林木种子休眠与萌发机制研究进展*
2021-12-15王艳梅张小雪朱秀征张志华耿晓东蔡齐飞
王艳梅 张小雪 朱秀征 张志华 李 志 耿晓东 蔡齐飞 刘 震
(河南农业大学林学院 中原地区森林资源培育国家林业和草原局重点实验室 郑州 450002)
休眠是植物为长期有效抵抗外界恶劣环境而产生的适应性,影响植物的分布时空(Wangetal., 2019a)。种子休眠与萌发是林木生命活动中的关键发育步骤,在生物学和生态学上具有重要意义。种子休眠在一定程度上可降低同物种间的个体竞争(Finkelsteinetal., 2008),但这类种子只有解除休眠的状态才能够萌发,给种群的延续造成了困扰,给森林培育及遗传育种带来了挑战。近年来,相关研究者对种子的休眠特性与萌发机制进行了大量研究并取得一些成果,为种子贮存、科学解除休眠提供了指导。
1 林木种子休眠
1.1 种子休眠定义
有关“种子休眠”目前仍存在争议,Crocker(1916)提出“种子在生长被抑制之前或伴随生长过程产生休眠”,并将自然条件下来年春季不能出苗的成熟种子称为“二年种子”。Harper(1957)认为植物种子存在生长与休眠2种状态,此观点意味着种子在不发芽时是休眠的,但种子不发芽与休眠是显著区别的2种现象(Vleeshouwersetal., 1995)。种子休眠程度决定种子能够发芽的条件范围(Fenneretal., 2005)。休眠的定义适用于种子休眠的具体情况(Graemeetal., 1981),但大多都是指“种子为植物提供的一种机制来延迟发芽,直到下一代生存的最佳条件”(Finkelsteinetal., 2008)。一些林木种子的休眠是由胚胎周围组织的机械屏障阻碍胚根正常发育而诱发的(Bewley, 1997a; 1997b),但休眠并非是由不利的环境条件强加于种子的停滞发育状态(Lietal., 1997)。目前,种子休眠被广泛认为是“有利环境下,具有生活力的种子仍不能萌发”的现象。
1.2 休眠的原因
1.2.1 环境因素 林木种子通过感知温度来判断外界环境变化。温度信号参与调节种子酶促反应、呼吸作用等(秦爱丽等, 2020),诱导种子休眠以及休眠的深度。不同林木种子休眠特性对温度的响应不同,例如,板栗(Castaneamollissima)在贮藏期间,需要较低温度(0~2 ℃)延长休眠期(王贵禧等, 1999); 杉木(Cunninghamialanceolata)种子暴露在35 ℃条件下会产生二次休眠(Caoetal., 2016)。
光通过作用于种子内的光敏因子在休眠中发挥重要作用。远红光条件相当于黑暗环境,有利于解除种子休眠,红光相当于白光,可以延缓休眠解除,而白光中的蓝光,能够促进胚中ABA合成基因的表达,正调节种子休眠(李振华等, 2019)。
氧气是种子内部代谢活动的动力。供氧不足时,种子内部营养物质转化会受到抑制,并且种胚会因缺氧而产生休眠。红松(Pinuskoraiensis)(Qietal., 1993)、马占相思(Acaciamangium)(符近等, 1997)、三药槟榔(Arecatriandra)(杨期和等, 2005)等种子因种皮结构透气性较差而产生休眠。
充足的水分能推动种子内部贮藏物质的转化及细胞器的活化,是种子发育的动力。因此,环境湿度也会影响其休眠。
1.2.2 种子内源因素 种皮结构影响着种子透水性、通气性以及透光性。酸豆(Tamarindusindica)(马天晓等, 2012)、刺槐(Robiniapseudoacacia)(Naziretal., 2014)、桧柏(Sabinachinensis)(马拉青呼, 2019)、砂生槐(Sophoramoorcroftiana)(安毅鹏等, 2020)和顶果木(Acrocarpusfraxinfolius)(柯碧英等, 2020)等林木种子种皮坚硬或外包裹蜡质层,阻碍水分、气体的交换从而诱导休眠。青钱柳(Cyclocaryapaliurus)(杨万霞, 2004)、大山樱(Prunussargentii)(王艳华等, 2005)等种子种皮对胚根伸长产生抑制作用,诱导休眠。
林木种子中可能含酚类、有机酸、生物碱及醛类等萌发抑制物,这些抑制物存在于种子不同部位,能够阻碍种子吸水、改变渗透压、抑制呼吸及酶活性等。黄檗(Phellodendronamurense)(Chienetal., 2006)、三桠苦(Melicopepteleifolia)(鲁强, 2019)等种子由于种皮存在抑制物或者被覆萌发抑制物而产生休眠。欧榛(Corylusavellana)(Pipinisetal., 2020)、中泰南五味子(Kadsuracoccinea)(陈伟等, 2020)等果皮中含有抑制物; 青钱柳(杨万霞, 2004)、紫椴(Tiliaamurensis)(高博等, 2015)、珊瑚朴(Celtisjulianae)(刘序, 2019)、三桠苦(Evodialepta)(鲁强, 2019)、紫楠(PhoebeSheareri)(姜宗庆等, 2020)等种皮存在抑制物; 羊角槭(Aceryangjuechi)种子的内、外种皮与种胚均存在萌发抑制物质(陈香波等, 2017),而三角槭(Acerbuergerianum)(肖志成等, 2008)、南方红豆杉(Taxuswallichianavar.mairei)(于海莲, 2009)等种子的种胚含有抑制物。
种子种胚发育不成熟也可导致休眠。圆齿野鸦椿(Euscaphiskonishii)(游双红, 2013)、中泰南五味子(陈伟等, 2020)、长白楤木(Araliaelata)(范文丽, 2015)等种子成熟后,胚存在形态休眠(种胚尚未形成、胚分化尚不完善或种胚完成分化但体积很小),仍需从胚乳或其他组织中吸收养料继续生长,直到完成形态后熟; 三药槟榔(杨期和等, 2005)、铜钱树(Paliurushemsleyanus)(曹运梅等, 2019)、三桠苦(鲁强, 2019)、顶果木(柯碧英等, 2020)等种子的种胚在形态上似已发育完全,但种胚的生理状态不具备发芽条件,即生理后熟。而刺楸(Kalopanaxseptemlobus)(黄玉国等, 1990)、银杏(Ginkgobiloba)(崔慧, 2017)等种子同时存在形态与生理后熟现象。
胚乳作为种子的营养组织对其休眠具有重要影响。在紫荆(Cercischinensis)种子发育过程中,胚乳细胞壁具有较高的机械强度,胚乳对胚生长的机械阻碍成为其休眠的重要因素(周健, 2016); 南京椴(Tiliamiqueliana)种子胚乳细胞内物质代谢(杨艳华, 2009)及北五味子(Schisandrachinensis)(郎文香等, 2010)胚乳中内源抑制物是引起种子休眠的主要因素。
种子休眠的产生往往是多种因素共同作用的结果。青钱柳(杨万霞, 2004)、黄檗(Chienetal., 2006)、紫椴(高博等, 2015)、珊瑚朴(刘序, 2019)等种子是由于种皮结构密实且含有萌发抑制物质而诱导种子进入深度休眠状态; 红松(Qietal., 1993)、三药槟榔(杨期和等, 2005)等种子休眠是由于机械障碍及种胚发育不成熟诱导; 铜钱树(曹运梅等, 2019)、顶果木(柯碧英等, 2020)等种子同时存在种皮障碍和生理后熟现象; 中泰南五味子果肉、种皮都存在抑制物且种胚发育不完整,是其休眠的原因(陈伟等, 2020); 圆齿野鸦椿(游双红, 2013)、三桠苦(鲁强, 2019)种子休眠原因比较复杂,不仅存在种皮结构障碍、含有萌发抑制物,并且种胚发育也不完全。
1.3 种子休眠类型划分
植物种子的休眠种类繁多,针对这种复杂的生理现象,Crocker(1916)根据解除休眠的方法将种子休眠划分为种皮不透水或不透气、种皮的抑制作用、成熟的胚、胚内代谢失调、次生休眠、混合类型(其中2种或2种以上的组合)等7种类型。Villiers(1975)认为种子休眠可分为2种类型: 一是种子胚发育被外覆物抑制而休眠,二是胚本身特性诱发的休眠。Roberts等(1977)根据休眠出现期将种子休眠划分为初生休眠、强迫休眠及次生休眠3类。Nikolaeva(1977)根据诱发种子休眠原因将休眠划分为外源性、内源性休眠以及组合休眠,并且认为物理休眠(透水障碍)、机械休眠(胚外被覆物阻碍其生长)及化学休眠(含萌发抑制物)属于外源性休眠,生理和形态休眠应划分到内源性休眠的范畴。Fenner等(2005)在Roberts理论基础上,将“强迫休眠”划入了“次生休眠”的范畴,认为种子休眠有初生和次生休眠2种。陈伟等(2010)依据诱发休眠的部位将种子休眠分为胚休眠、种壳休眠和综合休眠。潘琳等(2016)将种子休眠类型分为生理休眠(包括光信号、胚及抑制物等导致)和强迫休眠2类。Baskin等(2004)将种子休眠分为生理休眠、形态休眠、形态生理休眠、物理休眠以及综合休眠5种类型。与Nikolaeva分类系统相比,Baskin等的分类方法将化学休眠、机械休眠划入生理休眠范畴,是目前较全面的种子休眠类型划分系统。
1.4 种子休眠机制
1.4.1 内源激素调控学说 内源激素的变化影响着植物响应环境的灵敏性,是调节植物(芽、种子等)生长发育的关键机制(王艳梅等, 2012),在植物生命活动中具有重要作用(房丽莎等, 2020)。Wareing等(1971)认为种子休眠由脱落酸(abscisic acid,ABA)、赤霉素(gibberellin,GA)、细胞分裂素(cytokinin,CK)等激素决定。种子内部ABA水平影响休眠深度(Yanetal., 2020),并积极调控休眠的诱导及维持,ABA代谢及信号通路中相关基因调控种子休眠(表1)。种子处于休眠状态时,有高水平的ABA以及较低水平的GA含量,GA通过其生物合成通路和信号转导通路负向调控种子休眠(表2); 而生长素(auxin,IAA)在其信号转导过程中,通过增强种子对ABA的敏感性,正向调控休眠(表3); 乙烯(ethylene,ETH)对种子休眠解除有一定的促进作用(康云清, 2016)(表4)。

表1 脱落酸代谢及信号通路中与种子休眠调控相关基因

续表1 Continued

表2 赤霉素代谢及信号通路中与种子休眠调控相关基因

表3 生长素代谢及信号通路中与种子休眠调控相关基因

表4 乙烯代谢及信号途径中关键基因与种子休眠调控
另外,油菜素甾醇(brassinosteroids,BR)和细胞分裂素、水杨酸(salicylic acid,SA)、茉莉酸(jasmonic acid,JA)等激素也能够通过ABA或GA的信号通路间接调控种子的休眠(Néeetal., 2017)。因此,ABA、GA的生物合成与代谢控制着种子休眠产生以及休眠的深度,是种子休眠与解除的关键。
1.4.2 其他调控学说 光敏素(phytochrome,PHY)主要通过使基因活化或表达、调节某些酶的活性、改变细胞膜的透性、调控脱落酸和赤霉素代谢与信号基因的表达来影响休眠。PHY在感知光照及温度信号时有不同分工: 作为光受体,PHY调控种子萌发受光波长及辐射强度的影响; 作为温度受体,PHY受环境及种子吸胀温度影响(李振华等, 2019)。
基因调控学说认为,基因调控种子的休眠过程是复杂的。ABA对基因组转录具有阻遏作用,影响DNA活性,并通过抑制特定类型mRNA的产生维持休眠(Wareingetal., 1971)。种子在休眠过程中仍然进行核酸的代谢(藤伊正, 1980),高浓度的ABA能够抑制核酸的正常代谢,干扰核糖核酸RNA的合成,从而阻抑了蛋白质和有关酶的合成,使种子不能进行正常的代谢活动(郑光华, 2004)。在休眠解除的初期,油松(Pinustabulaeformis)(周学权, 1989)、玉兰(Yulaniadenudata)(杨孝汉等, 1991)种子RNA的合成受到翻译水平的抑制,之后新RNA合成,DNA转录开始参与休眠的调节。
细胞膜相变论认为,温度能够通过影响细胞膜的状态而调控种子休眠性。温度升高,可能使细胞膜从凝胶态转变为流体态,从而影响相关酶活性,并导致细胞膜透性变化和物质渗透,从而调控与种子休眠相关的代谢过程。
除此之外,还有学者提出呼吸途径调节学说、代谢物调控途径、Ca2+调节理论等来描述种子休眠的机理。
2 种子休眠解除方法与机制
2.1 激素解除休眠
种子由抑制物质起主要作用转向促进物质占主导地位,是解除休眠的决定性因素。休眠解除过程受多种激素控制。长白楤木种子在贮藏过程中赤霉素3A(gibberellin A3,GA3)、ABA、玉米素核糖苷(zeatin riboside,ZR)、吲哚-3-乙酸(indole-3-acetic acid,IAA)4种激素的消长变化对种胚发育及休眠解除具有重要作用(范文丽, 2015)。Wang等(2018)研究山桐子(Idesiapolycarpa)种子休眠解除过程ABA、IAA、GA3和ZR水平的变化,发现IAA、GA3、反式玉米素核糖苷(trans-zeatin riboside,tZR)能够有效促进种子休眠解除。ABA含量与GA3/ABA比值可能是解除休眠期的关键因素(Wangetal., 2019b),并且山桐子种子的发芽率与GA3/ABA 和(IAA+GA3+tZR)/ABA的比值均显著相关(王艳梅等, 2018)。
2.2 层积解除休眠
种子经过层积处理一方面能够增加种皮的透性,促进种子生理后熟; 另一方面可以减少萌发抑制物,从而调控种子内部激素水平向利于休眠解除与萌发方向发展。不同休眠特性种子,层积时所需温度不同(刘震等, 2004; 王艳梅等, 2015)。新成熟的杉木(Caoetal., 2016)、桧柏(马拉青呼, 2019)、山茶(Camelliajaponica)(张宁等, 2015)等种子需要经过低温层积解除休眠,获得更高的发芽率及更快的发芽速度; 而山茱萸(Cornusofficinalis)种子需要变温层积环境(洑香香等, 2013)。
2.3 化学物质解除休眠
利用酸、碱、过氧化物、含氮化合物等化学物质处理能够破坏种皮来增加种皮透性打破种皮障碍、调节代谢或者降解萌发抑制物等解除种子休眠(鲁强, 2019)。酸豆(马天晓等, 2012)、刺槐(Naziretal., 2014)、砂生槐(安毅鹏等, 2020)等一些由种皮包裹蜡质层或有其他被覆物导致休眠的种子,可以利用酸、碱液有效破除种皮障碍。
2.4 物理方法解除休眠
2.4.1 烟和热解除休眠 燃烟中的一氧化氮(nitric oxide,NO)具有刺激植物萌发的作用(Keeleyetal., 1997); 烟中可溶解的氰醇类(cyanohydrins),如氨基乙腈(glycoloni-trile) 、丙酮氰醇(acetone cyanohydrin)等通过缓慢释放氰化物(cyanide)可以打破种子休眠状态,刺激植物萌发。从燃烟中提取的化合物卡里金(karrikins,KAR1)达到一定浓度后可提高种子的萌发率(Flemattietal., 2004)。并且,植物对KAR1的响应依赖其休眠程度,需要经过后熟的植物对KAR1敏感性会更强。有些植物种子可能先需要合适的温度和湿度打破休眠,增加对KAR1的敏感性,进而再促进萌发(周峰, 2013)。热处理打破休眠的本质在于增加种皮的透气性,解除休眠则需要根据种子类型施加不同温度处理。
2.4.2 激光解除休眠机制 激光对生物体有一定的光压作用,可通过电磁效应、光效应、热效应和压力效应等在一定范围内对生物体、组织、细胞等产生极大的影响,导致种子内部生物能的改变,调控种子发育,从而促进休眠解除(李蓉等, 2005; 李宇赤等, 2019)。
2.4.3 物理损伤解除休眠 砂生槐(安毅鹏等, 2020)等“硬实”种子的休眠可利用去皮或划破种子等物理方式,增加种皮透性打破机械束缚,以解除休眠。
3 林木种子萌发
3.1 萌发的定义
种子萌发通常指种子开始吸收水分(种子吸胀),重新开启代谢过程,胚根突破周围结构完成萌发(Bewleyetal., 2013)。种子吸胀时种皮软化,透气性提高,并通过糖酵解途径、三羧酸循环和磷酸戊糖等途径进行呼吸作用,获取能量,推动种子内部物理状态发生变化,继而在适宜环境下完成萌发。
3.2 激素调控种子萌发
激素通过其生物合成通路和信号转导通路拮抗地调控种子萌发。ABA与GA通过抑制对方合成与分解代谢的基因而产生拮抗作用,由激素含量变化引起的种子发育过程中的生物发生,影响种子萌发状态。与ABA相比,GA在种子休眠解除到萌发的过程中发挥主要作用(Finkelsteinetal., 2008),其通过消弱种皮障碍、弱化胚乳的束缚推动萌发;Auxin能够负向调控种子萌发(帅海威等, 2016); ETH可以促进种子种胚细胞的增长、胚轴伸长(康云清, 2016),也能够减轻ABA对萌发的抑制、与GA相互促进,在种子萌发中也很重要(方志荣等, 2007); CK在种子萌发时,能够减轻种子受逆境(干旱、盐碱和重金属等)胁迫程度,调控胚胎发育(宋松泉等, 2021)。除此外,BR、SA、JA等也能够作用于ABA与GA的信号通路促进种子萌发(表5)。

表5 激素代谢及信号途径中关键基因与种子萌发调控

续表5 Continued
3.3 miRNA调控种子萌发
在萌发阶段,种子内部蛋白活性恢复,细胞中某些基因开始表达转录成信使RNA(Messenger RNA,mRNA),这些mRNA与种子内部原本的贮藏mRNA开始翻译与萌发有关的蛋白质。microRNA(miRNA)是一类小分子非编码RNA,参与种子发育、休眠与萌发等植物生命过程(魏俊等, 2017),它通过和靶基因mRNA碱基配对,以降解mRNA或阻碍其翻译进而抑制目标基因的表达,也可以直接切割目标mRNA调控种子萌发(表6)。miRNA在天女木兰(Magnoliasieboldii)种子萌发时的表达集中在细胞生理、代谢等生物过程(魏俊, 2017); 对银杏种胚差异miRNA靶基因分析,发现其主要富集在激素信号、代谢通路、生物合成等途径(何青松, 2018)。因此,miRNAs的差异表达特性在萌发发育中有重要调控作用。

表6 miRNA参与种子萌发调控功能
种子萌发与内部能量物质的代谢、激素信号启动以及传导等过程密切相关,除此之外,还受到转录翻译、细胞伸长及活化、修复机制和细胞器重组等遗传性因素的影响。
3.4 营养物质调控种子萌发
种子萌发是一个需能过程,这个过程涉及到营养物(碳水化合物、蛋白质、脂肪等)的分解以及能量的释放(尹华军等, 2004)。可溶性糖(葡萄糖、蔗糖)能够通过糖酵解、磷酸戊糖途径为种子萌发提供能量,并且能够维持细胞渗透调节。可溶性糖开始下降,说明种子已经处于活跃状态,开始消耗可溶性糖来进行萌发(张存虎, 2018)。在种子内部多种功能蛋白质中,贮藏蛋白与种子萌发关系是最为密切(岳佳铭等, 2021)。银杏种子胚乳中的贮藏蛋白质是氮源的供应者,在种子萌发过程中,贮藏蛋白质被动员,降解为胚生长所需的营养物质(李生平, 2004)。淀粉作为重要的供能物质,经淀粉酶水解后,为种胚生长发育供能。高温下,淀粉酶活性会受到抑制,导致紫椴种子中淀粉分解缓慢,可溶性糖和蛋白代谢也受抑制,导致种子难以正常萌发(段加玉等, 2013)。可溶性蛋白有利于增强种子的抗逆性,且在细胞渗透调节中具有重要作用,经相关酶降解后,有利于种子吸胀,为细胞分裂生长、种子发育提供所需能量。天女木兰(刘月洋, 2010)、桢楠(Phoebezhennan)(龙汉利等, 2013)、珊瑚朴(刘序, 2019)等种子萌发过程中,可溶性蛋白、淀粉、可溶性糖等贮藏物质水平皆总体下降,为萌发供能。
3.5 活性氧分子调控种子萌发
活性氧(reactive oxygen species,ROS)是在逆境下调节植物生长发育的一种重要的多功能信号分子(Qietal., 2018),在植物遭受逆境胁迫时,ROS会过度积累而导致植物的多种细胞成分氧化损伤,最终导致种子的萌发能力降低,而参与ROS调控通路又是种子降低氧化危害、应对逆境胁迫的重要策略(Chenetal., 2014)。ROS对种子萌发的作用是间接的。对于马槟榔(Capparismasaikai)(赵东兴等, 2018)、美国无刺皂荚(Gleditsiatriacanthosvar.inermis)(刘婷等, 2018)、大叶女贞(Ligustrumlucidum)(贺桂兰, 2020)等一些种皮透气性差的种子,使用过氧化氢(hydrogen peroxide,H2O2)可以软化种皮,降低种皮对胚生长的束缚作用,促进萌发。SA和H2O2共同处理玉米能显著提高内源SA和H2O2的含量,且与GA生物合成相关的基因ZmGA20ox1和ZmGA3ox2会表达上调,与GA分解代谢相关的ZmGA2ox1基因表达会明显下调,进而降低种子萌发抑制基因ZmRGL2的表达,促进种子萌发(Lietal., 2017)。ABA抑制种子的萌发可能与ABA的降解增加H2O2的积累量有关(Anandetal., 2019),而ROS又能通过转录效应来刺激GA的生物合成(Lietal., 2018)。以上研究表明,ROS通过调节激素代谢来调控种子萌发。因此,保持种子内部ROS含量的平衡对种子萌发非常重要。
4 种子休眠与萌发
休眠与萌发是种子2个不同的发育阶段,既存在联系又相互区别。休眠能够决定种子发芽所需的条件,当给予适宜的外部环境后,解决种子自身产生的抑制作用便可以引起萌发。这2个步骤的核心是ABA/GA平衡: 成熟种子处于休眠状态时,ABA和IAA含量高,GA含量低; 种子萌发时,其他激素也能够影响 ABA与GA的生物合成与代谢信号通路,激素间相互作用可以精确控制种子休眠与萌发(图1)。
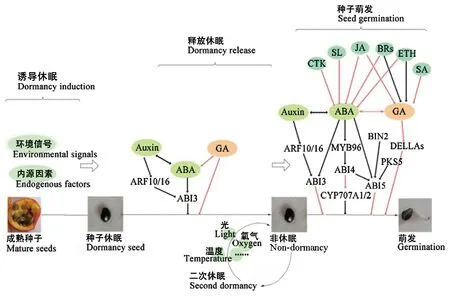
图1 种子休眠解除与萌发过程
5 结论与展望
种子休眠与萌发受遗传因素和环境因素的控制,是植物生长发育进程中的2个关键环节。目前,对少数植物种子休眠与萌发的机制已有较为清晰的认识,尤其在激素信号调控休眠与萌发方面,已取得重大进展。种子休眠与萌发的机制在不同植物中具有一定的保守性,但单、双子叶植物间及不同植物间存在一定的特异性。与模式植物及一些农作物、园艺作物相比较,林木生长周期长,相关的遗传基础和分子机制尚不清楚,有关其林木种子休眠解除与萌发机制研究不够完善,林木种子休眠与萌发的分子机制仍知之甚少,林木种子休眠与萌发的探究仍是林学研究的难点。因此,今后可在以下几方面进行深入研究: 1)自然萌发率低的木本植物种子休眠、萌发机制; 2)影响种子休眠和萌发的因素之间是如何相互作用的,尤其激素和环境对不同休眠类型的种子休眠和萌发的调控; 3)多角度多学科交叉探讨种子休眠与萌发过程中发生的生理生化代谢反应和分子信号转导网络; 4)更多蛋白互作的体内证据,调控林木种子休眠与萌发的差异基因的筛选、主效基因挖掘及其功能分析。
