LPS经鼻AD小鼠脑海马mRNA表达谱分析
2021-12-06程良慧曾娜王燕唐亮
程良慧 曾娜 王燕 唐亮

摘要:目的: 研究LPS经鼻阿尔茨海默(Alzheimer's disease, AD)样小鼠模型组与对照组之间mRNA的表达差异。方法: 利用基因芯片技术检测AD模型组与对照组mRNA表达谱。对差异mRNA进行KEGG通路分析和GO分析。运用qRT-PCR对基因芯片结果进行验证。结果: 在AD模型组和对照组之间共检测到123个mRNA(其中43个上调,80个下调)表达有显著差异(倍数变化值>2,P<0.05)。GO与KEGG分析表明差异表达的mRNA主要参与细胞分化,钙离子结合,炎症和神经元功能或神经系统疾病等。qRT-PCR验证基因芯片结果是可信的。结论: AD模型组与正常对照组间的mRNA表达谱存在较大差异,差异表达的mRNA在AD发病中起着重要作用。
关键词: mRNA;阿尔茨海默病;微阵列
阿尔茨海默病(Alzheimer's disease,AD)是一种随年龄增长无性别差异的进行性神经变性疾病。随着世界范围内人口人数增多和人口老龄化的日益加重,全球患阿尔茨海默病的人数呈指数增加。根据国际阿尔茨海默协会(International alzheimer's association,ADI)2014年公布的数据,全球约有44.40×106人患病,每年新增人数约7.70×106人,平均每3秒就有1名AD患者。据2015年世界阿尔茨海默病报告,中国痴呆人数高达950万,就这一趋势到2050年会有高达135×106人患病。不仅AD的患病率高,治疗的费用及护理费用也非常昂贵,给平常家庭和社会造成沉重的经济负担。有数据显示,2010年,全球每年用于AD的治疗及护理费用约604×109美元。
AD主要临床表现为记忆渐进性减退、认知功能障碍、行动能力逐渐丧失等。AD的神经性特性是老年斑(Senile plaque,SP)的出现、β淀粉样蛋白沉积(β-amyloid,Aβ)、神经原纤维缠结(Neurofibrillary tangles ,NFTs)及神经元丢失伴胶质细胞增生等[1-3]。AD的发病机制尚不清楚。AD的发生与遗传、自由基损伤、凋亡、炎症等因素有关。AD的致病因素包括教育程度低、头部外伤、性激素水平降低、家族遗传史、吸烟、抑郁症状、心理压力、血管因素、体温较低、社会活动较少、轻度认知损伤以及病毒感染等[4]。AD的早期阶段是沿着海马穿过通路脑区逐渐出现灰质体积萎缩出现记忆认知障碍,随着病情的加重,颞叶、额叶与顶叶新皮质也会受到影响,导致语言能力,视觉,行为等功能障碍。
信使核糖核酸(Messenger RNA,mRNA)是生命活动的重要物质之一,是一类由DNA(脱氧核糖核酸)转录合成的携带遗传信息的单链核糖核酸(RNA)。mRNA在核糖体上作为蛋白质合成的模板,能决定肽链的氨基酸排列顺序。有实验证明,mRNA参与了细胞的调节与分化[5]。在生物体内由特定的mRNA来编码多肽链。mRNA虽在细胞内的种类繁多,但其拷贝数只有1-10个。研究表明,mRNA能调节正常基因转录的稳定性及在神经的发生、突触的可塑性中发挥着重要作用。Wu等[6]发现Bhlhb2 mRNA为miR-494的靶基因,Bhlhb2(Bhlhe40)是脑源性神经营养因子的重要转录抑制因子,它的沉默促进了神经突轴突与树突的发生。还有许多研究发现异常表达的 mRNA 与多种重要疾病的发生发展也密切相关,例如肝癌、肺癌、食管腺癌、前列腺癌等癌症[7-8]以及心脑血管疾病、糖尿病等慢性疾病[9]。最近许多研究发现一些易感基因会影响AD发生如BACE1、PS1/2、APP、APOE、SORL1[10-11]等。尽管mRNA与多种疾病的发生发展都有着密切联系,但是上述基因并不能完全解释AD的整个发病机制。因此,本研究在前期建立LPS经鼻(由鼻侧滴注LPS)AD样雄性小鼠模型,通过基因芯片技术探究AD小鼠海马mRNA的表达谱,为进一步研究AD的发病机制提供实验证据。
一、材料与方法
1.动物分组及处理
取5月龄雄性KM小鼠24只,体重28±6g,25±1.5℃恒温饲养,饮食飲水自由。24只小鼠分为2组:模型组(n=12)和对照组(n=12)。模型组经鼻滴注10%LPS溶液0.3ul,对照组经鼻滴注0.9%生理盐水0.3ul,持续5个月,构建AD小鼠模型。
2.RNA样本收集与芯片检测
样品总RNA利用NanoDrop ND-2000 (Thermo Scientific)定量并经Agilent Bioanalyzer 2100 (Agilent Technologies)检测RNA完整性。RNA质检合格后,样本的标记、芯片的杂交以及洗脱参照芯片标准流程。首先,总RNA反转录成双链cDNA,再进一步合成用Cyanine-3-CTP(Cy3)标记的cRNA。标记好的cRNA和芯片杂交,洗脱后利用Agilent Scanner G2505C(Agilent Technologies)扫描得到原始图像。
3.数据分析
采用Feature Extraction 软件 (version10.7.1.1, Agilent Technologies)处理原始图像提取原始数据。接着利用Genespring软件(version 13.1, Agilent Technologies)进行quantile标准化和后续处理。标准化后的数据进行过滤,用于比较的每组样本中至少有一组100%标记为“P”的探针留下进行后续分析。利用T检验的p值和倍数变化值进行差异基因和差异mRNA筛选,筛选的标准为上调或者下调倍数变化值>= 2.0且p值<= 0.05。对差异基因和差异lncRNA进行非监督层次聚类,利用热图的形式展示差异基因在不同样本间的表达模式。
4.荧光定量RT-PCR验证芯片结果
从上述两组海马组织内提取总RNA后行qRT-PCR,随机挑选6对差异表达mRNA验证基因芯片筛选结果。
5.GO与KEGG分析
GO(基因本体论,Gene Ontology)包括三大板块,生物学过程(Biological Process,BP),细胞组分(Cellular Component,CC)和分子功能(Molecular Function,MF),所以有三类结果。统计每个GO 条目中所包括的差异基因个数,并用统计检验的方法计算每个GO 条目中差异基因富集的显著性。
利用KEGG数据库对差异基因进行Pathway分析,并且用统计检验的方法计算每个Pathway条目中差异基因富集的显著性。计算富集显著性的p值,小的p值表示差异基因在该Pathway 中出现了富集。
二、结果
1. mRNA差异筛选
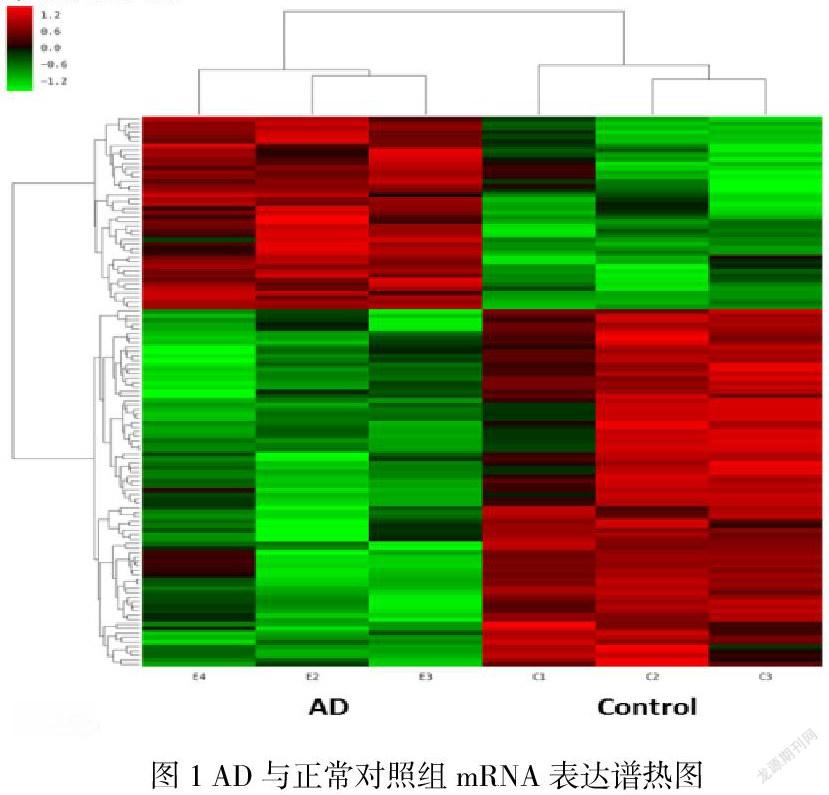
AD模型组与对照组通过差异倍数和显著水平所测到的mRNA差异表达的有123个,其中上调的有43个,下调的有80个(图1);其中有上调和下调最多的已知mRNA,部分mRNA与神经元/神经系统疾病、炎症、嗅觉通路相关的mRNA。
2.qRT-PCR结果
随机挑选4个上调或下调的mRNA并设计引物,然后对芯片结果进行qRT-PCR验证。Itpr2与Icam1的 AD模型组低于空白对照组表示下调,Kcnq5与Cb1b的AD模型组高于空白对照组表示上调,其结果与芯片结果一致(图2)。芯片结果是可信的。
3. KEGG分析
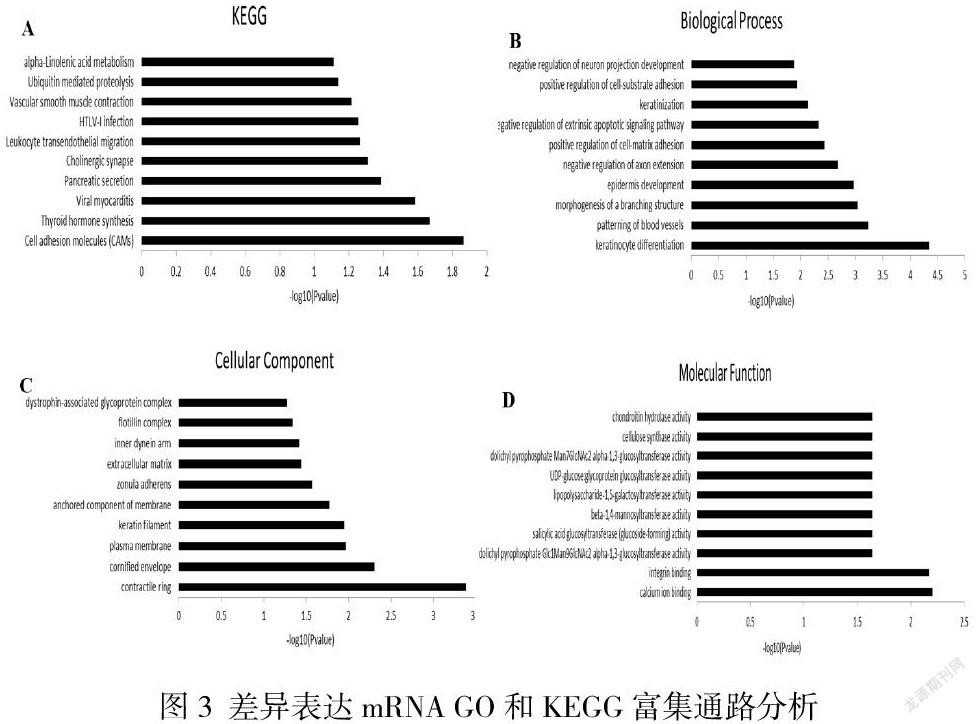
如图3A,差异表达的mRNA相关KEGG富集通路有90条。一些差异表达的mRNA参与炎症(包括NF-kappa B信号通路、肿瘤坏死因子信号通路、炎性介质管理的渠道,和细胞粘附分子)和神经元功能或神经系统疾病(包括阿尔茨海默病、亨廷顿氏舞蹈症中的刺激神经组织的互动,和突触囊泡循环)等。
4.GO分析
在生物学过程(Biological Process)中细胞分化的P值最小,在细胞组分(Cellular Component)中细胞成分的收缩环富集数最多,在分子功能(Molecular Function)中钙离子结合最显著,由此可见GO富集分析显示,差异表达的mRNA主要集于其生物过程中的角质形成细胞分化、细胞成分的收缩环和分子功能中的钙离子结合。(图3B-D)。
三、讨论
在本研究中,我们研究了mRNA在经鼻LPS介导的小鼠阿尔茨海默病模型中的表达谱。AD小鼠与对照组共鉴定出123个mRNA (43个上调,80个下调)表达差异。
LPS可以通过腹腔注射或大脑立体定向注射来建造AD模型[4]。前者可能导致全身炎症反应,后者可能在模型中模拟急性或亚急性反应。两种模型均不能复制AD的慢性、局部炎症和进行性退行性性变。此外,LPS是空气污染物PM2.5的组成成分,可通过鼻吸收,绕过血脑屏障。研究表明,AD和PD患者长期暴露于LPS中后,会出现嗅觉丧失和嗅球病理改变[5]。在本研究中,我们成功构建了单侧经鼻灌注LPS的AD模型,为脑内慢性炎症介导的AD发病机制提供了一种新的研究手段。据我们所知,这是首次在LPS介导的AD模型中研究海马mRNA,这可能有助于我们了解有关mRNA与脑内慢性炎症介导的AD发病机制。
本研究中所检测出的差异表达基因与炎症及神经元功能或神经疾病有关,如阿尔茨海默病,亨廷顿氏症,神经活跃的配体受体,以及突触小泡周期。综上所述,我们已经识别出一些失调的mRNA可能是AD的潜在的生物标志物或靶点。
四、结论
本研究共检测到123个mRNA(其中43个上调,80个下调)。此类mRNA在AD发病机制中有重要作用,后期将对关键基因进行功能验证。
参考文献
[1] 黄鑫, 高旭东, 梁剑平等. 阿尔兹海默病致病机制及环境对其发病影响[J]. 西北民族大学学报(自燃科学版), 2016, 37(2): 51-56, 82.
[2] 李穎新, 李建更, 阮晓钢. 肿瘤基因表达谱分类特征基因选取问题及分析方法研究[J]. 计算机学报, 2006(02):324-330.
[3] 杨楠楠, 魏阳, 徐倩,等. 阿尔兹海默病表观遗传学研究的进展[J]. 中华医学遗传学杂志, 2016, 33(2):252-255.
[4] 赵波, 张新宇, 付学锋. 阿尔兹海默病动物模型的研究进展[J]. 神经解剖学杂志, 2012, 28(001):102-104.
[5] 朱亚萍, 薛雯, 张军平. 阿尔茨海默病与嗅觉障碍[J]. 中国临床医生 2008年36卷10期, 32-33页, ISTIC, 2008.
[6] Wu W, Bhagat T D, Yang X, et al. Hypomethylation of noncoding DNA regions and overexpression of the long noncoding RNA, AFAP1-AS1, in Barrett's esophagus and esophageal adenocarcinoma[J]. Gastroenterology, 2013, 144(5): 956-966.e954.
[7] Wang Y, He L, Du Y, et al. The long noncoding RNAlncTCF7 promotes self-renewal of human liver cancer stem cells through activation of Wnt signaling[J]. Cell Stem Cell, 2015, 16(4): 413-425.
[8] Qiu M, Feng D, Zhang H, et al. Comprehensive analysis of lncRNA expression profiles and identification of functional lncRNAs in lung adenocarcinoma[J]. Oncotarget, 2016, 7(13): 16012-16022.
[9] Moran I, Akerman I, Van De Bunt M, et al. Human beta cell transcriptome analysis uncovers lncRNAs that are tissue-specific, dynamically regulated, and abnormally expressed in type 2 diabetes[J]. Cell Metab, 2012, 16(4): 435-448.
[10] Perez SE, Lumayag S, Kovacs B, et al. β-Amyloid Deposition and Functional Impairment in the Retina of the APPswe/PS1ΔE9 Transgenic Mouse Model of Alzheimer’s Disease[J]. Invest Ophthalmol Vis Sci, 2009, 50(2): 793-800.
Filali M, Lalonde R, Rivest S. Cognitive and non-cognitive behaviors in an APPswe/PS1 bigenic model of Alzheimer's disease[J]. Genes Brain & Behavior, 2009, 8(2): 143-148.
作者簡介:
唐亮,副教授,研究方向:神经退行性疾病发病机制及治疗研究。
本项目受湖南省教育厅科研项目(19C0214),湖南省自然科学基金青年项目(2019JJ50697),广西省教育厅科研项目(2021KY1959),长沙市杰出创新青年人才计划(kq2009095)资助。
