基于高通量测序技术的苗期抗条锈基因YrCHI 的遗传定位
2021-12-16韩金妤吴启蒙韩德俊吴建辉
韩金妤,刘 丹,张 旭,吴启蒙,韩德俊,吴建辉
(西北农林科技大学农学院,旱区作物逆境生物学国家重点实验室,陕西杨凌 712100)
小麦条锈病是由小麦条锈菌(Puccinia striiformis f.sp.tritici)引起的一种气流传播性真菌病害,是世界范围的重要病害之一,具有分布广泛、大面积暴发流行等特点。我国是世界上规模最大的条锈病流行区系,常年发生面积400.0 万~533.3 万hm2,一般发生区可损失产量10%~20%,严重流行区可达30%以上,自20 世纪50 年代以来,发生4 次大流行,造成巨大产量损失[1-2]。21 世纪以来,由于条锈菌小种的变异导致新致病性小种不断涌现,新小种发生频率一直攀升,造成了西北、西南等小麦条锈病流行区连年成灾,且具有高发态势,严重威胁着我国小麦安全生产[3-5]。因此,对小麦条锈病需加强防治以免小麦条锈病大区流行造成不可挽回的损失。而对于小麦条锈病的防治,目前主要有2 种途径:化学农药和种植抗病品种。其中,化学杀菌剂的应用在病害应急防治中确实发挥了重要作用,但也带来了一系列的“后遗症”,大规模的不规范使用增加了生产成本,还造成了环境污染[6]。大量研究与生产实践表明,抗病品种的选育和应用是防治条锈病最具生态效益和经济安全的措施[3]。但由于小麦条锈菌小种变异频繁,如最新出现的致病小种条中34(CYR34)及其V26 致病类群,导致我国当前生产上大部分小麦品种“丧失”抗性[5]。因此,亟需从小麦材料中发掘和利用新的抗病基因资源以提高作物抗病性。
随着新一代测序和芯片技术的成熟及快速发展,以单核苷酸多态性(Single nucleotide polymorphism,SNP)为标志的第3 代标记被人们大量开发出来,SNP 广泛分布在各个基因组中,而且SNP 类型的标记具有成本低、共显性、易于实现高通量分析等技术优势,因此,很快在水稻、玉米、小麦等作物的基因发掘与利用中大量使用[7]。目前利用测序技术已广泛应用在小麦正向遗传学研究中,如多样性评估、QTL 定位和全基因组关联分析等。同时,针对SNP 的高通量检测技术也相继开发出来,如基于LGC Genomics 公司的竞争性等位基因特异性PCR(KASP)标记[8]、基于北京嘉诚公司的等位基因特异的定量PCR 基因分型系统(AQP)标记[9],这类标记可以更高效地实现基因分型。与凝胶电泳相比,KASP/AQP 标记的使用节省了在SSR 标记筛选、群体基因型分析上的时间,解放了人力,也极大地降低了单个标记分析的成本,提升了技术的可适用性;同时,也为育种家们提供了高通量分子标记辅助选择的技术手段。
西北农林科技大学植物免疫团队长期从事小麦抗条锈病研究,其中小麦抗病遗传与分子设计育种课题组前期从世界各地征集小麦5 000 余份,经过多年多点的鉴定,筛选出数百份条锈病抗性优良的种质[10-18]。其中,来自美国农业部USDA 的品种PI 90279 和国际玉米小麦改良中心(CIMMYT)的品种Chilero 自引进我国后,一直对我国小麦条锈病保持良好的抗性[19-20]。特别是整个全生育期(苗期)针对当前流行最广泛的CYR34 小种都具有抗性,因此,发掘鉴定这类抗性基因对于未来应用于抗病育种具有重要的意义。虽然Chilero 在墨西哥已经保持抗病多年,且PONCE-MOLINA 等[24]对其成株期抗性进行了遗传定位,并分别在染色体1B、5D 和7B 上发现成株期抗性QTL位点Lr46/Yr29、QLr.cim-5DS/QYr.cim-5DS 和QYr.cim-7BL,但是Chilero 与PI 90279 一样,苗期抗性的遗传基础一直尚未得到解析,苗期抗性与成株期抗性之间的关系也尚未得到分析。
本研究在前期研究基础上,拟对2 个小麦品种条锈病全生育期(苗期)抗性进行遗传解析和分子作图,以期完成对目标遗传位点的定位,并获得与目标位点紧密连锁的分子标记,用于分子标记辅助选择育种。研究所发掘的条锈病基因及其连锁标记可用于分子辅助选择育种,可为我国小麦条锈病抗性育种的改良提供重要材料和基因资源。
1 材料和方法
1.1 试验材料
试验以澳大利亚感病品种Avocet S(AvS)为母本、抗病品种PI 90279 和Chilero 为父本,分别杂交构建186 个F2家系和154 个重组自交系(RIL)的遗传群体。铭贤169、Avocet S 和小偃22 为感病对照。
供试小麦条锈菌生理小种为CYR34(实验室代号V26-Lab),由西北农林科技大学植物保护学院提供,经过鉴定、纯化,隔离繁殖后备用。AvS/PI 90279的F2材料均由抗病遗传与分子设计育种课题组保存,AvS/Chilero 的RIL 群体由河南科技大学王春平教授惠赠。
1.2 条锈病苗期抗性鉴定与遗传分析
小麦苗期抗性鉴定在西北农林科技大学植物病理研究所温室进行。将AvS、PI 90279、Chilero、感病对照以及遗传群体种植在10 cm×10 cm×10 cm的塑料方盆中,对于亲本和RIL 家系,每个材料7~8 粒,播种在方盆的4 个角,待小麦幼苗生长至二叶完全伸展时(一般15 d 左右),采用抖粉法(条锈菌小种CYR34∶滑石粉≈1∶50(V/V))进行接种[20]。接种后在9~11 ℃条件下避光保湿24 h,之后将小麦幼苗置于温室中培养,温度控制在18 ℃以下,光照强度为20 000 lx,16 h/8 h 光暗周期,相对湿度80%左右,待感病对照充分发病后(接种后15~18 d)记载AvS、PI 90279、Chilero 和后代遗传群体的反应型。反应型(Infection type,IT)按0~9 级标准记载,其中,反应型0~6 级为抗病型(R),7~9级为感病型(S)。每隔3 d 调查一次,最终结果以最高反应型为准。
根据亲本以及后代群体的反应型,在Excel 中用χ2法对调查获得的分离比进行适合性检测,确定最适合的分离比率,明确供试品种对特定条锈病小种抗性的基因数目、互作方式及抗病特点,进而明确抗条锈病基因的数目及其相互关系。
1.3 集群分离分析
待小麦生长至3 叶期左右,采集各亲本及单株/家系的第3 叶,利用CTAB 法进行DNA 的提取。根据AvS×PI 90279 和AvS×Chilero 的F2和RIL 家系的条锈病抗性分离特点,采用混池分析法(bulked segregant analysis,BSA),分别选择10 株极端抗病单株/家系和10 株极端感病单株/家系按照等量DNA 构建2 组抗感池,经质检无问题后,将构建的抗感池和亲本送往北京博奥生物技术有限公司(http://www.capitalbiotech.com/),利用小麦最新660K SNP 芯片v2.0 进行扫描。样品下机后需进行质量检测合格,然后通过GTC 软件(Affymetrix 公司,http://www.affymetrix.com/estore/)对数据点进行SNP 分型和聚类分析。为了保证结果准确性和可靠性,依据以下过滤标准对SNP 标记进行剔除:无多态性及低质量的、缺失值大于20%的、分群不明显的、等位频率低于5%的等等。基于SNP 所在的染色体物理位置,利用自编Perl 语言脚本确定AvS×PI 90279 和AvS×Chilero 群体抗感池间差异SNP主要集中的染色体区域,初步推断目标基因所在的染色体区域。
1.4 全基因组定位
前期利用DArT 标记对AvS×Chilero 的RIL 群体中140 个家系进行测序分型,并完成了该群体的全基因组范围内的连锁图谱的构建,该数据同样由河南科技大学王春平教授提供。利用IciMapping v4.1(Inclusive Composite Interval Mapping)软件对该群体的苗期抗性表型进行QTL 分析,LOD 阈值设为2.5,用表型变异解释率PVE 描述发掘到的位点的效应。
2 结果与分析
2.1 苗期抗性鉴定与遗传分析
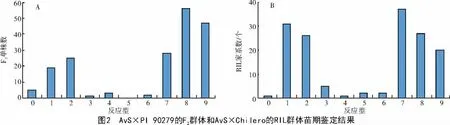
苗期鉴定结果表明,Avocet S 对CYR34 表现为高度感病(IT=9),而PI 90279 和Chilero 表现为高度抗病(IT=1)(图1)。而在AvS×PI 90279 的F2和AvS×Chilero 的RIL 群体后代中,抗病个体/家系绝大多数集中在0~3 反应级上。F2抗病株有53 株(IT 为0~6),感病株有133 株(IT 为7~9)(图2-A),经卡方测验(χ2=0.58,P=0.27>0.05),符合1∶3 的期望比。AvS×Chilero 的154 个RIL 家系中,纯合抗病家系数为68 个,纯合感病株系数为84 个(有2 个家系未获得表型)(图2-B),经卡方测验(χ2=0.84,P=0.19>0.05),符合1∶1 的期望比。说明小麦抗病种质PI 90279 和Chilero 苗期抗性分别由1 对主效基因控制。

2.2 AvS×PI 90279 和AvS×Chilero 群体的抗感池分析
利用小麦660K 芯片对2 个群体的抗感池分别进行扫描,然后将抗感池间表现出纯合差异的SNP进行比较整合。其中在AvS×PI 90279 群体中,从660K 芯片扫描的结果统计抗感池间和亲本共同的多态性SNP 有6 269 个,根据660K 的物理图谱信息,发现分别有2 523、3 746 个SNP 分布在5B 和7B 上,约占总差异的75.71%,另有55 个SNP 未获得定位信息,剩余SNP 落在其他染色体上且在各个染色体上数量不等(图3-A)。以上结果说明,位于5B 和7B 染色体上的SNP 极有可能与抗病基因相关联。同样的,在AvS×Chilero 群体中,从660K 芯片扫描的结果统计抗感池间和亲本共同的多态性SNP 有6 946 个,根据660K 的物理图谱信息,发现分别有2 418、4528 个SNP 分布在5B 和7B 上,约占总差异的39.65%,另有719 个SNP 未获得定位信息,剩余SNP 落在其他染色体上且在各个染色体上数量不等(图3-B)。以上结果同样说明,位于5B和7B 染色体上的SNP 极有可能与抗病基因相关联。考虑到AvS 品种为5BL/7BL 易位系,因此,差异SNP 实际上是位于同一条染色体上。

进一步根据5B 和7B 上的SNP 物理位置信息,统计每兆碱基(Mb)中含有的差异SNP 数量,结果显示,AvS×PI 90279 群体中的7B 染色体上绝大部分SNP 都位于600~720 Mb 区间(图3-C),同样的,AvS×Chilero 群体中的7B 染色体上绝大部分SNP 也都位于600~720 Mb 区间(图3-D),因此,初步推测该区间极有可能就是目标基因或QTL所在的位置;而5B 染色体上的SNP 分布较为分散(未展示)。此外,将2 个群体的抗感池差异SNP 比较发现,2 个群体共同拥有的差异SNP 为2 062 个,其中,5B 上有728 个(图3-E),7B 上有1 334 个(图3-F)。综合以上结果,AvS×PI 90279 和AvS×Chilero 群体中对于条锈菌小种CYR34 的抗性均由1 对基因控制,且位于同一染色体7B 同一区域,推断二者很可能为同一个基因。
2.3 7B 染色体上目标位点的验证
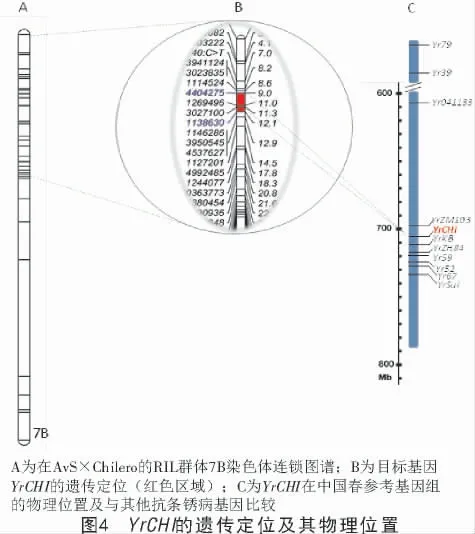
为了验证上述混池分析的结果,进一步利用AvS×Chilero 的RIL 群体的基于DArT 标记的全基因组连锁图谱[24]对苗期抗性表型进行QTL 定位(图4),在7B 染色体的连锁图谱上成功定位出一个主效位点,暂命名为YrCHI,表型变异解释率为82.38%,位于DArT标记4404275 和1138630 之间,根据2 个标记的序列,与中国春基因组比对后发现分别对应于701.28、708.85 Mb,即YrCHI 位于701.28~708.85 Mb区间,该区间包含在混池分析所定位的区间(600~720 Mb),证实了混池分析的结果。
3 讨论
3.1 定位在小麦7B 染色体上的条锈病抗性基因
小麦7B 染色体是一个富含抗病基因的染色体,前人已发掘定位出大量抗条锈病基因[21-23],根据最新的7B 染色体物理图谱,其中Yr6 和Yr63 位于7BS 上,其他基因均位于7BL 上,YrCHI 位于701~710 Mb 之间,其中,Yr79 和Yr39 距离YrCHI 超过300 Mb,Yr041133 距离YrCHI 约100 Mb,YrZM103位于700 Mb 左右,其他基因如YrKB、YrZH84、Yr59、Yr52、Yr67 和YrSui 等都集中在710~740 Mb 的区间。从物理位置看,只有YrZM103 和YrCHI 有重叠区域。从育种系谱上分析看,YrZM103 来源于品种郑麦103,郑麦103 的系谱为“周13/D8904-7-1//郑004”,Chilero 系谱为“4777*2//FKN/Gabo 54/3/Ve ery#5/4/Buckbuck/Pavon F76”[24],其中周麦13 的亲本之一是周8425B,而周8425B 的亲本之一是Nainari60,Nainari60 的系谱为“Supremo/Mentana//Gabo-55/3/Thatcher/Querearo//Kentana/5/Gabo-55”(http://wheatpedigree.net/sort/sh-ow/43385),经仔细分析发现,郑麦103 和Chilero 确实存在比较久远的共同亲本“Gabo”,因此,有可能YrZM103 和YrCHI 为同一个基因,但需要更进一步的等位性分析试验来确认。而前人在Chilero 材料中7B 染色体上定位一个主效的成株期抗条锈病QTL 位点QYr.cim-7BL,该位点介于DArT 标记100006719 和1112830 之间,与YrCHI 的两翼标记4404275 和1138630 相距10 Mb左右,因此认为,控制成株期抗性的QTL 与苗期针对CYR34 小种的抗性位点YrCHI 属于2 个不同的基因位点。
3.2 抗病基因的发掘是培育抗病品种的基础
随着现代生物技术的进步和分子标记技术的应用,大量的抗条锈病基因已被发现并且得到了命名。截至2021 年6 月,正式命名的小麦条锈病抗性基因有83 个,暂命名基因超过300 个[21,25]。虽然发掘的基因数目看似“可观”,但条锈菌不断变异,致使大部分抗条锈病基因已在我国“丧失”抗性,目前仍具有良好苗期抗性的基因有Yr5、Yr15、Yr50、Yr61、Yr69 等[26]。然而这些基因几乎全部来自小麦近缘属种、六倍体斯卑尔脱小麦(Triticum spelta album,Yr5)、野生二粒小麦(Triticum dicoccoides,Yr15),偃麦草属(Thinopyrum,Yr50、Yr69)[27]。这些基因随着外源渗入片段进入普通小麦染色体中,往往还携带一些其他不良的性状,即“连锁累赘”,要想打破他们之间的紧密连锁关系目前还是很困难,比如虽然黑麦1RS 染色体携带抗三锈基因Yr9、Lr26 和Sr31 以及抗白粉基因Pm8,但是1RS 染色体还携带黑麦碱(secalin)基因影响面包加工品质。这类外源基因往往需要通过较长的时间进行选择“改造”以提升其产量品质等,才有可能被育种家利用。因此,发掘的新抗病基因不仅要满足抗性强,抗谱广,还要在产量、品质、农艺性状等方面满足要求,载体品种不能有重大缺陷,这样才能很好地被育种利用。选育推广的栽培品种是优良基因的集合体,一般不存在连锁累赘,往往不需要进行长时间的改良即可利用[28]。如果能从普通小麦品种中发掘和利用抗条锈病基因,将有利于加快抗病育种进程。西北农林科技大学小麦抗病遗传与分子设计育种课题组前期从全世界范围内收集了大量种质,并进行了一系列的性状鉴定评估。本研究从2 个种质中鉴定出相同的针对CYR34 小种呈现抗性的苗期基因,PI 90279 虽为美国农业部收集的种质,但其来源地为中国,不存在适应性的问题,其株高80 cm 左右,穗长9 cm 左右,千粒质量44.9 g,穗粒数53 个,籽粒为红色,沉降值为29(数据未发表);与对照小偃22 相比,开花期晚5~6 d,成熟期相当。而Chilero为CIMMYT 推广的品种,自引进我国后,其生态适应性良好,尚未发现不良性状,其为春性小麦,且含有矮秆基因Rht2、抗叶锈病基因Lr13、高分子量谷蛋白亚基基因Glu-A1a、Glu-B1c、Glu-D1a(http://wheatpedigree.net/sort/show/12452);与对照小偃22 相比,开花期早6~7 d,成熟期早4~5 d。因此,2 个种质在未来的抗病育种中一定具有良好的应用前景。
4 结论
本研究结果表明,利用混池分析结合高通量测序技术快速实现苗期抗条锈病基因YrCHI 的快速定位,发掘大量的SNP 可开发高通量的分子标记用以辅助选择,所发掘的YrCHI 及其分子标记为小麦抗病育种提供良好的基因资源及其种质资源,也为条锈病抗性改良奠定了基础。
志谢:本研究特别感谢河南科技大学王春平教授赠予的小麦Chilero 遗传群体。
