两个产孢梗束的节肢动物病原真菌新种
2021-12-21邹晓韩燕峰梁宗琦瞿娇娇
邹晓 韩燕峰 梁宗琦 瞿娇娇

摘 要:本文报道了来自中国贵州省两个产孢梗束的虫生真菌新种。一个为虫草属1新种,新莲蓬虫草Cordyceps neonelumboides sp.nov.。该种寄生于蜘蛛体上,可孕部莲座状,成熟子囊孢子断裂成次生孢子,分隔细胞长15.3~18.0 μm。其无性型为刺束梗孢属,瓶梗指状,5.4~7.2 μm×0.8~1.0 μm;分生孢子长柱状,两端略尖,12.6~13.5 μm×1.5~1.8 μm,形成长的分生孢子链。另一个为被毛孢属1新种,毛虫被毛孢Hirsutella campes sp.nov.。该种显著特征为菌丝具隔,产孢细胞细长,锥形,或基部圆柱状,21.6~38.4 μm×2.4~4.8 μm,向上逐渐变细,宽0.9~1.5 μm,分生孢子单孢或多孢,橘瓣状、梭形,6.0~8.4 μm×2.9~4.3 μm。文中讨论了两个新种与相近种之间的区别以及系统发育关系。
关键词:虫草属;被毛孢属;形态特征;系统发育;分类
中图分类号:Q939.5
文献标识码:A
文章编号:1008-0457(2021)06-0001-10
国际DOI编码:10.15958/j.cnki.sdnyswxb.2021.06.001
Abstract:This paper reported two new species of entomopathogenic fungi producing synnema in Guizhou Province,China.The first new species of entomopathogenic fungi was Cordyceps neonelumboides sp.nov.This fungus was parasitic on the spider body,the stroma was lotus-like,the mature ascospores were broken into secondary spores,and the length of separated cells was 15.3~18.0 μm.The anamorph was the genus Akanthomyces,and the conidiogenous cell was 5.4~7.2 μm×0.8~1.0 μm,cone-shape.The conidia were long cylindrical,slightly pointed at both ends and its size was 12.6~13.5 μm×1.5~1.8 μm.The other new species was the genus Hirsutella,H.campes sp.nov.The distinctive feature of this species was that the hyphae were septal,the spore-forming cells were slender,tapered,or cylindrical at the base,21.6~38.4 μm×2.4~4.8 μm.The spore-forming cell was gradually thinning upwards,and the width of cell was 0.9~1.5 μm.The conidia were monospores or polyspores,orange petal-like,fusiform,6.0~8.4 μm×2.9~4.3 μm.This research also discussed the phylogenetic relationship between two new species and similar species.
Keywords:Cordyceps;Hirsutella;morphological characteristics;phylogenic analysis;taxonomy
蟲生真菌(entomopathogenic fungi)是一类能够寄生昆虫、蜘蛛等节肢动物并可导致寄主死亡的真菌,是自然界中节肢动物和线虫等种群的重要生物调控因子[1]。自然界中虫生真菌资源丰富,种类超过1000多种,
分布于壶菌门、子囊菌门、接合菌门、担子菌门、半知菌门等各个亚门中,其中得到广泛研究与应用的主要是子囊菌门类真菌[2]。随着人类生活水平的提高和生物技术的发展,虫生真菌被广泛应用于食品、医药、生防等多个方面。越来越多的研究发现,部分虫生真菌(蛹虫草、冬虫夏草、蝉花等)具有抗菌、抗病毒、抗肿瘤、抗疲劳、抗衰老、降血脂血糖、提高人体免疫力等药用功效[3-6]。因此为满足人们对具有药用价值和保健作用的虫生真菌资源日益增加的需求,近年来国家陆续批准蛹虫草、蝉花(蝉棒束孢)作为新资源食品。一直以来,虫生真菌研究则更多关注微生物杀虫剂对农林经济害虫的控制。如以白僵菌等广谱性虫生真菌为模式,在致病相关基因的作用与功能、真菌逃避昆虫免疫的分子机理、真菌识别宿主与寄主专化性形成的机理、真菌有性生殖调控、活性成分合成代谢机理、真菌抗逆机理以及遗传改造提高虫生真菌的应用效率等方面展开研究[2]。除了杀虫,研究还发现虫生真菌具有多重生态价值,可与植物进行元素交换,能够与植物建立共生体系并具有提高作物产量、抵抗植物病害等生态功能[7]。因此,开展虫生真菌种类资源及分布研究,对发现新资源、开发新的生物制品及筛选高效生防菌具有十分重要的理论意义和应用价值。
虫草属[Cordyceps(Fr.)Link]真菌是寄生在昆虫躯体直至导致昆虫死亡的一大类子囊菌;有时也寄生蜘蛛,甚至某些大团囊菌地下子实体上;现隶属于虫草菌科(Cordycipitaceae)、肉座菌目(Hypocreales)、子囊菌纲(Ascomycetes),是虫草菌科的模式属[8]。虫草属约有564种(Index Fungorum 2015),曾于2007年正式被迁移到拟虫草科(Cordycipitoideae)[9]。Sung 等[9] 将虫草属中的一些种转移至Ophiocordyceps (Ophiocordycipitaceae)、Elaphocordyceps (目前名为Tolypocladium, Ophiocordycipitaceae)、Metacordyceps (Clavicipitaceae)、Claviceps (Clavicipitaceae)、Tyrannicordyceps (Clavicipitaceae),Epichloe (Clavicipitaceae)、Podostroma (Hypocreaceae)以及Podocrea (Hypocreaceae)等几个属中。目前虫草属中包含180个形态学种(Index Fungorum,2021),42个种具有分子序列数据[8]。该属真菌能感染昆虫,产生明亮、具柄、肉质的子座,子囊壳表生或埋生,子囊孢子可断裂或不断裂成次生孢子;该属的无性型能产生类似棒束孢、蜡蚧、拟青霉状的瓶梗或分生孢子[10]。
Mains[11]列出了8种已知的寄生在蜘蛛上的虫草属种。随后Kobayasi[12],Hywel-Jones[13]和Aung等[14] 报道了一些寄生在蜘蛛上的虫草属种。目前为止,寄生在蜘蛛上的虫草有16种,分别为:C.arachneicola Kobayasi,C.arachnogena Kobayasi,C.caloceroides Berk.& M.A.Curtis,C.coccidioperitheciata Kobayasi & Shimizu,C.cylindrica Petch,C.engleriana Henn.,C.grenadensis Mains,C.ignota Marchion,C.mrciensis Aung,J.C.Kang,Z.Q.Liang,Soytong & K.D.Hyde,C.nelumboides Kobayasi & Shimizu,C.ogurasanensis Kobayasi & Shimizu,C.pseudonelumboides Kobayasi & Shimizu,C.singeri Mains,C.thaxteri Mains ,C.wittii Henn 和 C.kuiburiensis Himaman,Mongkols.,Noisrip.& Luangsa-ard (Index Fungorum,2021)。
被毛孢属(Hirsutella Pat.)是由Patouillard[15]根据模式种H.entomophilila建立的,隶属于线虫草菌科(Ophiocordycipitaceae)、肉座菌目(Hypocreales)、子囊菌纲(Ascomycetes)[8]。该属真菌是无性型属,其有性型与Ophiocordyceps属相关联,是昆虫病原真菌中种类较多和意义较大的类群之一。被毛孢种能寄生多种鳞翅目昆虫的幼虫和蛹,此外还寄生鞘翅目的昆虫、叶蝉、飞虱、蚂蚁、蚜虫、介壳虫、蝗虫、椿象、黄蜂、蟋蟀、蝇、蜘蛛以及螨类和线虫[16]。除少数种外,被毛孢对寄主有一定的专一性。严格意义上的被毛孢属真菌的形态特征为:寄主为节肢动物或线虫;孢梗束直立、柱状或者纤细、向上逐渐变尖;产孢细胞分散或聚集,从孢梗束的外层菌丝上发出,基部膨大(瓶形或近球形或锥形),颈部伸长变细,有时颈部会出现多个短的小颈;分生孢子近柱状、圆形、近似梭形或橘瓣状,外具黏液或者2~3个孢子聚集在一个液滴内。孢梗束颜色从灰白色、棕色到深褐色[17-21]。
被毛孢属自建立以来,历经分类地位变更和分类特征的变化,该属真菌分类趋于科学且种类不断增加。随着“一种真菌一个名称”新命名规则的实施,被毛孢属一度被建议移除,而将其归于线虫草属Ophiocordyceps中[9]。因此,随后一些仅具有被毛孢无性型形态的新种也被置于Ophiocordyceps属中进行发表,如O.myrmicarum sp.nov.[22],O.retorta sp.nov.[23]。近年来,Ophiocordycipitaceae的分类学研究在真菌新规则下进展迅速。根据最新的分类学报道,被毛孢属(Hirsutella)作为独立的属重新被置于Ophiocordycipitaceae中[8]。因此,今后具有被毛孢无性型形态新种的发表仍然要将其归为被毛孢属,如我们最近发表的新种Hirsutella flava sp.nov.和H.kuankuoshuiensis sp.nov.[17]。目前被毛孢属中包含80个形态学种(Index Fungorum,2021)。
在对贵州省虫生真菌资源调查过程中,采集到两个分别寄生于蜘蛛和鳞翅目幼虫的真菌标本,均能产生孢梗束,分离获得了对应的无性型菌株并对其进行形态学鉴定和基于多基因的系统发育分析。结果分别鉴定为虫草属和被毛孢属中的一个新成员,增加了子囊菌类真菌的物种资源。
1 材料与方法
1.1 材料
1.1.1 标本
标本1采自贵州省习水县习水国家级自然保护区(N28°07′34″,E105°50′29″),寄主为节肢动物蛛形纲(Arthropoda,Arachnida)一种蜘蛛。标本编号为GZUIFR-XS4692,采集时间为2012年8月,采集人:邹晓,菌株编号为GZUIFR-xishui-4692。
标本2采自贵州省宽阔水国家级自然保护区(N28°06′25″,E107°02′23″),寄主为鳞翅目(Lepidoptera)一种毛虫。标本编号为GZUIFR-2010MC,采集时间为2010年7月,采集人:邹晓,菌株编号为GZUIFR-2010MC-1,GZUIFR-2010MC-2 和 GZUI FR-2010MC-3。
上述标本与菌株均保存于贵州大学真菌资源研究所(Institute of Fungal Resources Collection,Guizhou University,GZAC)。
1.1.2 培养基及主要试剂和仪器
PDA培养基[13];葡萄糖、琼脂、苯胺蓝、乳酸、石炭酸、甘油及样本脱水试剂均为分析纯,购自国药集团化学试剂有限公司;真菌DNA提取试剂盒OMEGAFungal DNA kit,购自OMEGA Bio-Tek公司;光学显微镜BK5000,购自OPTEC公司;掃描电镜S-3400N,购自HITACHI公司。
1.1.3 染色液
棉蓝染色液:苯胺蓝0.025 g,乳酸10.000 g,石炭酸10.000 g,甘油20.000 g,蒸馏水10 mL。配制完后用双层擦镜纸过滤后备用。
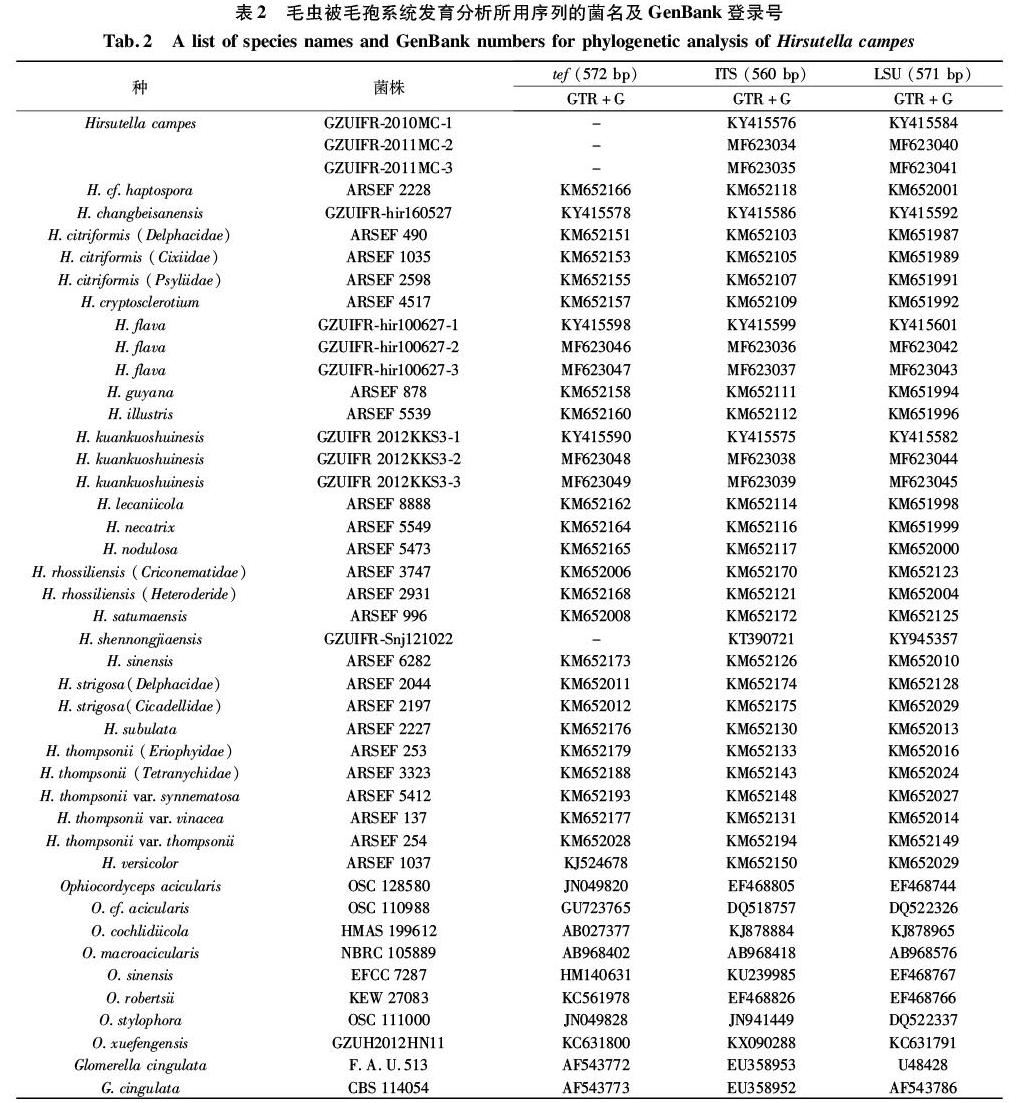
1.1.4 分子系统学分析所用序列
详见表1和表2。
1.2 方法
1.2.1 菌种分离、培养与形态学鉴定
参照Qu等[17]的方法,用无菌水冲洗标本表面,然后用75%乙醇表面消毒3~5 s。将虫体掰断,露出虫体腹部,用无菌镊子挑取虫体断面的菌核接种到PDA平板上,22 ℃培养30 d。光学显微镜观察:用透明胶带黏取菌落边缘具产孢结构的菌丝制片,经棉兰染液染色3 min左右,置于光学显微镜下观察,使用软件Paint Shop Pro v.5.0.1 (Corel,Ottawa,Canada)記录菌种产孢结构信息。扫描电镜观察:参照Qu等[17]的方法制样,在PDA培养基上取1 cm2的菌株培养物,于4%戊二醛溶液中4 ℃浸泡过夜,用磷酸盐缓冲液 (137 mmol/L NaCl,2.7 mmol/L KCl,8.1 mmol/L Na2HPO4,1.5 mmol/L KH2PO4,pH 7.4)冲洗3次,每次10 min;分别用30%、50%、70%、90%、95%、100%乙醇进行逐级脱水,每级10 min;脱水后使用超临界CO2干燥样品,最后喷金于扫描电镜下进行观察拍照。
1.2.2 DNA的提取,PCR扩增及测序
从PDA平板上刮取菌丝0.05~0.10 g用于DNA的提取,采用真菌DNA提取试剂盒提取样本DNA。随后以真菌基因组DNA为模板,选用各引物对进行多基因序列的扩增,引物为:(1)983F和2218R [10]用于tef1扩增;(2)LROR和LR5 [24]用于28S rRNA基因扩增;(3)ITS1和ITS2[25]用于ITS rDNA扩增。PCR产物送由上海Invitrogen公司纯化和测序。
1.2.3 基于多基因序列的系统发育分析
测序结果经 BioEdit 软件分析和手工校正,经Sequin软件编辑后提交GenBank获取登录号;菌株GZUIFR-xishui-4692的ITS序列经BLAST后下载相似性大于84%的相关近缘属菌株的相应序列(表1),标本GZUIFR-2010MC相应菌株的tef1、ITS和28S-rDNA序列经BLAST比对,下载相似性大于84%的近缘属种的这3个基因序列(表2);序列进行多重比对并使用SequenceMatrix 1.7.8软件拼接,通过jModeltest 2.1.7运行合适的核苷酸置换模型[26];在RAxML中使用推荐的参数构建最大似然系统发育树[27],用MrBayes 3.1.2重建系统发育树,运行50万代后舍弃25%的样本[17];最后使用Figtree 1.4.2编辑发育树图。
2 结果与分析
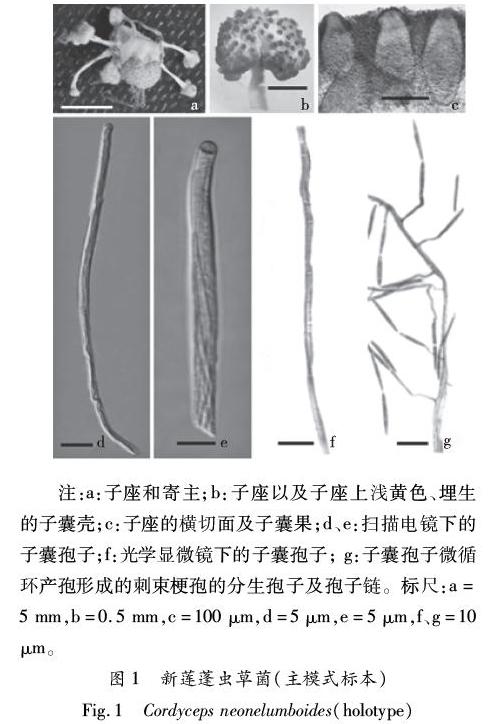
2.1 新莲蓬虫草 新种 图1
Cordyceps neonelumboides X.Zou,J.J Qu,Z.Q.Liang and Y.F.Han sp.nov.Figure 1
子座从寄主背部长出,浅黄色,散生,不分枝。子座柄柱状,肉质,白色,3~5 mm×0.3mm。可孕部莲座状,直径1.1 mm ~1.3mm,厚0.8 mm ~1mm(图1-a、b)。子囊壳垂直埋生,拟椭圆形,675 μm×195~205 μm(图1-c)。子囊柱状,485~575 μm×3.6~5.4 μm;子囊帽宽3.6 μm ~5.5 μm,高2.7 μm ~3.1 μm。子囊孢子丝状,多隔,510~650 μm×1~1.3 μm,断裂形成次级孢子,分隔细胞长15.3 μm ~18 μm(图1-d、e、f)。
无性型:Akanthomyces neonelumboides。瓶梗和分生孢子直接通过子囊孢子微循环产孢形成。瓶梗指状,长5.4 μm ~7.2 μm,宽0.8 μm ~1.0 μm。分生孢子长柱状,两端略尖,12.6~13.5 μm×1.5~1.8 μm,形成长的分生孢子链(图1-g)。
寄主:节肢动物,蛛形纲,蜘蛛。
模式标本:GZUIFR-XS4692标本及其分离株GZUIFR-xishui-4692。
语义学:源于标本子座的形状。
MycoBank:MB812302;Facesoffungi number:FoF 00646;GenBank:MZ926848。
2.2 毛虫被毛孢 新种 图2
Hirsutella campes X.Zou,J.J.Qu,Z.Q.Liang and Y.F.Han sp.nov.Figure 2
孢梗束从毛虫头部伸出,细长、褐色至顶部淡褐色;12~15 cm×0.5~1.0 mm (图2-a)。该菌在PDA培养基上生长缓慢, 20~22 ℃下12 d菌落直径为1.3 cm ~1.8 cm;菌落边缘绒毛状,中部淡黄色凸起,背面分泌深褐色色素,扩散圈较大,1.0 cm ~1.5 cm (图2-b、c)。菌丝光滑或具疣,无色,具隔,宽1.8 μm ~3.6 μm。产孢细胞瓶梗状,大部分垂直于菌丝,少部分由菌丝顶端直接形成,细长,21.6 μm ~38.4 μm,锥形,或基部圆柱状,宽2.4 μm ~4.8 μm,向上逐渐变细,宽0.9 μm ~1.5 μm (图2-d、e、h、i)。分生孢子长梭形或橘瓣状,6~8.4 μm×2.9~4.3 μm,光滑,无色,单生,偶有两个或多个形成孢子团包裹于黏液鞘中(图2-f、g、j)。
有性阶段:虫草。
寄主:鳞翅目,毛虫。
模式标本:GZUIFR-2010MC标本及其分离株GZUIFR-2010MC-1,GZUIFR-2010MC-2 和 GZUIFR-2010MC-3。
语义学:源于寄主毛虫,拉丁语campes。
MycoBank:MB819707; GenBank:KY4155 76,KY415584。
2.3 系统发育分析
基于ITS序列,以Tolypocladium inflatum(OSC 71235)、T.ophioglossoides (NBRC106332)为外群,采用贝叶斯推理法和最大似然法(ML)构建了菌株GZUIFR-xishui-4692与虫草属其他种的系统发育树,如图3所示。在进化树中,新莲蓬虫草与Akanthomyces araneicola 关系密切并聚集在一起,Bootstrap值为90/92。这一分支与Gibellula属以及Akanthomyces的几个种聚在一类,说明新莲蓬虫草与Akanthomyces联系紧密,这与形态学结果一致。
另外,基于tef1,ITS 和 28S rDNA序列,以Glomerella cingulata(F.A.U.513和CBS 114054)为外群,采用贝叶斯推理法和最大似然法(ML)构建了菌株GZUIFR-2010MC与被毛孢属其他种的系统发育树,如图4所示。所用供试序列明显分为6个进化分支,分别为:H.thompsonii,H.citriformis,H.sinensis,H.guyana,H.nodulosa和H.flava,这与之前报道的结果一致[22,23]。在进化树中,毛虫被毛孢与H.strigose Petch关系密切并聚集在H.sinensis 分支中,Bootstrap值为89/92。这一类群的被毛孢形态学上的共同特征是瓶梗尺寸较大,有的种甚至超过100 μm,基部呈圆柱形,细长或锥形,逐渐变细,颈部有疣状突起、通常大于30 μm,因此系统发育分析与形态学结构相对应。但毛虫被毛孢具锥形瓶梗,颈部细长但比H.strigosa短,与H.sinensis分支其他种具有明显差别。
分子系统发育分析进一步印证了新莲蓬虫草、毛虫被毛孢与其他物种的区别与联系,因而将这两个种作为新成员分别引入虫草属和被毛孢属。
3 结论与讨论
分子系统发育分析在微生物分类方面已形成一种主流,快速、简便、省时、客观的特点恰好弥补了传统形态学分类的不足,但受基因库完整性、序列保守度及同源性的限制,而不能准确地鉴定出所有的物种,因此形态学和分子系统发育相结合的分类方法,很大程度上节约了鉴定时间,并提高了分类鉴定的正确率[39]。在真菌分类方面,ITS是识别大多数真菌成功率最高的序列[40],其可以准确鉴定到真菌种级水平,已得到广泛应用。在束梗孢类分类方面,ITS rDNA依然是很重要的分类参考指标[41]。在本研究中,从新莲蓬虫草的分离菌株GZUIFR-xishui-4692的基因组中成功扩增出ITS序列,而tef1和28S rRNA基因未能扩增出来,这可能是因为使用的通用引物不适合该物种。但在束梗孢类真菌的研究中,如Akanthomyces、Hirsutella、Hymenostilbe等中的一些种除ITS序列外的保守基因序列的扩增比较困难[42],因此ITS rDNA在这类种的鉴定和识别上具有重要的实用价值。
虫草属中寄主为蜘蛛的种有多个,与本研究的新莲蓬虫草(C.neonelumboides)标本宏观形态上较为接近的是拟莲状虫草(C.pseudonelumboides)[43]和莲状虫草(C.nelumboides)[44]。与C.pseudonelumboides最大的区别在于子囊壳的埋生方式,拟莲状虫草为半埋生,而本标本的埋生方式为全埋生,且二者在子囊壳宽度上也不同(表3)。C.neonelumboides主要特征为子囊孢子断裂产生次生孢子,且子囊孢子长度为15.3~18 μm×1~1.3 μm,比C.nelumboides和C.pseudonelumboides要大得多。此外,C.nelumboides的无性型为Hirsutella,而C.neonelumboides的无性型为Akanthomyces,两个种区分较明显。在系统发育分析中,本种与Akanthomyces araneicola关系密切独立聚为一支,但形态学上两者区别较大。A.araneicola的标本没有形成子座,瓶梗基部膨大,分生孢子梭状,2.5~5.0 μm×1.3~1.9 μm;但本種无性型产生的瓶梗为指状,分生孢子长圆柱状,12.6~13.5 μm×1.5~1.8 μm。因此,形态学和系统发育分析结果显示本研究标本GZUIFR-XS4692为虫草属1新种Cordyceps neonelumboides sp.nov.。
在本研究中多基因系统发育树分析结果表明(图4),标本GZUIFR-2010MC分离菌株与其他种有明显的区别,聚在H.sinensis这一分支,形态特征都具有膨大基部和尺寸较大的瓶梗。本研究种与H.strigose Petch聚在一起,表明二者遗传距离较近,关系较为亲密。但形态特征方面,H.strigose产孢细胞较长,36~90 μm,具小梗和瘤状结构,且其分生孢子具隔[31,45],这些特征与本文研究种区别明显。本研究标本与被毛孢属中其他近似种的形态学比较见表4。形态学分析和ITS1-5.8S-ITS2 rDNA序列系统发育分析都支持该研究标本为被毛孢属1新种Hirsutella campes sp.nov.。
参 考 文 献:
[1] EVANS H C.Natural control of arthropods,with special reference to ants (Formicidae),by fungi in the tropical high forest of Ghana[J].Journal of Applied Ecology,1974:37-49.
[2] 王成樹.虫生真菌研究与应用的发展与机遇——2012年虫生真菌专刊序言[J].菌物学报,2012,31(3):305-306.
[3] 钱正明,李文庆,孙敏甜,等.冬虫夏草化学成分分析[J].菌物学报,2016,35(4):476-490.
[4] 梁宗琦,韩燕峰,梁建东,等.冬虫夏草Ophiocordyceps sinensis研究中几个值得关注的问题[J].微生物学通报,2010,37(11):1692-1697.
[5] WANG M,MENG X Y,LE Y R,et al.Cordyceps militaris polysaccharides can enhance the immunity and antioxidation activity in immunosuppressed mice[J].Carbohydrate Polymers,2012,89(2):461-466.
[6] FATMA E L,ZAHRAA Z,MAHMOUD M F,et al.Effect of Cordyceps sinensis and taurine either alone or in combination on streptozotocin induced diabetes[J].Food and Chemical Toxicology,2012,50(3):1159-1165.
[7] 邹晓.虫生真菌环境适应研究简述[J].山地农业生物学报,2020,39(6):1-7.
[8] HYDE K D,NORPHANPHOUN C,MAHARACHCHIKUMBURA S S N,et al.Refined families of sordariomycetes[J].Mycosphere,2020,11(1):305-1059.
[9] SUNG G H,HYWEL-JONES N L,SUNG J M,et al.Phylogenetic classification of Cordyceps and the clavicipitaceous fungi[J].Studies in Mycology,2007,57:50-59.
[10] SUNG G H,SUNG J M,HYWEL-JONES N L,et al.A multi-gene phylogeny of clavicipitaceae(Ascomycota,Fungi):identification of localized incongruence using a combinational bootstrap approach[J].Molecular Phylogenetics and Evolution,2007,44(3):1204-1223.
[11] MAINS E B.Species of Cordyceps on spiders[J].Bulletin of the Torrey Botanical Club,1954,81:492-500.
[12] KOBAYASI Y.Keys to the taxa of the genera Cordyceps and Torrubiella[J].Transactions of the Mycological Society of Japan,1982,23:329-364.
[13] HYWEL-JONES N L.The biological diversity of invertebrate pathogenic fungi.In "Biodiversity of Tropical Microfungi"ed.K.D.Hyde[M].Hong Kong:Hong Kong University Press,2001:107-120.
[14] AUNG O M,KANG J C,LIANG Z Q,et al.Cordyceps mrciensis sp.nov.from a spider in Thailand[J].Mycotaxon,2006,97:235-240.
[15] PATOUILLARD N.Une Clavariée entomogène[J].Revue de Mycologie,1892,3:67-70.
[16] VAN DER GEEST L P S.IPM potentials of microbial pathogens and diseases of mites[M].In:Integrated management of arthropod pests and insect borne diseases,Ciancio A,Mukerji KG (Eds)Springer,Dordrecht,2010:249-309.
[17] QU J J,ZOU X,CAO W,et al.Two new species of Hirsutella (Ophiocordycipitaceae,Sordariomycetes)that are parasitic on lepidopteran insects from China[J].MycoKeys,2021,82:81-96.
[18] CORY J S,WILSON K R,HAILS R S,et al.Endocrine interactions of insect parasites and pathogens (Edwards,J P and WeaverR J eds)[M].Oxford:BIOS Scientific Publishers,2001:233-244.
[19] THUMMEL C S.Molecular mechanisms of developmental timing in C.elegans and Drosophila[J].Developmental Cell,2001,1(4):453-465.
[20] HALL D G,HENTZ M G,MEYER J M,et al.Observations on the entomopathogenic fungus Hirsutella citriformis attacking adult Diaphorinacitri (Hemiptera:Psyllidae)in a managed citrus grove[J].BioControl,2012,57(5):663-675.
[21] 董彩虹,李文佳,李增智,等我國虫草产业发展现状,问题及展望——虫草产业发展金湖宣言[J].菌物学报,2016,35(1):1-15.
[22] SIMMONS D R,LUND J,LEVITSKY T,et al.Ophiocordyceps myrmicarum,a new species infecting invasive Myrmica rubrainMaine[J].Journal of Invertebrate Pathology,2015,125:23-30.
[23] QU J J,ZHOU Y M,YU J P,et al.Estimated divergence times of Hirsutella (asexual morphs)in Ophiocordyceps provides insight into evolution of phialide structure[J].BMC Evolutionary Biology,2018,18:111.
[24] REHNER S A,SAMUELS G J.Taxonomy and phylogeny of Gliocladium analysed from nuclear large subunit ribosomal DNA sequences[J].Mycological Research,1994,98:625-634.
[25] WHITE T J,BRUNS T D,LEE S B,et al.Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics[M].In:Innis MA,Gelfand DH,Sninsky JJ,White TJ (Eds)PCR Protocols:a guide to methods and applications,San Diego:Academic Press,1990:315-322.
[26] GUINDON S,GASCUEL O.A simple,fast,and accurate algorithm to estimate large phylogenies by maximum likelihood[J].Systematic Biology,2003,52:696-704.
[27] STAMATAKIS A,HOOVER P,ROUGEMONT J.A rapid bootstrap algorithm for the RAxML Web-Servers[J].Systematic Biology,2008,75:758-771.
[28] KOBAYASI Y.The genus cordyceps and its allies[J].Tokyo Bunrika Daigaku,Science Report,1941,5(84):53-260.
[29] PETCH T.Studies in entomogenous fungi Ⅳ.Some Ceylon Cordyceps[J].Transactions of the British Mycological Society,1924,10:28-45.
[30] KOBAYASI Y,SHIMIZU,D.Some species of Cordyceps and its alles on spiders[J].Kew Bulletin,1977,31:557-566.
[31] PETCH T.Notes on entomogenous fungi[J].Transactions of the British Mycological Society,1939,23:127-148.
[32] SPEARE A T.On certain entomogenous fungi[J].Mycologia,1920,12:62-76.
[33] BAAZY S,MITKIEWSKI R ,TKACZUK C ,et al.Diversity of acaropathogenic fungi in Poland and other European countries[J].Springer Netherlands,2008,46:53-70.
[34] LIU X J,GUO Y L,YU Y X,et al.Isolation and identification of the anamorphic state of Cordyceps sinensis (Berk.)Sacc[J].Mycosystema,1989,8:35-40.
[35] MINTER D W,BRADY B L.Mononematous species of Hirsutella[J].Transactions of the British Mycological Society,1980,74:271-282.
[36] PETCH T.Notes on entomogenous fungi[J].Transactions of the British Mycological Society,1937,21:34-67.
[37] EVANS H C,SAMSON R A.Cordyceps species and their anamorphs pathogenic on ants (Formicidae)in tropical forest ecosystems I.The Cephalotes (Myrmicinae)complex[J].Transactions of the British Mycological Society,1982,79:431-453.
[38] FISHER F E.Two new species of Hirsutella Patouillard[J].Mycologia,1950,42(2):290-297.
[39] 燕勇,李卫平,高雯洁,等.rDNA-ITS 序列分析在真菌鉴定中的应用[J].中国卫生检验杂志,2008,18(10):1958-1961.
[40] SCHOCH C L,SEIFERT K A,HUHNDORF S,et al.Nuclear ribosomal internal transcribed spacer (ITS)regionas auniversal DNA barcode marker for fungi[J].Proceedings of the National Academy of Sciences,2011,109:6241-6246.
[41] 張延威,陈万浩,邹晓,等.来自山西鸟窝的金孢属一新种[J].菌物学报,2016,35(11):1337-1343.
[42] 邹晓,周叶鸣,梁宗琦,等.瓶梗颈部螺旋且寄生卷叶蛾幼虫的被毛孢新种[J].菌物学报,2016,35(7):807-813.
[43] KOBAYASI Y,SHIMIZU D.Cordyceps species from Japan 4[J].Bulletin of the National Science Museum Tokyo,1982,8(3):79-91.
[44] KOBAYASI Y,SHIMIZU D.Some species of Cordyceps and its allies on spiders[J].Kew Bulletin,1977,31(3):557-566.
[45] HODGE K T.Revisionary Studies in Hirsutella (anamorphic Hypocreales:Clavicipitaceae)[microform][M].Ann Arbor,Mich:University Microfilms International,1998:144-146.
