外源核苷酸对干酪乳杆菌生长的促进作用及机制
2021-12-03丁婷,李勇
丁 婷,李 勇
(北京大学公共卫生学院,北京 100191)
益生菌是一类对人体有益的活性微生物,主要定居于肠道、生殖道内。益生菌具有多种生理功能,不仅可以调节肠道功能,维持肠道菌群的平衡,还可以促进钙、磷等微量元素的吸收,而且在治疗腹胀、腹泻、哮喘、痛风、糖尿病等疾病时发挥着重要作用[1-3]。然而,益生菌产品也具有一定的应用局限性,如对外界环境及胃环境敏感,或营养条件要求高而易失活,起不到对人体的有益作用。现有能提高益生菌活性的物质有低聚果糖、低聚半乳糖、葡聚糖等,但这些快速发酵性寡糖能引起人体腹泻和胀气等缺点[4]。因此,寻找对人体安全且能促进益生菌生长增殖的物质亟待解决。
核苷酸是生物体内极重要的低分子化合物,是DNA和RNA的基本组成单位,作为细胞的主要成分,在细胞代谢、能量和功能调节等方面起到重要作用。研究发现,核苷酸具有多种生理功能,如免疫调节[5]、肠道菌群调节[6-7]、肠道修复[8]、抗感染[9]、促进生长发育[10-11]、延长寿命[12]等。由于机体从头合成核苷酸的能力有限,在一些特殊情况下内源合成的核苷酸不能满足机体需要,因此补充外源核苷酸则尤为重要。核苷酸也由“非必需营养素”转变为“半必需营养素”和“条件营养素”。由于其安全性,目前已添加到婴幼儿配方奶粉中使用。
干酪乳杆菌(Lactobacillus casei)是在食品中应用广泛的一种益生菌,具有降血压、降胆固醇、缓解乳糖不耐症、调节肠道菌群等益生作用,逐渐成为研究、开发的焦点,因此寻找促进干酪乳杆菌生长繁殖的物质具有重要意义。研究表明,外源核苷酸是肠道菌群的重要益生元[13-15]。鉴于此,本研究以干酪乳杆菌为研究对象,探究外源核苷酸对干酪乳杆菌的益生效应及机制,以期为提高干酪乳杆菌的存活率提供支撑。
1 材料与方法
1.1 材料与试剂
干酪乳杆菌Q1为从辽宁锦州酸菜中分离纯化鉴定的菌株,保藏于本实验室-80 ℃冰箱。菌株按1%比例接种于MRS肉汤培养基中,37 ℃、160 r/min摇床培养24 h进行活化。
外源核苷酸5’-单磷酸胞苷(cytidine 5’-monophosphate,5’-CMP)、5’-单磷酸腺苷(adenosine 5’-monophosphate,5’-AMP) 大连珍奥生物技术股份有限公司;N-己酰基-L-高丝氨酸内酯(N-hexanoyl-L-homoserine lactone,C6-HSL) 西格玛奥德里奇贸易有限公司。
1.2 仪器与设备
LabRAM HR Evolution拉曼光谱仪 法国HORIBA Scientific公司;LRH型生化培养箱 上海一恒科技有限公司;LDZX-50FBS立式压力蒸汽灭菌器 上海申安医疗器械厂;TCS-SP8激光共聚焦显微镜(confocal laser scanning microscope,CLSM) 德国徕卡公司。
1.3 方法
1.3.1 外源核苷酸对干酪乳杆菌生长的影响
以MRS培养基为空白对照,分别加入1%、2%、3%、4%的5’-CMP和5’-AMP,接种1%的干酪乳杆菌,37 ℃摇床培养24 h,每隔4 h取一次样,采用二倍稀释法对干酪乳杆菌进行活菌计数,绘制生长曲线。
1.3.2 干酪乳杆菌生长变化规律
微生物生长动力学模型可以描述不同生长环境和条件下微生物的数量与时间的关系。修正的Gompertz[16]和修正的Logistic模型[17]通常用于拟合微生物的生长数据。分别用修正的Gompetz模型、修正的Logistic模型非线描述干酪乳杆菌的生长变化规律。
修正的Gompertz模型:
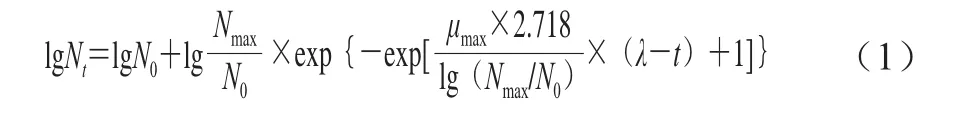
式中:t为时间/h;Nt为时间t时的菌数/(CFU/mL);Nmax为N0为最大和初始菌数/(CFU/mL);μmax为最大比生长速率/h-1;λ为生长延滞时间/h。
修正的Logistic模型:
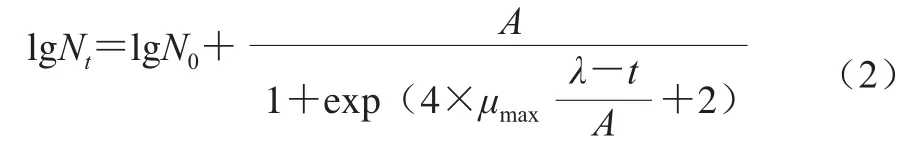
式中:Nt为时间t时的菌落数/(CFU/mL);N0为初始菌落数/(CFU/mL);A为拟合参数;μmax为最大比生长速率/h-1;λ为生长延滞时间/h。
1.3.3 外源核苷酸对干酪乳杆菌生物被膜、胞外聚合物(extracellular polymeric substances,EPS)的影响
干酪乳杆菌(1%)接种于MRS肉汤培养基中37 ℃、160 r/min摇床培养24 h。按1∶100比例稀释后,将100 µL菌液和4.0%的5’-CMP和5’-AMP分别加入孔板中,MRS肉汤作为对照。孔中放入无菌盖玻片和打磨过的锌片(0.5 mm×0.5 mm,0.3 mm),37 ℃静置培养48 h后,用无菌水缓慢冲洗盖玻片3 次,0.4%结晶紫溶液染色20 min,在显微镜下观察细菌生物被膜的变化。将锌片用无菌水缓慢冲洗除去未黏附的细菌。用2.5%的戊二醛溶液固定30 min后干燥30 min。用0.01 g/100 mL的吖啶橙溶液避光染色15 min后,磷酸盐缓冲液(phosphate buffered saline,PBS)冲洗3 次,干燥30 min。加入10 μL的封片剂,用CLSM进行观察(发射波长525 nm,激发波长488 nm)。锌片用无菌水缓慢冲洗后干燥,用拉曼光谱检测外源核苷酸对干酪乳杆菌EPS含量和化学成分的影响。
1.3.4 外源核苷酸添加对干酪乳杆菌粗提物群体感应抑制活性及抗生物被膜活性的影响
干酪乳杆菌(1%)接种于MRS肉汤中,37 ℃、160 r/min摇床培养24 h。按1∶100比例稀释后取1 mL菌液加入100 mL的MRS肉汤中,加入4.0%的5’-CMP和5’-AMP,MRS肉汤作为对照,37 ℃、160 r/min摇床培养48 h后,将菌液于12 000 r/min离心4 min,取上清液。用同体积的乙酸乙酯(含0.1%冰醋酸)萃取2 次后静置1 h。将萃取后的乙酸乙酯进行旋转蒸发(35 ℃)去除溶剂,加入1 mL甲醇溶解粗提物并用0.22 μm的滤膜过滤,贮藏于-20 ℃冰箱备用。
测定干酪乳杆菌粗提物的群体感应抑制活性前,需测定粗提物的最小抑菌浓度,以排除抑菌性的干扰。粗提物的最小抑菌浓度参照文献[18]所述方法进行测定。在亚抑菌浓度下,粗提物的群体感应抑制活性通过报告菌株紫色色杆菌(Chromobacterium violaceum)CV026进行验证。紫色色杆菌CV026培养至OD600nm为1.0。按1∶100的比例将培养物加入冷却至50 ℃左右的LB琼脂培养基,培养基中加入10 μL C6-HSL,混合均匀后倒平板(平板直径90 mm)并用牛津杯打孔。将100 μL粗提物加入平板的孔中,置于培养箱中28 ℃培养24 h并观察平板颜色变化。每组3 个平行。为探究干酪乳杆菌粗提物抗生物被膜活性,将志贺菌(Shigella)按1%比例接种于LB肉汤培养基中28 ℃、160 r/min摇床培养24 h,按1∶100比例稀释后取100 µL菌液加入96 孔板中,分别加入不同培养条件的干酪乳杆菌粗提物,不加粗提物的肉汤作空白对照。将96 孔板放置于28 ℃培养箱中静置24 h。培养后,用无菌PBS冲洗3 遍。将VK3(丙酮溶解,10 mmol/L)加入2,3-二-(2-甲氧基-4-硝基-5-磺苯基)-2H-四氮唑-5-甲酰苯胺(2,3-bis (2-methoxy-4-nitro-5-sulfo-phenyl)-2H-tetrazolium-5-carboxanilide,XTT)(0.5 mg/mL,PBS溶解)中达到1 μmol/L浓度。将200 μL XTT-VK3溶液加入96 孔板中,37 ℃避光放置2 h,在490 nm波长处用酶标仪进行测试。每个样品重复4 次取平均值。
1.3.5 干酪乳杆菌粗提物群体感应抑制活性的影响因素
为测定酶对干酪乳杆菌粗提物群体感应抑制活性的影响,向干酪乳杆菌粗提物中分别加入最适pH值下的胃蛋白酶、木瓜蛋白酶、胰蛋白酶和碱性蛋白酶,使其最终质量浓度为2.0 mg/mL,将上述溶液于37 ℃水浴锅中放置4 h后用紫色色杆菌CV026平板测定其群体感应抑制活性,其中未加酶的粗提物作为对照。为测定温度对干酪乳杆菌粗提物的群体感应抑制活性的影响,将干酪乳杆菌粗提物置于80 ℃和100 ℃水浴锅中30 min,未处理的粗提物作为对照。
1.4 统计分析
2 结果与分析
2.1 外源核苷酸对干酪乳杆菌生长增殖的影响
图1显示,干酪乳杆菌在添加外源核苷酸的MRS培养基中较空白对照生长快,菌体迅速繁殖,很快进入对数生长期,活菌数较不添加核苷酸的空白组明显提高,外源核苷酸浓度越高促进效果越显著。其中,5’-AMP对细菌生长的促进作用更明显,表明外源核苷酸可以加速细菌的增殖。用微生物生长动力学模型对细菌的生长曲线进行非线性拟合,结果显示,修正的Gompertz方程和修正的Logistic方程可以较好地描述微生物的生长动态(R2>0.919 78)。修正的Gompertz方程和修正的Logistic方程拟合后所得相关参数如表1所示。添加5’-CMP和5’-AMP后,细菌的最大比生长速率(μmax)较空白对照明显增加,而延滞期(λ)则明显缩短。上述结果表明外源核苷酸5’-CMP和5’-AMP对细菌的生长有明显的促进作用。
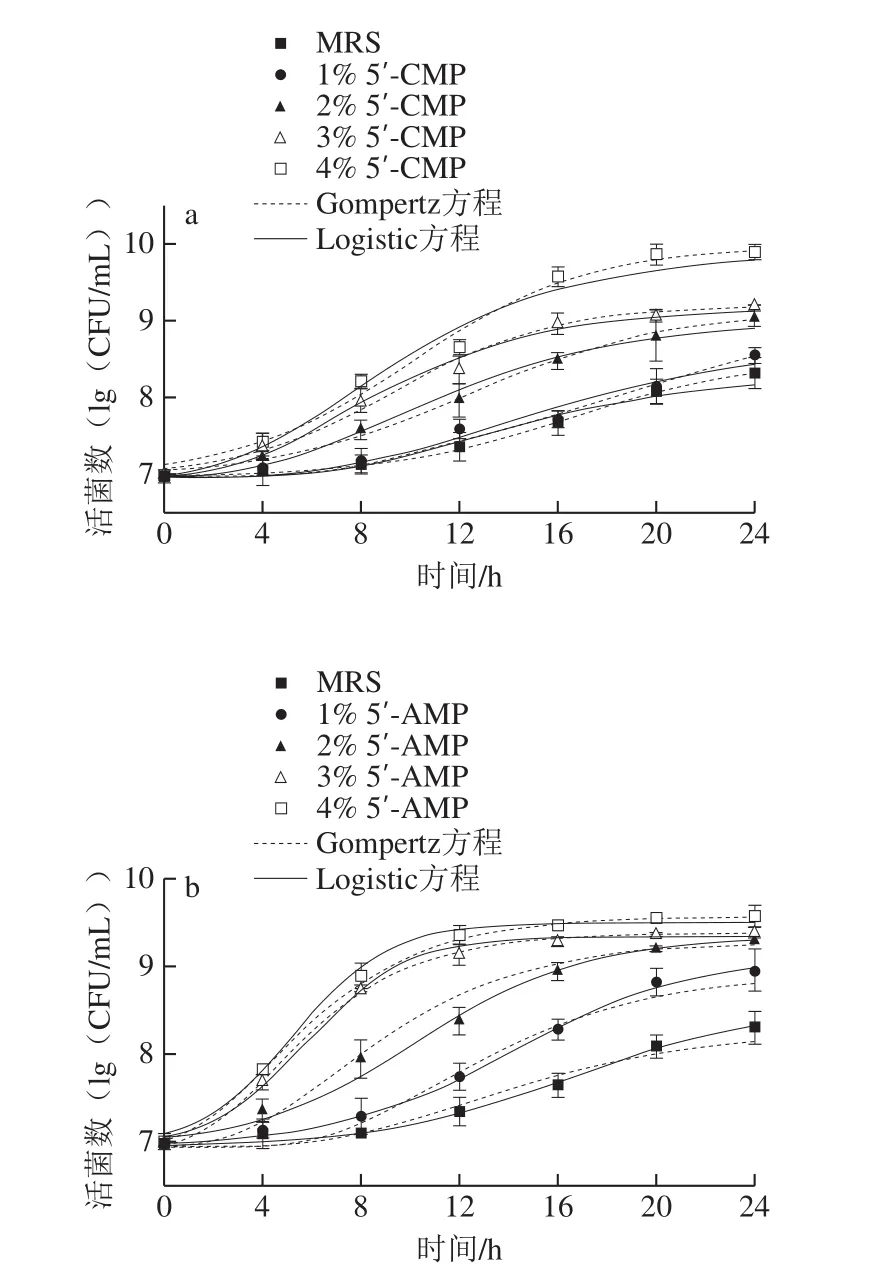
图1 干酪乳杆菌生长曲线用修正的Gompertz方程以及修正的Logistic方程拟合结果Fig.1 Growth curves of L.casei fitted with modified Gompertz and modified Logistic equation
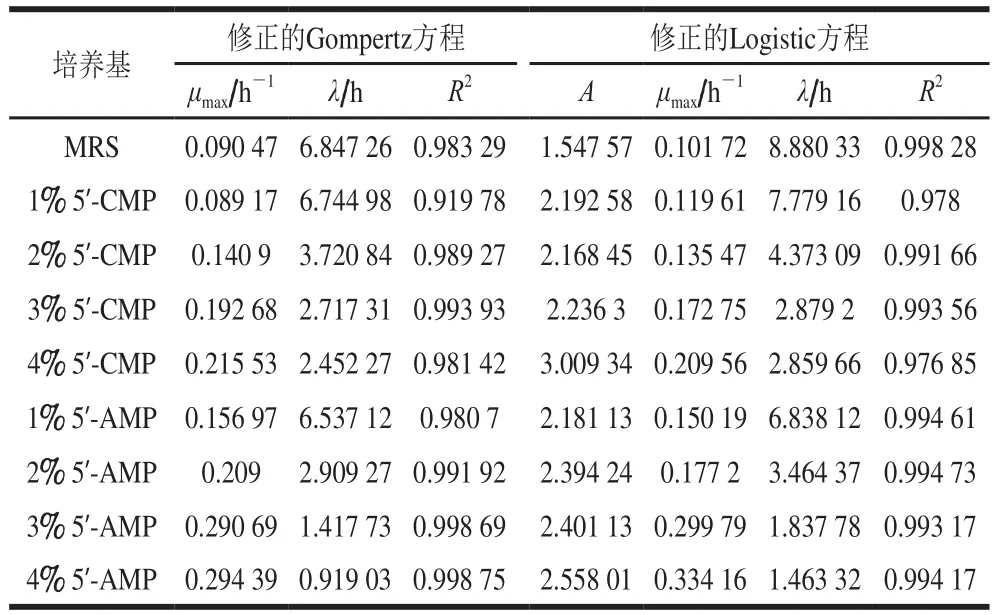
表1 干酪乳杆菌用修正的Gompertz和修正的Logistic方程非线性拟合相关参数Table 1 Nonlinear parameters of L.casei fitted with modified Gompertz and modified Logistic equation
2.2 外源核苷酸对干酪乳杆菌生物被膜的影响
图2a~c显示,干酪乳杆菌添加外源核苷酸后,细菌生物被膜分泌增多,细菌由较为分散的状态变得聚集,5’-AMP对细菌生物被膜的刺激作用更加明显。图2d~f显示,对照生物被膜厚度为30 μm,添加5’-CMP后生物被膜增加到55 μm,添加5’-AMP后生物被膜则增加到70 μm,结果表明外源核苷酸5’-CMP和5’-AMP促进了干酪乳杆菌生物被膜的分泌。
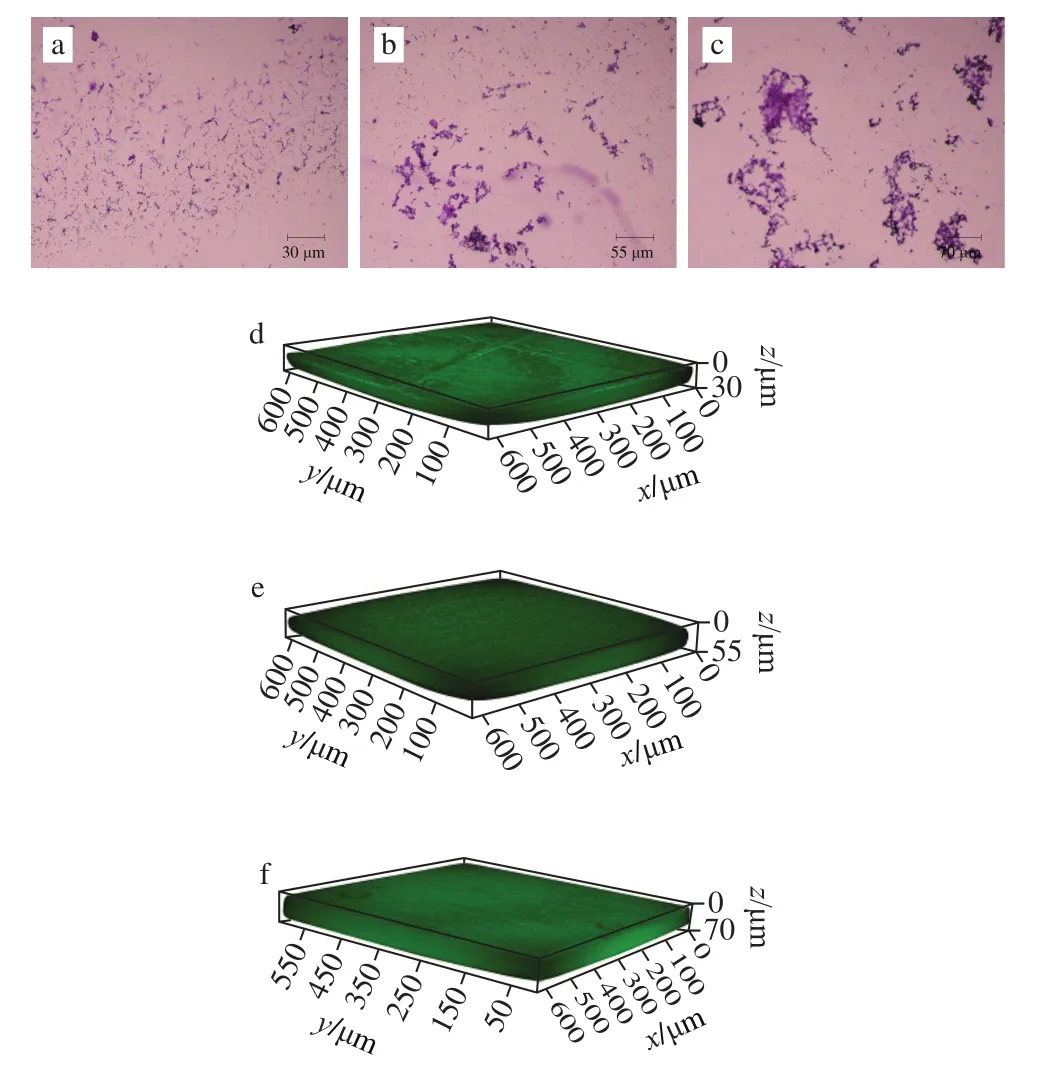
图2 光学显微镜及CLSM观测外源核苷酸对干酪乳杆菌生物被膜的影响Fig.2 Effect of exogenous nucleotides on the biofilm of L.casei Q1observed by optical microscopy and CLSM
2.3 外源核苷酸对干酪乳杆菌EPS的含量及化学成分的影响
由图3可以看出,添加5’-CMP和5’-AMP后,EPS的拉曼光谱峰强度增加。通过比对拉曼光谱特征峰[19]可以看出,干酪乳杆菌所产EPS的主要成分为蛋白质、碳水化合物、核酸以及脂质。添加5’-CMP和5’-AMP后,由EPS拉曼光谱图可以看出,在544~553 cm-1处对应的碳水化合物分泌量增加;720~730 cm-1处对应的物质为腺嘌呤、色氨酸,添加5’-CMP和5’-AMP后细菌产生的这些物质含量也有所增加。在1 001~1 005 cm-1处的拉曼光谱条带主要是苯丙氨酸;在1 200~1 290、1 300 cm-1的拉曼光谱峰显示EPS中含有酰胺物质,添加5’-CMP和5’-AMP后此处的拉曼光谱峰强度增加,表明这两种物质对EPS中酰胺III的产生具有刺激作用。1 605~1 620 cm-1和2 920~1 855 cm-1的拉曼光谱特征峰显示,细菌所产的EPS中含有酪氨酸、色氨酸、苯丙氨酸、酰胺I、不饱和脂质以及孢粉素等物质,这些物质的变形振动主要由C=C、CH2、CH3基团引起。添加5’-CMP和5’-AMP后拉曼光谱显示对应的物质分泌量明显增加。因此,外源核苷酸可以明显增加干酪乳杆菌分泌EPS的量。
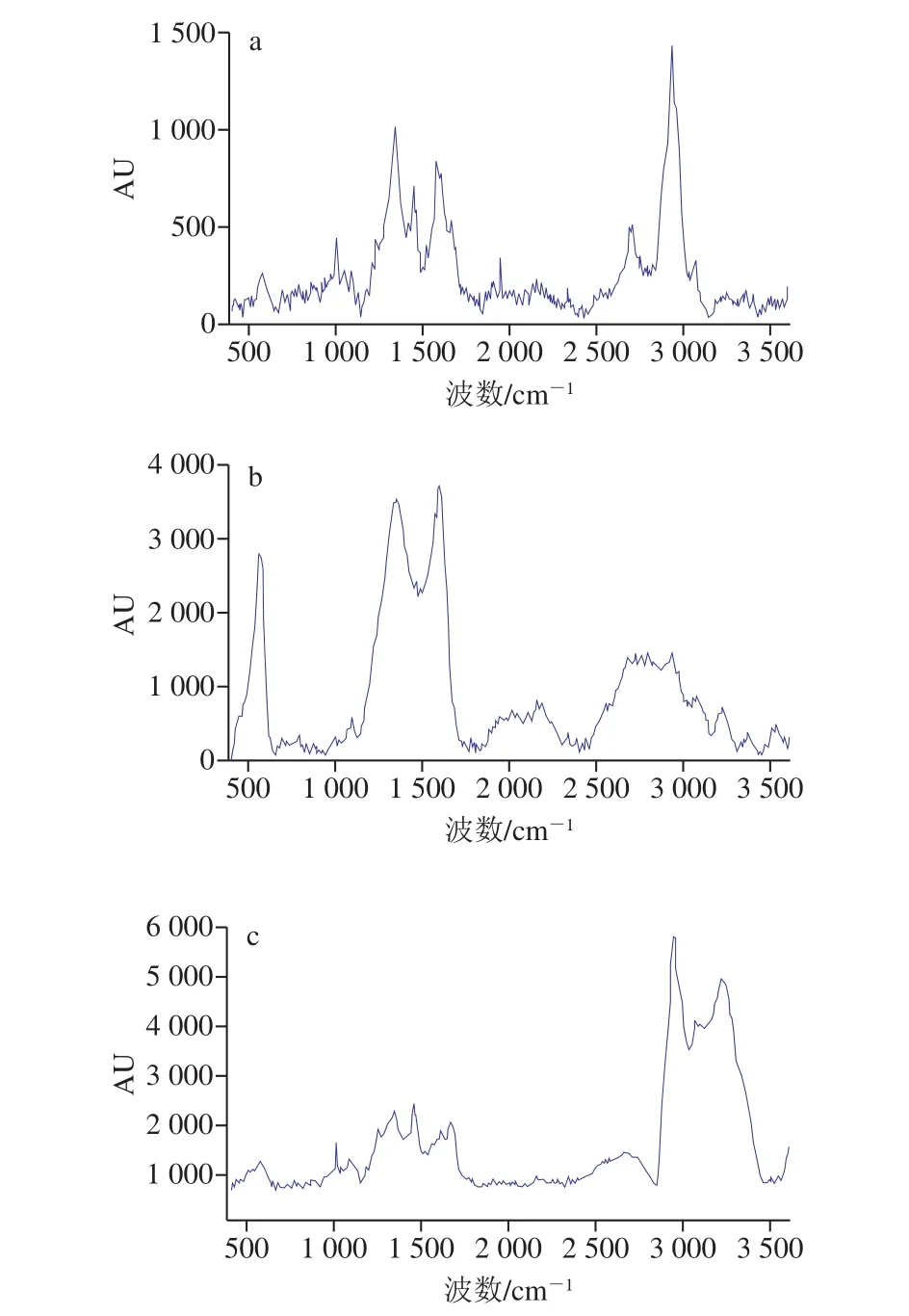
图3 外源核苷酸对干酪乳杆菌EPS含量及化学成分的影响Fig.3 Effect of exogenous nucleotides on the content and chemical composition of EPS produced by L.casei
2.4 外源核苷酸对干酪乳杆菌粗提物群体感应抑制活性的影响
紫色色杆菌CV026是一种群体感应报告菌株,此菌自身不能产生信号分子,但是遇到外源的短链信号分子时,能产生紫色杆菌素使平板变紫色[20]。干酪乳杆菌粗提物的最小抑菌浓度为2 mg/mL。从图4可以看出,添加C6-HSL的平板呈现紫色,当孔中注入亚抑菌浓度下的干酪乳杆菌粗提物时,孔周围出现不透明的黄色晕圈,表明干酪乳杆菌粗提物具有群体感应抑制活性。干酪乳杆菌的培养基添加外源核苷酸(5’-CMP和5’-AMP)后,不透明的黄色晕圈增大,表明细菌分泌的具有群体感应抑制活性的物质增多,尤其是5’-AMP的促进作用更明显,上述结果表明外源核苷酸具有刺激干酪乳杆菌产生群体感应抑制剂的作用。
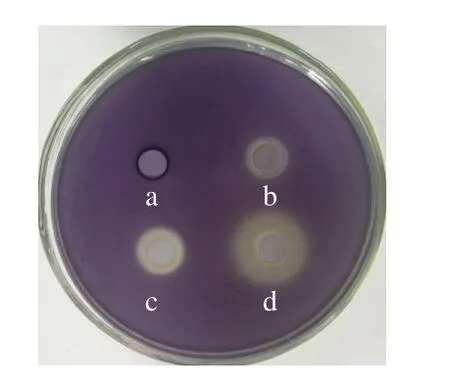
图4 5’-CMP和5’-AMP对干酪乳杆菌粗提物的群体感应抑制活性影响Fig.4 Effect of exogenous 5’-CMP and 5’-AMP on quorum sensing inhibitory activity of crude extract of L.casei Q1
2.5 外源核苷酸对干酪乳杆菌粗提物抗生物被膜能力的影响
从图5可以看出,添加亚抑菌浓度下的干酪乳杆菌粗提物后,志贺菌分泌的生物被膜含量减少。外源核苷酸加入干酪乳杆菌培养基后,其粗提物对志贺菌生物被膜的抑制作用明显增加,5’-AMP的添加对粗提物抗生物被膜活性促进作用更显著。以上结果表明外源核苷酸的添加增加了干酪乳杆菌抗生物被膜物质的产生。

图5 外源核苷酸对干酪乳杆菌粗提物抗生物被膜活性的影响Fig.5 Effect of exogenous nucleotides on anti-biofilm activity of crude extract of L.casei Q1
2.6 蛋白酶和温度对干酪乳杆菌粗提物群体感应抑制活性的影响
从图6可以看出,干酪乳杆菌粗提物经4 种酶处理后,群体感应抑制活性消失,表明粗提物对蛋白酶敏感,因此粗提物中的群体感应抑制活性物质推测为蛋白质类物质。用高温处理干酪乳杆菌粗提物后,其群体感应抑制活性不变,表明粗提物中的群体感应抑制活性物质具有良好的热稳定性。
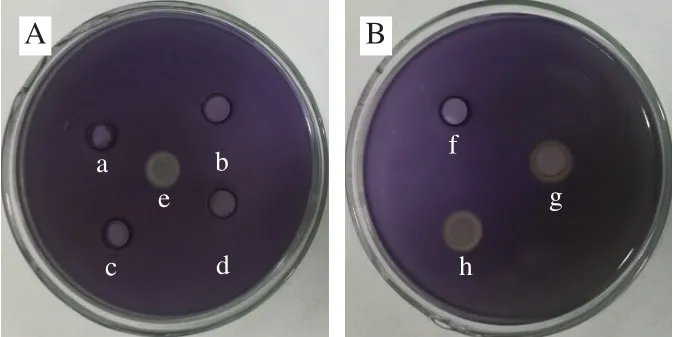
图6 蛋白酶(A)和温度(B)对干酪乳杆菌粗提物群体感应抑制活性的影响Fig.6 Effects of protease (A) and temperature (B) on the quorum sensing inhibitory activity of crude extract of L.casei Q1
3 讨 论
干酪乳杆菌具有的多种对人体有益的功效已被证实[21-23]。但是由于易失活的特点也限制了其应用。因此,外源核苷酸这种安全性好且能促进益生菌生长的物质受到越来越多关注。本研究将外源核苷酸(5’-CMP和5’-AMP)添加到干酪乳杆菌的培养基后,细菌的最大比生长速率提高,延滞期缩短,表明这两种外源核苷酸对细菌的增殖及生长代谢具有促进作用。
细菌产生的生物被膜是细菌为了适应环境,黏附于固体或有机腔道表面,形成微菌落,并分泌EPS将自身包裹其中而形成的膜状物[24]。生物被膜和EPS均受到细菌群体感应系统的调控[25-26]。群体感应是一种细菌之间通过分泌信号分子而进行通讯的机制。信号分子具有多种类型,大多数革兰氏阴性菌由高丝氨酸内酯类作为信号分子;革兰氏阳性菌由寡肽作为信号分子。此外,细菌种间(内)信息交流则由呋喃硼酸二酯类介导[27-30]。添加外源核苷酸后,干酪乳杆菌分泌的生物被膜和EPS含量都明显增加,表明外源核苷酸很可能促进了干酪乳杆菌群体感应系统,从而导致生物被膜和EPS含量增加,使得细菌对外界不利环境的抵抗能力增加。对干酪乳杆菌粗提物的群体感应抑制活性测定也表明,外源核苷酸的添加刺激了干酪乳杆菌分泌具有群体感应抑制活性物质的产生。干酪乳杆菌的粗提物对蛋白酶敏感,因此粗提物中的群体感应抑制活性物质推测为蛋白质类物质。用高温处理干酪乳杆菌粗提物后,其群体感应抑制活性不变,表明粗提物中的群体感应抑制活性物质具有良好的热稳定性,初步推断此物质为细菌素类物质。Chahad等[31]发现,从海水鱼中分离的乳酸菌对多种致病菌具有拮抗性,其粗提物热处理后仍具有抑菌性,经鉴定此物质为II型乳酸菌素。Campos等[32]也发现分离于大菱鲆的乳酸菌可以产生细菌素抑制多种病原微生物。
志贺菌属即通称的痢疾杆菌,细菌性痢疾是最常见的肠道传染病,夏秋两季患者最多[33-34]。传染源主要为病人和带菌者通过污染了痢疾杆菌的食物、饮水等经口感染。志贺菌侵入肠黏膜组织并释放内毒素引起症状,症状为剧烈腹痛、腹泻、发热,严重者出现痉挛和休克[35]。由于抗生素的滥用,造成志贺菌产生了耐药性,因此,寻找新的抗菌靶点或开发新型抗菌药物显得十分重要和迫切。而以上实验结果表明,外源核苷酸可以刺激干酪乳杆菌分泌对抗志贺菌等食源致病菌的物质,这一发现也表明外源核苷酸可作为优质益生元使用,对提高益生菌活力、减少肠道致病菌感染、调节肠道菌群具有巨大的潜力。
4 结 论
本研究发现外源核苷酸(5’-CMP和5’-AMP)对干酪乳杆菌的生长、生物被膜、EPS产生、粗提物的群体感应抑制活性、抗生物被膜活性均有明显的促进作用。2 种外源核苷酸很可能促进了干酪乳杆菌的群体感应系统,从而增强了细菌对环境的抵抗力所致,因此5’-CMP和5’-AMP可作为优质益生元使用,对增强市售益生菌产品的存活率、调节肠道菌群、维护肠道健康具有重要意义。
