低氮胁迫对谷子苗期光合指标及生理性能的影响
2021-12-16王宇珅孟晓伟韩渊怀
王宇珅,张 敏,孟晓伟,韩渊怀
(1.山西农业大学农学院,山西太谷 030801;2.黄土高原特色作物优质高效生产省部共建协同创新中心,山西太谷 030801)
氮是作物生长的必需元素之一,对维持作物新陈代谢、生长发育、高产、稳产具有重要意义。缺氮时作物表现出生长发育受阻,植株个体矮小,叶色发黄的典型表型,影响产量的形成。在农业生产中,施加氮肥往往成为确保作物稳产、高产及优质的主要栽培手段。然而,过量施加氮肥不仅造成氮肥利用效率低下,而且还给生态环境带来了巨大的压力,造成一系列生态失衡问题。据统计,我国氮肥利用效率约为35%,低于世界40%~60%的水平[1]。如何提高氮肥利用效率,特别是提高作物在缺氮环境下的氮利用效率成为亟待解决的关键问题。研究表明,在氮缺乏时,水稻和小麦可通过促进其根系伸长来获得深层土壤的氮素;在玉米的研究中也发现,具有较长根系的品种,具有更高的氮利用效率[2-5]。时丽冉等[6]研究发现,低氮耐受的谷子品种在缺氮时苗高和生物量降低幅度较小、氮利用效率较高、根长增长幅度较大。在苦荞的研究中也发现,缺氮抑制其地上部生长,而促进根系伸长生长,导致根冠比增加[7]。
缺氮也会影响植物体内氮还原同化酶活性。谢孟林等[8]研究发现,氮缺乏时玉米苗期根系硝酸还原酶(NR)、谷氨酰胺合成酶(GS)、谷氨酸脱氢酶(GDH)活性均显著下降。刘鹏等[9]研究表明,耐低氮型高粱品种叶片可保持相对较高的NR 活性。张楚等[7]研究表明,低氮胁迫下苦荞苗期NR 活性、蛋白质含量显著下降。吕雪梅[10]研究发现,NO3--N 缺乏时,小麦根系NR 和GS 活性明显提高;而NH4+-N缺乏时,根系GS 和谷氨酸合成酶(GOGAT)活性显著提高。此外,氮匮乏也会影响植物体内抗氧化相关酶活性。缺氮时,苦荞根系SOD、POD 活性以及根系MDA、可溶性糖及游离脯氨酸含量显著升高。李强等[11]研究发现,氮供应不足时,玉米具有较高的POD 活性以维持其正常的生命活动。王俊等[12]研究发现,对低氮耐受的烟草品种在低氮环境下具有较高的SOD 活性,且积累较少的MDA。
植物中70%的氮存在于叶绿体中,因此,氮素是否充足直接影响着植物叶片叶绿素含量、光合速率以及荧光特性[13]。有研究表明,氮供应不足时谷子叶片中叶绿素含量显著降低[14]。WU 等[15]研究发现,缺氮时,玉米气孔导度(Gs)和光饱和点(Ls)降低,且随着缺氮程度加剧,玉米叶绿素含量显著降低。ZHU 等[16]研究发现,低氮处理下柳枝稷的叶片净光合速率和总叶绿素含量显著降低。ZHAO 等[17]研究表明,缺氮显著降低了高粱叶片叶绿素含量和净光合速率,导致生物量产量降低。于雪等[18]研究表明,缺氮显著降低了甜菜叶片暗适应下PSⅡ光化学最大效率(Fv/Fm)和光化学淬灭(qP),增加了非光化学淬灭(qN),进而抑制PSⅡ光化学活性,降低了光合作用。时丽冉等[19]研究发现,随着供氮水平的降低,小黑麦叶片的初始荧光(Fo)和最大荧光(Fm)逐步升高,PSⅡ原初光能转化效率(Fv/Fm)、PSⅡ潜在活性(Fv/Fo)以及PI abs 均表现为下降趋势。何海锋等[20]研究表明,施中氮提高了柳枝稷的净光合速率(Pn)、气孔导度、细胞间隙CO2浓度(Ci)和蒸腾速率(Tr)等光合指标,进而提高干物质积累。以上结果表明,可通过评价低氮处理下谷子形态、氮同化相关酶、抗氧化相关酶活性及光合性能指标来评价作物的耐低氮能力。
谷子(Setaria italica),是禾本科自花授粉作物,也是我国北方重要的杂粮作物。由于其具有较强的抗逆性,耐旱耐贫瘠,且基因组较小,已经逐渐发展成为新的模式作物。目前针对氮亏缺条件下,谷子苗期形态、氮同化关键酶、抗氧化酶活性及光合性能相关研究的报道还较少,不利于对苗期谷子耐低氮特性的深入研究。
本研究在水培条件下,分析谷子在低氮胁迫下谷子表型性状、生理生化指标及光合性能指标的变化,旨在为谷子耐低氮营养胁迫研究奠定基础,为培育耐逆作物品种提供理论依据。
1 材料和方法
1.1 试验材料
试验材料为EMS 诱变晋谷21 获得的超早熟突变体Xiaomi,经多年种植鉴定,性状可稳定遗传。该材料由山西农业大学杂粮分子育种团队提供。
1.2 试验设计
试验在人工气候室内进行。选取籽粒大小饱满、均匀一致、无病虫害的种子1 000 粒,用10%次氯酸钠和75%酒精依次消毒后,无菌水冲洗5~7 次,置于育苗棉上,蒸馏水浇灌,确保种子萌发。待幼苗长至两叶一心期时,改用Hoagland 全营养液进行培养,每3 d 更换一次营养液,培养期间每天对材料进行通气处理1 h。培养条件为光期16 h,温度28 ℃;暗期8 h,温度22 ℃。待幼苗长至五叶一心期时,开始对材料进行低氮胁迫(LN,硝酸钾为氮源),正常氮处理(NN)为对照。其中,正常氮浓度为2 mmol/L,低氮胁迫浓度为0.2 mmol/L。
1.3 测定指标及方法
低氮胁迫7、15 d 时,分别选择长势一致的正常氮处理及低氮处理的谷子幼苗各10 株,分别对其株高、旗叶长、旗叶宽、叶厚、茎粗、根长、根冠比等进行测定。
低氮胁迫15 d 时,采用凯氏定氮法测定谷子叶片及根系的全氮含量。
低氮胁迫2 h、72 h、15 d 时,测定谷子叶片叶绿素含量。
低氮胁迫7 d 时,采用便携式光合仪(CIRAS-3,美国PP SYSTEMS)测定净光合速率、气孔导度、蒸腾速率及水分利用效率(3 个生物学重复)。
低氮胁迫15 d 时,采用便携式植物效率分析仪(Handy PEA,英国Hansatech),测定叶绿素荧光动力参数(5 个生物学重复)。
硝酸还原酶(NR)活性采用改良版离体法进行测定[21-22];过氧化物酶(POD)活性采用愈创木酚法进行测定[21-22];过氧化氢酶(CAT)活性采用紫外吸收法测定[21-22];超氧化物歧化酶(SOD)活性采用氮蓝四唑法测定[21-22];丙二醛(MDA)含量采用硫代巴比妥酸法测定[21]。
1.4 数据分析
试验采用Excel 2019 和SPSS 22.0 软件进行数据分析。
2 结果与分析
2.1 低氮胁迫对谷子苗期表型性状的影响

由图1 可知,低氮胁迫7 d 时,谷子根系与对照(正常氮)相比显著增长;而胁迫15 d 后,与对照相比,低氮处理后谷子株高显著降低,表现为老叶发黄、嫩叶叶片早衰叶色较浅、叶表面积减少,而根系显著增长、根表面积增加、侧根数量增加(图1)。
进一步对低氮胁迫下谷子苗期的农艺性状进行分析,结果发现(表1),与对照(正常氮)相比,低氮处理下谷子的根长和总根长显著增长,其中,根长显著增加了45.18%(7 d,P<0.05)和50.20%(15 d,P<0.01),总根长在低氮胁迫15 d 时极显著增加了27.73%(P<0.01),表明缺氮时,谷子通过增加根系长度来增加其对氮素的吸收面积。与对照相比,低氮胁迫15 d 时,谷子的根冠比极显著增加12.17%(P<0.01),表明低氮环境可诱导谷子根系发育,使根冠比增加进而响应外界低氮胁迫。
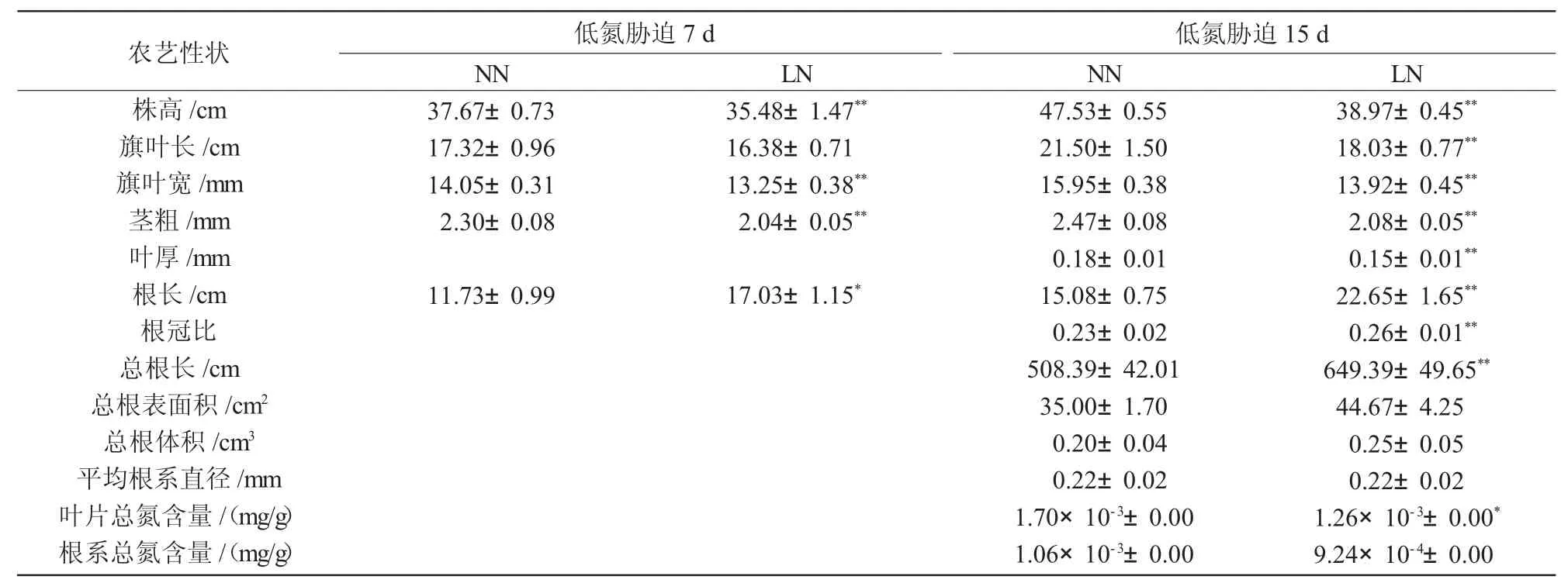
表1 不同氮处理对谷子苗期农艺性状的影响
低氮处理7 d 时,谷子地上部生长显著受到抑制,其中,株高、旗叶宽、茎粗分别较对照显著降低了5.81%、5.69%、11.30%;而在低氮处理15 d 时,谷子的株高、旗叶长、旗叶宽、茎粗、叶厚和叶片总氮含量与对照之间差异更加显著,较对照分别降低了18.01%、16.14%、12.73%、15.79%、16.67%、25.88%。以上结果表明,低氮会抑制谷子地上部分生长,而促进其根系伸长,增加根表面积。
2.2 低氮胁迫对谷子苗期光合性能的影响
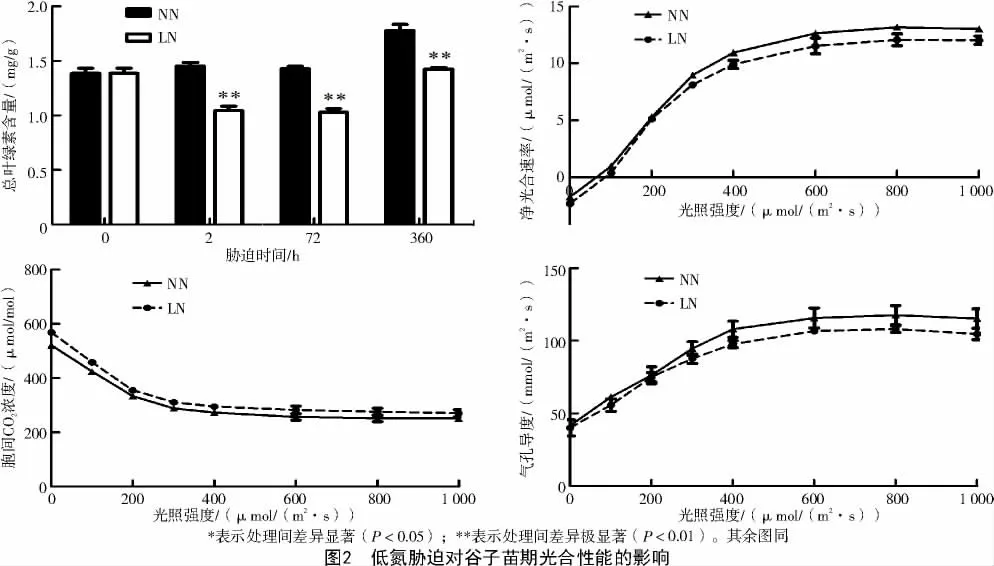
由图2 可知,低氮处理下谷子的总叶绿素含量极显著下降,且在胁迫2 h、72 h、15 d 时,与对照(正常氮)相比分别降低了28.10%、28.07%、19.84%(P<0.01)。表明缺氮造成叶绿素合成受阻,从而影响植物的光合作用。分析低氮胁迫下谷子叶片的净光合速率及光响应曲线,发现低氮处理7 d 时,叶片净光合速率较对照表现为下降的趋势,且在不同光照强度下低氮处理的净光合速率均低于对照,表明缺氮会降低谷子的光合效率(图2)。同时,低氮胁迫下谷子叶片气孔导度下降,而胞间CO2浓度高于正常氮处理,表明低氮胁迫可能影响了光合作用的碳同化效率(图2)。
进一步分析低氮胁迫15 d 后谷子叶片叶绿素荧光参数,结果表明(表2),PI abs、φEo、φRo、ETo/CSm 极显著降低(P<0.01)。叶片Fv/Fm 在低氮环境下变化较小,且随着光强增加并未发现与对照之间存在显著差异,表明叶片光系统不存在光抑制现象,因此,缺氮可能是通过抑制光系统Ⅱ的电子传递效率,进而影响其光合效率。
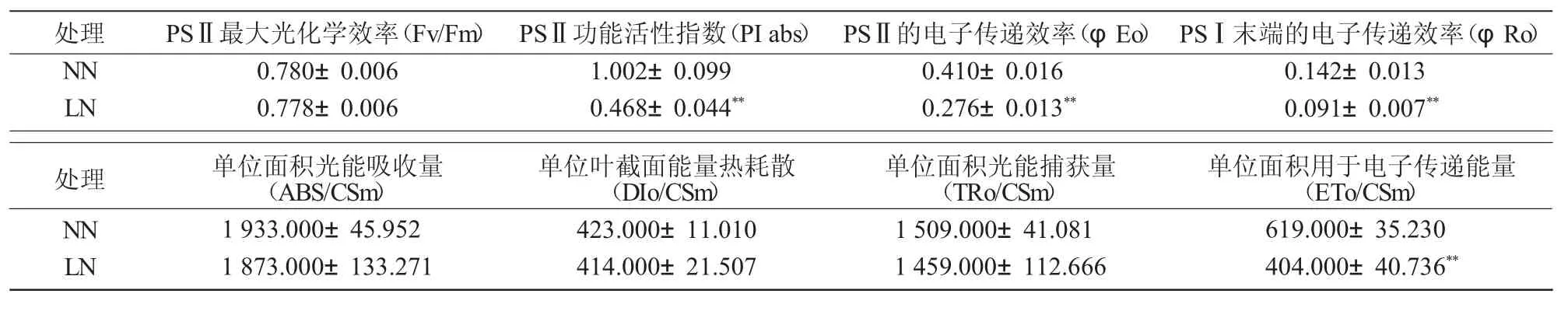
表2 低氮胁迫对谷子苗期叶片荧光参数的影响
2.3 低氮胁迫对谷子苗期膜完整性的影响
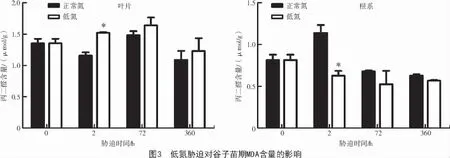
丙二醛(MDA)含量是常用的膜脂过氧化指标,MDA 含量越高,其膜脂过氧化程度就越严重,膜的透性越大。由图3 可知,与对照(正常氮)相比,低氮胁迫导致叶片MDA 含量增加,而根系MDA 含量表现为减少的趋势。其中,谷子叶片MDA 含量在处理2 h 时显著增加了31.42%(P<0.05),表明缺氮时,谷子细胞膜稳定性受到破坏,影响细胞的结构完整性。而根系的MDA 含量降低,且在处理2 h 时较对照(正常氮)显著下降了44.94%(P<0.05),表明低氮胁迫下,谷子根系细胞膜稳定性较好,有利于维持根系对氮素的吸收。
2.4 低氮胁迫对谷子苗期硝酸还原酶活性的影响
由图4 可知,低氮处理72 h 和15 d 时,谷子叶片硝酸还原酶(NR)活性显著降低,且分别降低了48.92%(P<0.01)和46.88%(P<0.05),硝酸还原酶为诱导性酶,受硝态氮的诱导,较低的叶片硝酸还原酶活性表明低氮胁迫下谷子根系吸收的硝态氮分配到叶片的量较少,进而抑制了NR 活性。低氮胁迫2 h 时,谷子根系NR 活性极显著增加了75.83%(P<0.01);而在低氮胁迫72 h 和15 d 时,相较于正常氮处理,低氮处理下的谷子根系NR 活性没有明显变化,表明在短期胁迫(2 h)时,谷子会通过提高NR 活性,促进根系对硝态氮的还原,增加氮吸收水平,从而响应短期低氮胁迫;而在长期胁迫时,由于NR 为硝态氮的诱导性酶,缺氮使得NR 活性诱导水平降低。
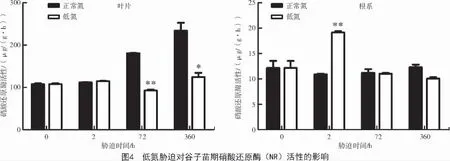
2.5 低氮胁迫对谷子苗期抗氧化酶活性的影响
从图5 可以看出,低氮处理下,谷子叶片过氧化物酶(POD)活性较对照(正常氮)表现为增加的趋势,根系POD 活性则表现为先增加后降低的趋势。与对照相比,低氮胁迫2 h 时谷子叶片POD 活性显著增加了45.01%(P<0.05);而在低氮胁迫72 h和15 d 时,叶片POD 活性与对照相比无显著差异。谷子根系POD 活性在低氮处理2、72 h 时分别较正常氮处理显著增加了20.97%(P<0.05)和41.85%(P<0.05);而在低氮胁迫15 d 时与对照间差异不显著。表明在相对较短期的低氮胁迫下(2、72 h),谷子叶片和根系POD 活性增强,加强其对超氧阴离子等的代谢来适应低氮胁迫。
从图5 可以看出,低氮胁迫2、72 h 时,谷子叶片CAT 活性与对照之间差异不显著,而低氮胁迫15 d 时,谷子叶片CAT 活性较对照显著降低了27.29%(P<0.05)。谷子根系CAT 活性则表现为在低氮胁迫2 h 时较对照显著增加了99.61%(P<0.05);而在胁迫72 h 和15 d 时,根系CAT 活性与对照之间无显著差异。
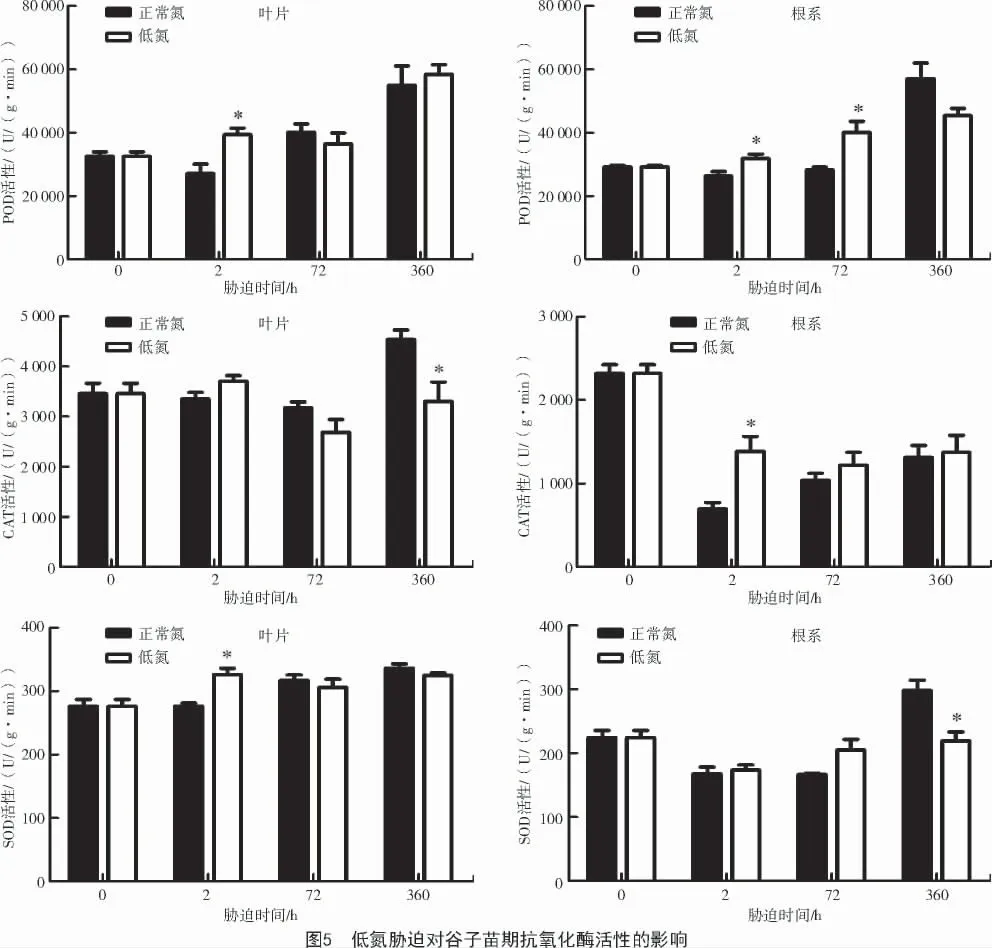
由图5 可知,低氮胁迫2 h 时,叶片SOD 活性较对照(正常氮)显著增加了18.07%(P<0.05);而在低氮胁迫72 h 和15 d 时,叶片SOD 活性与正常氮处理相比有所降低,但差异不显著。与正常氮处理相比,谷子根系SOD 活性在低氮处理15 d 时显著降低,降幅为26.51%(P<0.05);而在低氮胁迫2、72 h 时,根系的SOD 活性有所增加,但与对照间差异不显著。结果表明,低氮胁迫下,谷子叶片和根系SOD 活性均受氮胁迫显著影响,且在叶片中SOD主要通过在短期胁迫中发挥作用,而在根系中SOD主要参与低氮胁迫的长期响应。
3 结论与讨论
氮对作物维持正常生长发育,作物的高产、稳产及优质发挥着重要的作用[23]。张立媛等[24]研究发现,低氮胁迫下谷子株高显著降低,且赤谷6 号、赤谷8 号和赤谷9 号根长显著增长。连盈等[25]研究发现,低氮胁迫下谷子苗期形态上主要表现为叶片数减少、叶长叶宽减小、叶面积变小。程丽丽等[26]研究发现,低氮条件下光皮桦的株高显著降低,且老叶叶色发黄、叶片较小,表现为典型的缺氮表型,同时根冠比、总根长、根系总表面积和根系平均直径均有所增加,但其地上氮含量、地下氮含量和氮累积量均呈下降趋势。陈凌等[27]研究发现,不同糜子品种的株高、叶面积在低氮条件下均显著降低。本研究发现,低氮处理后谷子株高、茎粗显著降低,叶色较浅,叶面积减小,表明低氮胁迫显著影响了谷子叶片的生长发育;根系显著增长,根总长、根表面积及根体积增加,侧根数量增加,且随着胁迫时间的增加,地上部分及地下部分表型差异更加显著,这与前人研究结果一致,表明低氮胁迫会抑制谷子地上部分生长,优先供应地下部分发育以增加土壤中根系表面积和体积,进而响应低氮胁迫。此外,本研究发现,低氮处理15 d 后谷子叶片和根系的总氮含量降低,且叶片中的总氮含量显著下降,该结果与前人结果一致,表明低氮胁迫会影响谷子表型及氮含量。
氮素供应不足,植物叶片的光合性能和叶绿素荧光动力参数都受到很大的影响。植物叶片中的氮素绝大多数存在于叶绿体中,参与了一系列光合进程[28]。有研究表明,在低氮环境下,甜菜第1 对真叶局部逐渐变黄,第2 对真叶生长受阻并且出现早衰现象,功能叶叶绿素含量持续降低[29]。低氮胁迫后,谷子叶绿素含量和光合速率呈下降趋势,表明氮胁迫会降低叶绿素含量、影响叶片发育,进而使植株光合作用降低[30]。苗期低氮胁迫下,玉米叶绿素含量、Fv、Fm、Fv/Fo、Fv/Fm、Fv'/Fm' 和qP 等叶绿素荧光特性也均显著降低,且Pn、Gs 和Tr 呈下降趋势,Ci 则显著上升[31]。本研究发现,低氮处理下谷子叶绿素含量明显下降,这与刘大丽等[29]、李明哲等[30]研究结果一致。低氮处理7 d 时,谷子的净光合速率和气孔导度均呈下降趋势,而胞间CO2浓度呈上升趋势;低氮胁迫15 d 时谷子叶片PI abs、ETo/CSm、φEo 明显降低,表明缺氮抑制了光系统Ⅱ的电子传递效率。以上结果与前人研究一致,表明低氮胁迫下谷子叶片没有发生光抑制,对光能的吸收和捕获影响不显著,但显著降低电子传递的能力以及叶片活性。
植物吸收土壤中的铵态氮或硝态氮,需经过还原同化作用才能合成氨基酸或蛋白质。氮供应不足,植物氨基酸或蛋白质合成受阻,氮代谢途径发生改变,影响植物的生长发育。有研究发现,缺氮时玉米根系NR 活性降低,且NR 活性随氮素营养水平的提高而增强[8]。刘鹏等[9]研究表明,低氮处理后高粱的叶片NR 活性均有不同程度降低,且生育后期高粱叶片NR 活性降低,但是耐低氮型高粱叶片仍保持着相对高的NR 活性。本研究发现,低氮处理下,谷子叶片和根系中NR 活性均显著降低,且叶片中的NR 活性显著高于根系中NR 活性。低氮胁迫2 h 时,谷子根系中NR 活性显著高于对照(正常氮),而在低氮胁迫72 h 和15 d 时根系NR 活性较低氮胁迫2 h 时显著降低,这与前人研究结果不同,因此,推测短期低氮胁迫时,谷子根系NR 活性增强进而促进根系对硝态氮的还原,增加氮吸收水平。
氮胁迫会造成植物体内活性氧及超氧阴离子的大量积累,对植物膜结构造成破坏。POD、SOD 和CAT 是植物抗氧化主要酶,其活性水平能直接反映植物受外界逆境影响的程度,MDA 是膜脂过氧化产物,逆境胁迫下MDA 含量越高,其膜脂过氧化程度就越严重。程丽丽等[26]研究表明,低氮处理下3 个基因型光皮桦幼苗POD、SOD 活性均显著降低。张楚等[7]研究发现,缺氮时不同苦荞品种MDA 含量均显著升高。本研究发现,低氮处理后,谷子叶片及根系(胁迫2 h)的POD 活性表现为增加趋势,说明低氮胁迫下谷子可能通过增强叶片和根系POD 活性来维持细胞内活性氧的代谢平衡,从而降低膜脂受损程度,减缓叶片和根系衰老。低氮胁迫2 h 时,谷子叶片SOD 活性最强,之后开始下降;根系SOD 活性在低氮胁迫15 d 时显著降低,但在低氮胁迫2、72 h 时较正常氮相比呈上升趋势,表明短期低氮胁迫下,谷子可通过提高SOD 活性来维持植物自由基代谢平衡。谷子叶片CAT 活性在低氮胁迫2 h 时呈增加趋势,在胁迫72 h 和15 d 时呈明显下降趋势,而根系CAT 活性呈显著上升趋势。与对照相比,低氮胁迫下谷子叶片MDA 含量呈增加趋势;而根系MDA 含量较对照有下降的趋势,这与前人研究结果不同,推测可能是由于试验条件差异造成的结果。
综上所述,氮缺乏时,谷子通过促进根系发育、减小叶表面积以及降低株高等表型变化来响应逆境胁迫;同时通过调整叶片及根系NR、POD、SOD、CAT 活性及MDA 含量来调节体内氮代谢途径,响应低氮胁迫。
