庚烯酸和(E)-3,7-二甲基-2-辛烯-1,8-二酸也是鹰嘴豆象进行交配所必需的性信息素组分。类似的,在菜豆象嗅觉仪测定试验中,6种挥发性信息素混合时对雌虫的吸引力强于任何单一化合物的吸引力,十八烷醛和甲基 (E,R)-2,4,5-十四碳三烯酸酯具有相加效应,甲基 (2E,4Z)-2,4-癸二烯酸酯和(3Z,6E)-α-法呢烯也能够增强十八烷醛和甲基 (2E,4Z,7Z)-2,4,7-癸三烯酸酯的活性[33]。在无定向气流的条件下,西非花生豆象在嗅觉仪中的潜伏期比在定向气流条件下长;当两种组分混合时,雄虫明显比存在单一组分时更早到达气味源,且当 (2Z)-3-甲基-2-庚烯酸和(2E)-3-甲基-2-庚烯酸以1∶1或3∶1的比例混合作为气味源时,西非花生豆象雄虫到达气味源的比例最大;使用单一组分作为气味源时,豆象对(2Z)-3-甲基-2-庚烯酸反应更快[39]。
人工性信息素合成过程中,产物的立体化学纯度是一个重要的影响因素,不同异构体及其比例直接影响合成物质对目标昆虫的活性[48]。如罗得西亚豆象(雌虫)的挥发性信息素(3S,6E)-7-乙基-3,11-二甲基-6,10-十二碳二烯醛的 (S)-对映异构体吸引罗得西亚豆象的雄性,(R)-对映体却抑制该化合物的活性[35]。Yajima等[24]发现 (E)-3,7-二甲基-2-辛烯-1,8-二酸的R异构体活性较S异构体低,并且测定出合成样品R∶S=3.3~3.4∶1时才有效诱导绿豆象进行交配。鹰嘴豆象接触性信息素的立体化学与活性的关系也与配偶识别特异性密切相关[20]。
2.2 豆象昆虫性信息素的应用
2.2.1 诱捕
昆虫性信息素是昆虫分泌到体外的超微量、强生理活性和种属特异的化学信使,可作为一种高效、特异性强、无污染的新型绿色防虫技术和其他信息化学物质可以通过大规模诱捕、引诱剂、驱避剂、威慑剂和干扰交配的方法干扰害虫两性间的化学通讯联络,减少两性间的交配率来控制害虫种群[49-51]。大规模诱捕是在田间大量诱杀雄虫个体,导致田间雌虫交配比率降低,从而控制田间虫口密度;交配干扰是在一定区域内释放大量性信息素,扰乱雄虫定向寻觅雌虫交配,打断田间雌雄虫的信息通讯,最终无法交配繁殖[52]。
豆象长距离性信息素组分化合物混合使用的效果高于单一化合物,化合物不同的比例也会导致效果不同[47]。如(2Z,6E)-7-乙基-3,11-二甲基-2,6,10-十二碳三烯醛和(2E,6E)-7-乙基-3,11-二甲基-2,6,10-十二碳三烯醛以1∶1的比例混合使用,可以显著吸引雄性绿豆象,以此作为性引诱剂,结合诱捕器的使用,可以在野外诱捕雄性绿豆象[34]。
2.2.2 种群监测
性信息素在农林业上最广泛和成功的另一种应用就是害虫种群监测。用性信息素监测害虫与灯光等传统害虫种群监测手段相比,具有灵敏性高、专一性强、使用简便,尤其是低密度下有选择的捕获害虫的特性,多年来在害虫管理的“推—拉”策略中日益流行[53]。因此,关于豆象的性信息素监测研究也较多。如四纹豆象挥发性信息素的5种已鉴定的二羧酸类化合物会引起雄性四纹豆象的远距离定向移动,所以该性信息素可以作为引诱剂来监测四纹豆象的种群[30]。西非花生豆象性信息素——(2Z)-3-甲基-2-庚烯酸和 (2E)-3-甲基-2-庚烯酸,当以每种组分不少于25%作为引诱剂时,超过60%的雄性可被诱捕器捕获,这种引诱剂可用于花生种植园中西非花生豆象种群的监测[39]。Pimbert等[54]的嗅觉仪选择试验表明,巴西豆象雌性会产生一种性信息素,吸引雄性定向移动,可以利用该性信息素监测巴西豆象种群。同时,雌性巴西豆象的气味在雄性中引起强烈的触角电位反应(EAG),宿主植物挥发的气味会加强这种性信息素对雄性的吸引[55]。
表皮化合物是昆虫重要的信号特征,是昆虫代谢和生物学的重要决定因素,皂角豆象的表皮化合物表现出显著的两性异形,皂角豆象的年龄和交配状态也可通过表皮化合物区分,可根据该表皮化合物进行种群监测[56-58]。
3 展望
豆象性信息素多数为几种化合物的组合,少数为单一化合物,如鹰嘴豆象挥发性信息素(2Z)-3-甲基-2-庚烯酸。豆象性信息素具有高度物种特异性,如鹰嘴豆象和罗得西亚豆象雄性通过与其他同属物种主要的接触性信息素化合物的分子立体结构差异来实现配偶识别特异性,只与同种异性交配[16]。豆象也共享部分性信息素组分,如(E)-3,7-二甲基-2-辛烯-1,8-二酸是绿豆象和鹰嘴豆象共有的接触性信息素组分;(2Z)-3-甲基-2-庚烯酸,(2E)-3-甲基-2-庚烯酸(鹰嘴豆象不含)是四纹豆象、鹰嘴豆象、西非花生豆象共有的长距离性信息素。豆象长距离性信息素组分化合物混合使用的效果高于任一单一化合物,化合物不同的比例也会导致效果不同[47]。长距离性信息素与接触性信息素结合,能提高接触性信息素的活性[31]。
在田间,可利用豆象性信息素对田间种群进行诱捕和估计,在贮藏过程中对可能造成重大危害的种群水平,利用性信息素采取预防措施,对早期种群进行破坏。但应用昆虫性信息素防治豆象依然存在一些问题,如昆虫性信息素的制剂合成、大规模生产、推广应用、使用受环境影响较大等。所以,豆象性信息素的进一步研究可以从以下方面开展:首先,利用性信息素对豆象进行诱捕需要进行田间试验,摸索害虫种群数量、诱捕面积、诱捕试剂、不同作物环境布局、诱捕设施(诱捕器设置的距离、高度、类型、颜色等)、诱捕时间、诱捕效果之间的关系,进行种群监测的诱捕器、引诱剂、引诱时间还需进一步探索和完善。其次,利用豆象的阻碍素、抗聚集信息素和植物源驱避剂联合性信息素、聚集信息素及寄主挥发物,开发有效“推—拉”防治技术,同时筛选高效植物源驱避剂和引诱剂可以有效实现豆象的绿色防控。同时,也可明确豆象性信息素的受体基因及其感受调控机制,针对重要嗅觉相关蛋白,设计并开发更高引诱活性的性信息素类似物。
参 考 文 献:
[1] 杨帅.酵母挥发物与聚集信息素调控咖啡豆象行为的研究[D].北京:中国农业科学院,2017.
[2] 闫凤鸣.化学生态学[M].2版.北京:科学出版社,2011:48.
[3] 陈新.豆象科十种仓储害虫触角感器的超微结构研究[D].长春:东北师范大学,2013.
[4] 仲建锋,万正煌,李莉,等.低温和高温对仓储绿豆象的防治效果[J].中国农业科学,2013,46(1):54-59.
[5] DE SL F R,WERMELINGER T T,DA SILVA RIBEIRO E,et al.Effects of Phaseolus vulgaris (Fabaceae) seed coat on the embryonic and larval development of the cowpea weevil Callosobruchus maculatus (Coleoptera:Bruchidae)[J].Journal of Insect Physiology,2014,60:50-57.
[6] 李庆燕,刘金龙,赵龙龙,等.缓释技术在性信息素防治害虫中的应用[J].中国生物防治学报,2012,28(4):589-593.
[7] 董双林,闫祺,高宇,等.点蜂缘蝽聚集信息素及其应用研究进展[J].南京农业大学学报,2020,43(4):583-588.
[8] BUTENANDT V A,BECKMANN R,STAMM D,et al.Uber den sexual-lockstoff des seidenspinners Bombyx mori-reindarstellung und konstituition [J].Z.Naturforsch,1959,14:283-284.
[9] HOPE J A,HORLER D F,ROWLANDS D G.A possible pheromone of the Bruchid,Acanthoscelides obtectus (Say) [J].Journal of Stored Products Research,1967,3(4):387-388.
[10] HORLER D.(-) Methyl n-Tetradeca-trans-2,4,5-trienoate8 an Allenic Ester produced by the male dried bean beetle,Acanthascelides obtectus (Say) [J].Journal of the Chemical Society,1970,1:31-34.
[11] 本田博,軒静渊.鞘翅目贮粮害虫的化学生态学及其应用[J].国外农学.植物保护,1992(2):17-19,31.
[12] 孙磊.植物精油对绿豆象控制作用的研究及有效成分鉴定[D].郑州:河南工业大学,2016.
[13] 惠琴.绿豆象产卵选择性及绿豆豆荚挥发物的田间引诱效果研究[D].晋中:山西农业大学,2019.
[14] 杨萌萌.绿豆豆荚挥发物组分鉴定及对绿豆象电生理反应和触角感器扫描电镜观察[D].晋中:山西农业大学,2016.
[15] 王广利,迟德富.天牛化学通讯及其在害虫综合治理中的应用[J].林业科学,2007(9):88-95.
[16] 朱宁.两种星天牛的行为特性和引诱剂研究[D].杭州:浙江农林大学,2017.
[17] 張时雨,张苏芳,刘福,等.双条杉天牛不同虫期体壁碳氢化合物变化规律分析[J].林业科学研究,2020,33(1):84-91.
[18] NOJIMA S,SHIMOMURA K,HONDA H.Contact sex pheromone components of the cowpea weevil, Callosobruchus maculatus [J].Journal of Chemical Ecology,2007,33(5):923-933.
[19] TANAKA K,OHSAWA K,HONDA H,et al.Copulation release pheromone,erectin,from the azuki bean weevil (Callosobruchus chinensis L.) [J].Journal of Pesticide Science,1981,82:75-82.
[20] SHIMOMURA K,MATSUI S,OHSAWA K,et al.Saltational evolution of contact sex pheromone compounds of Callosobruchus rhodesianus (Pic) [J].Chemoecology,2016,26(1):15-23.
[21] SHIMOMURA K,MATSUI S,OHSAWA K,et al.Identification of cuticular compounds collected from Callosobruchus rhodesianus (Pic) eliciting heterospecific mating behavior with male Callosobruchus maculatus (F.) [J].Chemoecology,2017,27(2):65-73.
[22] SAVKOVIC U,VUCKOVIC I,STOJKOVIC B.The growth on different stored legume species affects the profiles of cuticular hydrocarbon (CHC) in Acanthoscelides obtectus (Say) [J].Journal of Stored Products Research,2012,50:66-72.
[23] 陈光旭.立体异构化学基础[M].北京:高等教育出版社,1958.
[24] YAJIMA A,AKASAKA K.Direct determination of the stereoisomeric composition of callosobruchusic acid,the copulation release pheromone of the azuki bean weevil ,Callosobruchus chinensis L.,by the 2D-Ohrui-Akasaka Method [J].Journal of Chemical Ecology,2007,33:1328-1335.
[25] SHIMOMURA K,AKASAKA K,YAJIMA A.Contact sex pheromone components of the seed beetle,Callosobruchus analis (F.) [J].Journal of Chemical Ecology,2010,36:955-965.
[26] SHIMOMURA K,MIMURA T,ISHIKAWA S,et al.Variation in mate recognition specificities among four Callosobruchus seed beetles [J].Entomologia Experimentalis et Applicata,2010,3:315-322.
[27] QI Y T,BURKHOLDER W E.Sex pheromone biology and behavior of the cowper weevil Callosobruchus maculatus (Coleoptera:bruchidae) [J].Journal of Chemical Ecology,1982,8(2):527-534.
[28] CORK A,HALL D R,BLANEY W M,et al.Identification of a component of the female sex pheromone of Callosobruchus analis (coleoptera:Bruchidae) [J].Tetrahedron Letters,1991,142(1):129-132.
[29] RAMASWAMY S B,SHU S,MONROE W A,et al.Ultrastructure and potential role of integumentary glandular cells in adult male and female Callosobruchus subinnotatus (Pic) and Callosobruchus maculatus (Fabricius) (coleoptera:Bruchidae) [J].International Journal of Insect Morphology and Embryology,1995,24(1):51-61.
[30] PHILLIPS T W,PHILLIPS J K,WEBSTER F X,et al.Identification of sex pheromones from cowpea weevil,Callosobruchus maculatus,and related studies with Callosobruchus analis (Coleoptera:Bruchidae) [J].Journal of Chemical Ecology,1996,22(12):2233-2249.
[31] CHILUWAL K,KIM J,BAE S D,et al.Effect of gamma irradiation on fecundity,sterility,and female sex pheromone production of Callosobruchus chinensis (Coleoptera:Bruchidae) [J].Journal of Economic Entomology,2019,112(1):156-163.
[32] TUDA M,RNN J,BURANAPANICHPAN S,et al.Evolutionary diversification of the bean beetle genus Callosobruchus (coleoptera:Bruchidae):Traits associated with stored-product pest status.[J].Molecular Ecology,2006,15(12):3541-3551.
[33] VUTS J,POWERS S J,CAULFIELD J C,et al.Multiple roles of a male-specific compound in the sexual behavior of the dried bean beetle,Acanthoscelides obtectus [J].Journal of Chemical Ecology,2015,41:287-293.
[34] SHIMOMURA K,NOJIMA S,YAJIMA S.Homofarnesals:Female sex attractant pheromone components of the southern cowpea weevil, Callosobruchus chinensis [J].Journal of Chemical Ecology,2008,34:467-477.
[35] SHIMOMURA K,KOSHINO H,YAJIMA A.2,3-Dihydrohomofarnesal:Female sex attractant pheromone component of Callosobruchus rhodesianus (Pic) [J].Journal of Chemical Ecology,2010,36:824-833.
[36] PIRKLE W H,BOEDER C W.Synthesis and absolute configuration of (-)-methyl (E)-2,4,5-tetradecatrienoate,the sex attractant of the male dried bean weevil [J].The Journal of Organic Chemistry,1978,43:2091-2093.
[37] PARSONS D M J,CREDLAND P F.Determinants of oviposition in Acanthoscelides obtectus:a nonconformist bruchid [J].Physiological Entomology,2003,28(3):221-231.
[38] VUTS J,FRANCKE W,MORI K,et al.Pheromone bouquet of the dried bean beetle,Acanthoscelides obtectus (Col.:Chrysomelidae ),Now Complete [J].European Journal of Organic Chemistry,2015,22:4843-4846.
[39] MBATA G N,SHU S,RAMASWAMY S B.Sex pheromones of Callosobruchus subinnotatus and Callosobruchus maculatus( Coleoptera:Bruchidae ):congeneric responses and role of air movement [J].Bulletin of Entomological Research,2000,90(2):147-154.
[40] YOSHIO T,HIROSHI N,HAJIME S,et al.Minor components of the female sex-attractant pheromone of the smaller tea tortrix moth (Lepidoptera:Tortricidae):isolation and identification.[J].Applied Entomology & Zoology,1979,14(1):101-113.
[41] 王安佳,張开心,梅向东,等.昆虫性信息素及其类似物干扰昆虫行为的机理和应用研究进展[J].农药学学报,2018,20(4):425-438.
[42] HO H Y,TAO Y T,TSAI R S,et al.Isolation,identification,and synthesis of sex pheromone components of female tea cluster caterpillar,Andraca bipunctata walker (Lepidoptera:Bombycidae) in Taiwan [J].Journal of Chemical Ecology,1996,22(2):271-285.
[43] 胡发广,毕晓菲,付兴飞,等.天牛信息素的研究进展[J].热带生物学报,2020,11(4):517-522.
[44] 樊慧,金幼菊,李继泉,等.引诱植食性昆虫的植物挥发性信息化合物的研究进展[J].北京林业大学学报,2004(3):76-81.
[45] BOSTEDOR R G,KARKAS J D,ARISON B H,et al.Farnesol-derived dicarboxylic acids in the urine of animals treated with zaragozic acid a or with farnesol [J].Journal of Biological Chemistry,1997,272(14):9197-9203.
[46] CHILUWAL K,PARK C G,KIM J,et al.Attractiveness of male azuki bean beetle to the synthetic blends of 2E- and 2Z-homofarnesals [J].Journal of Asia-Pacific Entomology,2019,20(4):1183-1189.
[47] SHU S Q,MBATA G N,RAMASWAMY S B.Female sex pheromone in Callosobruchus subinnotatus (Coleoptera:Bruchidae):Production and male responses [J].Annals of the Entomological Society of America,1998,91(6):840-844.
[48] 劉福,袁中伟,孔祥波,等.6种松毛虫性信息素前体化合物全合成研究进展[J].林业科学,2020,56(4):170-180.
[49] COX P D.Potential for using semiochemicals to protect stored products from insect infestation [J].Journal of Stored Products Research,2004,40(1):1-25.
[50] 马涛,黄志嘉,朱映,等.尺蛾科昆虫性信息素组分特征及应用进展[J].林业科学,2019,55(5):152-162.
[51] PHILLIPS T W,THRONE J E.Biorational approaches to managing stored-product insects [J].Annual Review of Entomology,2010,35:375-397.
[52] 徐海,商胜华,袁清婷,等.3种性诱剂对烟草和白菜斜纹夜蛾的引诱效果[J].山地农业生物学报,2020,39(6):42-46.
[53] COOK S M,KHAN Z R,PICKETT J A.The use of push-pull strategies in integrated pest management.[J].Annual Review of Entomology,2007,52:375-400.
[54] PIMBERT M,BIOCDNOTIQUE I DE,CNRS U A,et al.Analyse,en olfactometre tubulaire,de l′influence de differents stimulus olfactifs dans la recherche du partenaire sexuel par Zabrotes subfasciatus [J].Entomologia Experimentalis Et Applicata,1987,45(1):23-27.
[55] PIMBERT M,POUZAT J.Electroantennogram responses of Zabrotes subfasciatus to odours of the sexual partner [J].Entomologia Experimentalis Et Applicata,1988,47(1):49-53.
[56] GINZEL M D,BLOMQUIST G J.Insect hydrocarbons:biochemistry and chemical ecology [J].Annual Review of Entomology,2016,27:149-172.
[57] BOOKSMYTHE I,RUNDLE H D,ARNQVIST G.Sexual dimorphism in epicuticular compounds despite similar sexual selection in sex role-reversed seed beetles [J].Journal of Evolutionary Biology,2017,30(11):2005-2016.
[58] SHIMOMURA K,KOSHINO H,YAJIMA A,et al.A new sesquiterpenoid produced by female Callosobruchus rhodesianus (Pic):A possible component of the sex attractant pheromone [J].Tetrahedron Letters,2010,51(52):6860-6862.
