听觉响应信噪比在上行听觉通路中的演变
2021-12-16王敬敬宋长宝梁妃学
王敬敬,宋长宝,梁妃学
南方医科大学生物医学工程学院,广东 广州 510515
把声音特征转化为动作电位编码信息,并根据获取的声音信息含义做出感觉或行为响应是听觉神经系统的核心任务[1,2]。听觉神经系统为完成这一任务动用了由多个核团组成的庞大听觉信息处理网络,其中下丘(IC)、内侧膝状体(MGB)和初级听皮层(A1)处于听觉信息处理网络的核心地位[3,4],因此充分研究这些核团的听觉信息处理机制尤其重要。研究发现听觉系统中的每个脑区都有大量神经元对声音信息的不同特征进行编码[5,6],Pickles等[7]系统阐述了上行听觉通路核心脑区的解剖结构及生理特性,表明声音特征的编码主要体现在神经元动作电位的发放;Yildiz等[6]利用数学模型来评估神经元的编码特性,发现神经元听觉感受野符合非线性模型,该模型只能反映单个神经元对不同声音的响应但不能反映对声音信息的敏感特性;Zador 等[5]报道了听皮层神经元的编码既有密集性又有稀疏性,同时Zador用自发与诱发来表示听皮层神经元对声音刺激的响应,自发与诱发的大小可表示神经元对声音信息的敏感特性。而先前的研究往往聚焦于单一脑区的处理机制,声音特征的编码规律在听觉核团中如何演变目前所知甚少,因此本研究用诱发与自发的比值(信噪比,SNR)来表征神经元对特定声音刺激的编码强度,探究上行听觉通路中神经元编码的规律。
在生理状态下,感觉神经系统对外界的刺激响应受到大脑状态的调节。有研究指出[8-11]当小鼠处于运动状态时,给予小鼠视觉刺激,视觉皮层的响应增加而内侧膝状体的响应不变;与视觉皮层相反[12],听觉层的响应在运动状态下减少;Zador等[13]表明大鼠在参与任务时,听皮层神经元的诱发被抑制而内侧膝状体神经元的诱发没有改变。上述现象暗示,听觉响应SNR可能受到大脑状态的影响。因此,不同脑区的听觉响应SNR在不同大脑状态中是否维持稳定还需要进一步研究,本研究通过分析不同运动状态下小鼠IC、MGB、A1 脑区SNR的变化来对其稳定性进行探究。
为了了解听觉响应SNR在上行听觉通路中的演变规律及在不同大脑状态下是否维持稳定,本研究利用在体细胞贴附式记录方法记录小鼠在安静和跑动状态下不同听觉脑区的听觉响应,研究最佳频率下的听觉响应SNR在不同脑区及不同大脑状态下的变化规律。我们的结果显示不同脑区的诱发与自发没有相关性,自发与诱发在群体上的分布分别符合泊松分布与对数正态分布,多个脑区的SNR在不同大脑状态下维持稳定。我们的结果为深入理解不同脑区的神经元编码声音特征提供重要的理论基础。
1 材料和方法
1.1 动物
SPF级C57BL/6J小鼠共80只,小鼠周龄6~8周,体质量18~20 g,雌雄不限,购自南方医科大学实验动物中心(No.44002100018990)。所有动物经检查听觉能力良好,鼓膜经检耳镜检查无异常。所有实验均在南方医科大学实验动物福利与伦理委员会的批准和监督下进行(L2017207)。
1.2 手术准备
选取C57BL/6J小鼠1只,小鼠腹腔注射戊巴比妥钠(60 mg/kg)进行麻醉,并使用温控系统使动物体温维持在37 ℃,麻醉完全后将小鼠转置于脑立体定位仪上,在立体显微镜下依据小鼠脑图谱(第二版)分别确定下丘、内侧膝状体以及听皮层的脑区位置并进行位置标记。用牙科水泥将一个长约1.5 cm的平头金属钉粘合在颅骨前部正中位置,待小鼠头部牙科水泥完全凝固之后,再将小鼠固定到屏蔽室的实验平台上,在体视显微镜下用颅骨钻去除下丘、内侧膝状体以及听皮层上方的颅骨,充分暴露出相应的脑组织,将暴露在外的组织覆盖一层凡士林塑料膜以保护脑组织及创口。手术完成后,小鼠放回鼠笼恢复2 d,以备后续进行电生理实验。记录开始前将恢复好的小鼠头部固定在金属杆上,身体放置在可旋转的转盘中进行训练,直到小鼠能够在转盘上进行自由跑动或静止,将训练好的老鼠转移到屏蔽试验台进行听觉电生理记录。在手术和记录过程中,暴露的皮质由恒温人工脑脊液(ASCF:Nacl,132 mmol/L;NaHCO3,20 mmol/L;ΚCl,2.5 mmol/L;NaH2PO4,1.2 mmol/L;MgSO4,1.1 mmol/L;CaCl2,2 mmol/L;Hepes,3 mmol/L;glucose,15 mmol/L)保持湿润。
1.3 声音刺激
本研究采用美国TDT给声系统产生刺激声音。利用Labview软件编辑合成声音信号并通过数据采集卡(NI,6731)将声音信息输出到声音驱动系统(即TDT系统中喇叭驱动器(ED1)),驱动系统驱动放置在屏蔽室中的喇叭(ES1)完成发声目的。研究中所有的刺激声音均由精密的声音校正仪进行严格的校准。
1.4 刺激参数
本研究给予小鼠的刺激声音是由软件合成的一系列纯音,由8个声强级(0~70 dB,以10 dB递增),51个频率(2~64 ΚHz,0.1 octave递增)组成,可以判断记录到的听觉神经元的特征频率(CF)或最佳频率(BF)。在记录过程中,记录信号的采样率为10 ΚHz/s,每个刺激声音时长50 ms,记录时长为500 ms,在记录长度为50 ms时设置声音刺激开始,两个刺激声音之间的间隔为500 ms,在设置好记录参数之后开始记录。
1.5 电生理记录
电生理实验在听觉屏蔽室中进行。在电生理记录过程中,用平头金属钉将训练好的老鼠头部固定,在显微镜下去除记录脑区的凡士林覆盖膜,在拉制好的玻璃微电极内灌注含0.3%神经生物素的人工脑脊液(ACSF)并排出电极尖端气泡,玻璃电极稳定安装在前置放大器探头上后将玻璃微电极置于记录脑区正上方,通过压力表给予电极2个正压,滴加一滴人工脑脊液在脑区表面,利用电动显微操纵器使电极与液体充分接触,此时测量电极电阻显示为6 MΩ左右,说明小鼠与放大器连通,可进行数据采集实验。电极快速移动100 μm进入记录脑区表层,此时将正压减半,用推进器缓慢移动玻璃电极至所需脑区深度寻找听觉神经元。与此同时,通过观察Labview软件中显示的电极电阻是否增大来判断电极是否碰触到细胞,当观察到电极电阻忽然增大2倍,观察到有声诱发动作电位,此时撤掉电极正压,待电阻稳定不变后给予适当的负压吸引使神经元紧密贴附在电极上,最后给予小鼠合成的纯音刺激,记录该神经元的响应数据(图1A)。利用Matlab软件读出该神经元的感受野,判断该神经元的最佳频率(BF),给予小鼠声强60 dB,频率为BF的纯音刺激,记录最佳频率下神经元的听觉响应数据,神经元的听觉刺激响应如图1B。利用该记录方法分别寻找下丘、内侧膝状体和初级听皮层三个脑区的听觉神经元。声音刺激期间对应的时间窗口动作电位发放数为听觉诱发放电,记为信号;声音刺激前对应的时间窗口动作电位发放数为自发放电,记为噪音。当小鼠处于跑动或者静止时,利用转盘上的速度记录系统记录小鼠的跑动速度,根据转盘的速度来计算小鼠的奔跑速度从而判断小鼠的运动状态。
1.6 数据分析
本研究分别记录了小鼠上行听觉通路中下丘、内侧膝状体和听皮层脑区的神经元响应。记录到的动作电位利用Matlab软件设定的阈值进行识别。分别提取下丘、内侧膝状体和听皮层神经元动作电位的发放个数和发放时间,依据声音刺激参数得到听觉神经元的感受野,随后提取声强级60 dB下神经元不同频率声音刺激的动作电位个数并确定神经元的最佳响应频率(BF)。计算神经元在最佳频率刺激下动作电位在时间序列上的发放数,提取声音刺激开始前的时间窗口动作电位发放数为神经元的自发放数,并计算神经元的自发放率。对每次声音刺激后一段时间内的放电数进行统计,时间段的选择以声音刺激后有明显反应至反应恢复至声音刺激前的时间段为准,用刺激后的放电数减去刺激前放电数来计算诱发反应放电数的增量以及变化率,该增量记为声音刺激期间神经元的诱发反应。分别计算自发放与诱发在不同频率段的概率密度,并根据分布趋势选择合适的函数进行拟合,本研究中的拟合利用Matlab中的内置函数实现。同时定义声音开始后50 ms时间窗口内的最佳频率诱发数除以平均自发为该神经元的信噪比[12],计算IC、MGB、A1所有神经元的信噪比并比较其显著性差异。对小鼠不同运动状态下记录到的神经元进行类似的分析。
1.7 统计学分析
统计下丘、内侧膝状体和初级听皮层的自发与诱发差异性,3个脑区的信噪比差异性比较,以及在不同运动状态下各脑区的自发差异、诱发差异,最后分别比较在不同运动状态下3个脑区的信噪比。3个脑区之间的统计比较均是在整体有差异的情况下再进行两两比较。通过统计分析不同脑区神经元的群体响应探究听觉响应信噪比在上行通路中的演变。统计分析采用SPSS 23.0软件,实验数据以均数±标准误表示,不满足正态分布与方差齐性的数据采用非参数检验(Mann-WhitneyU检验),满足正态分布与方差齐性的数据采用单因素方差分析和配对T检验,P<0.05表示差异具有统计学意义。
2 结果
2.1 上行听觉通路各级核团听觉响应分布
本研究分别在小鼠IC,MGB 和A1 脑区记录到140,186和254个听觉响应神经元。IC,MGB和A1听觉诱发动作电位放电数均值分别为:3.76±0.15,2.41±0.09,2.61±0.14,它们的自发动作电位放电数均值分别为:4.77±0.29,7.41±0.22,6.57±0.47;进一步的函数拟合发现,3个脑区的自发在群体上都较好地符合泊松分布,而诱发更加符合对数正态分布(图1)。
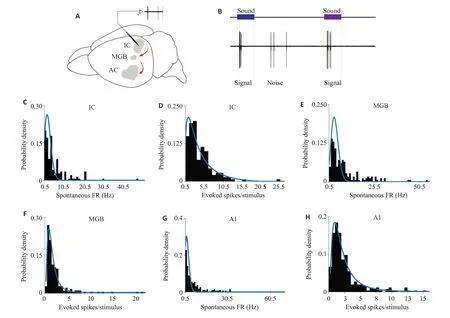
图1 上行听觉通路中各级脑区自发放与诱发的分布趋势Fig.1 Distribution trend of spontaneous firing rate and evoked spike number in response to stimuli in different brain regions in the ascending auditory pathways.A:Recording pattern.B:Signal of sound stimulus.C,E and G:Distribution trend of spontaneous firing rate in the inferior colliculus(IC),medial geniculate body(MGB),and primary auditory cortex(A1),respectively.D,F and H:Distribution trend of evoked spike number in response to stimuli in the IC,MGB andA1,respectively.
2.2 信噪比在上行通路中的演变
图2A~C分别表示IC、MGB、A1脑区神经元自发放与诱发的相关性,从图中可以看出,各级脑区自发放与诱发并没有相关性(IC:R2=0.019;MGB:R2=0.06;A1:R2=0.15)。在上行听觉通路中IC神经元的自发放最低,MGB神经元的自发高于A1(图2D);而IC神经元的诱发是最高的(图2E),本研究中MGB和A1神经元的诱发并没有统计学差异(P=0.123)。从自发放与诱发在各级脑区的演变规律推测上行听觉通路中各级脑区的信噪比变化情况(图2F),从图中可以看到,随着声音向上级脑区的传入,信噪比相对于下丘脑区出现降低的趋势,其中IC、MGB、A1 脑区的信噪比均值分别是2.04±0.14、0.7±0.04、1.08±0.07,从图中很明显可以看到内侧膝状体的信噪比最小。
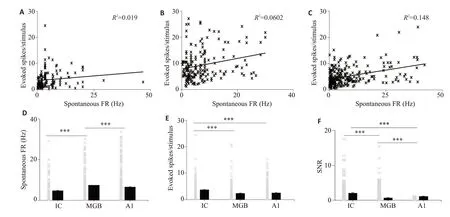
图2 信噪比在上行通路中的演变Fig.2 Evolution of signal-to-noise ratio(SNR)in the ascending auditory pathways.A-C:Correlation between spontaneous firing rate and evoked spike number in the IC,MGB and A1,respectively.D-E:Evolution of spontaneous firing rate and evoked spike number in the ascending auditory pathways.F:SNR in the IC,MGB andA1.***P<0.001.
2.3 运动状态下信噪比的变化
在本研究中,同时记录了小鼠在跑动与安静状态下神经元的响应(图3A),图3A中显示小鼠不同运动状态下的记录模式,固定小鼠头部的同时确保小鼠能够在转盘上自由移动,利用图3 B所示的方法分析不同运动状态下的自发放与诱发。图3C为一个典型的在跑动状态下听觉响应被抑制的例子:小鼠由安静不动的状态改变为跑动状态时,A1神经元的动作电位发放数减少。从群体上看,在A1中,运动状态下神经元的自发放(P<0.001)与诱发(P=0.041)均有下降(图3D,E),而在MGB和IC神经元中,当小鼠由安静状态变为运动状态时,其神经元的自发放与诱发均没有改变(图3F,G和H,I)。同样地统计跑动状态相对于安静状态的信噪比在上行听觉通路中的变化(图3J),从图中可以看出,跑动状态与安静状态相比,各级脑区的信噪比均没有发生显著变化。
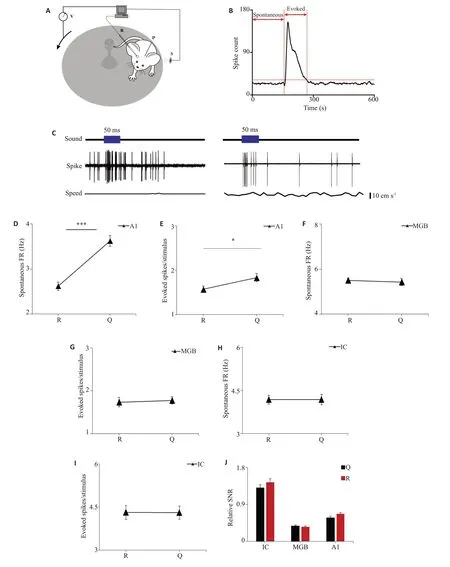
图3 不同状态下上行听觉通路神经元的响应和信噪比总结Fig.3 Summary of response and SNR of the neurons in the ascending auditory pathway in different states of motion.A:Experimental setup(R,recording electrode;P,head-fixation post).B:Calculation model.C:Spike at rest(Q)and during running(R).D-I:Spontaneous firing rate and evoked spike number at rest(Q)and during running(R)in theA1(n=95),MGB(n=98),and IC(n=68).J:Relative SNR.***P<0.001,*P<0.05.
3 讨论
以往研究显示[7,14-16],对数正态分布常被用来描述单个神经元的峰峰值(动作电位两个峰之间的距离)分布,而群体响应经常用指数分布来表示;同时有研究表示[5],听皮层群体神经元的发放率符合对数正态分布,但未对自发分布与诱发分布做区分。本研究对上行听觉通路各级脑区神经元在群体上的自发分布与诱发分布趋势分别进行描述,发现群体神经元的自发和诱发可以分别用泊松分布与对数正态分布来表示。另外从群体神经元对声音敏感性的角度出发,目前大多数研究主要是听皮层在背景噪音下信号的识别[17,18],针对单个神经元乃至整个上行听觉通路神经元的信噪比会发生怎样的变化并没有进行系统的分析。我们通过探究声音信息向上级脑区传输过程中信噪比发生了怎样的变化(本研究中没有考虑自发放为零的神经元),发现声音信息由下丘到内侧膝状体再到初级听皮层过程中,信噪比从IC到MGB呈现下降的趋势,而MGB到A1却又呈现增高趋势。根据以往的研究显示[19],抑制性神经元通过增强或抑制皮层感觉神经元的感受野对听觉加工敏感度有直接的影响,而听皮层神经元对简单音调信息的加工整合处理离不开抑制性中间神经元[20],不同的抑制性中间神经元作用的脑区不同,其中PⅤ中间抑制性神经元在皮层加工处理信息中发挥重要作用,SOM中间抑制性神经元则是在下丘加工处理信息时发挥作用,这可能导致不同脑区诱发与自发之间存在着差异,因此在上行听觉通路中听觉响应的信噪比产生了不同的变化。由于听觉响应的绝对值(听觉诱发动作电位)和相对值(诱发与自发动作电位的比值,信噪比)在上行听觉通路中都处于不断演变的过程,因此很难判断在听觉信息的上行传递过程中哪个指标能有效表征声音特征。
由于在单一状态下无法确定表征声音特征的指标,我们进一步研究不同运动状态下上行听觉通路听觉响应的演变,结果发现运动状态下听皮层的听觉响应绝对值受到抑制,但其相对值却能维持不变。同样的有研究发现,初级听皮层听觉响应在运动状态时减弱[12];Otazu等[13]发现当小鼠参与任务时,内侧膝状体神经元自发放增加,而诱发并没有受到影响;同时有研究表明小鼠运动状态下内侧膝状体神经元自发放与诱发均没有改变[12],因此我们推测在上行通路中IC、MGB、A1脑区也会受到运动状态的影响。但我们的研究结果显示,IC和MGB的听觉响应不受小鼠运动状态改变的影响,而A1神经元在跑动状态下自发与诱发都受到抑制。Zhou等[12]研究表明,听皮层L2/3层神经元在行为活跃时兴奋性和抑制性水平有所降低,而L4层神经元的兴奋性与抑制性水平没有发生改变,这可能导致听皮层在跑动状态下自发放与诱发水平降低。在视觉研究中[12],由光激活PⅤ抑制性神经元引起诱发放电的适度降低可导致视觉检测任务的性能发生显著变化,进一步证明了抑制性神经元与运动状态有关。由于IC和MGB受听皮层抑制性神经元调控较小,因此运动状态下群体神经元的自发放与诱发并没有明显改变。
由于神经元的自发放与诱发受到不同程度的抑制,可能导致听觉响应SNR出现差异。我们的结果显示IC、MGB和A1脑区在不同运动状态下听觉响应SNR没有显著变化,因此推测上行听觉通路中脑区的听觉响应SNR不受大脑状态的影响,听觉处理的敏感性至少在从静止到活跃状态期间保持不变。SNR的稳定性对于动物在不同状态时对外界感觉刺激的感知保持稳定来规避危险和与同类有效交流至关重要[12,21-36],然而SNR 的功能意义仍有待通过行为研究来检验。
综上所述,在听觉神经通路中,神经元以动作电位的方式来编码声音特征[7,13-15],但能更好编码声音特征的是听觉响应的绝对值还是相对值这一直是一个有争议的问题。通过系统研究听觉响应信噪比在上行通路中的演变,分别从自发放与诱发的分布趋势、信噪比在群体上的变化趋势以及在不同状态下信噪比发生的改变等方面探讨,结果显示听觉响应SNR可能是编码声音特征的更好指标。
