低氧胁迫对鱼肉品质影响机制的研究进展
2021-12-02赵曼曼熊光权孙卫青吴文锦丁安子
赵曼曼,熊光权,石 柳,孙卫青,乔 宇,吴文锦,丁安子,李 新,汪 兰
(1.湖北省农业科学院农产品加工与核农技术研究所,湖北 武汉 430064;2.长江大学生命科学学院,湖北 荆州 434025)
水中溶氧量是鱼类生存的基础,不同鱼类对氧气的需求量有所不同,适量的溶氧有助于鱼类的生长发育,但当水中溶解氧(dissolved oxygen,DO)质量浓度低于2 mg/L时,鱼类处于低氧环境[1]。低氧胁迫致使鱼类发生一系列复杂的理化反应,如提高血红蛋白载氧能力、增加红细胞数量、改变代谢方式以及降低代谢率等,低氧胁迫也会导致鱼类行为、能量代谢和肌肉品质发生改变[2-3]。
当水中溶氧充足时,鱼类可将体内糖、脂、氨基酸氧化释放的能量通过呼吸链合成三磷酸腺苷(adenosine triphosphate,ATP),为机体提供大量能量[4-5]。当水中溶氧量低时,鱼类主要通过厌氧糖酵解产生能量和乳酸,而这种异常的能量代谢致使鱼类生长缓慢、死亡率增加[6-7]。低氧使鱼体内活性氧(reactive oxygen species,ROS)水平急剧增加,过多的ROS会诱导脂质过氧化,脂质过氧化过程中产生的小分子醛、酮则会显著降低鱼肉风味,ROS还会诱导蛋白质氧化,并通过降解必需氨基酸、降低蛋白质消化率,影响鱼肉营养价值[8-10]。
本文综述低氧胁迫对鱼类生长发育、氧化应激、能量代谢以及肌肉品质的影响,深入阐明低氧胁迫通过氧化应激、能量代谢对鱼类肌肉品质的影响机制,以期为优质原料鱼肉生产提供理论指导。
1 低氧胁迫下鱼类生长发育和氧化应激变化
1.1 低氧胁迫对鱼类生长发育的影响
鱼类生长发育受内在因素和外在因素的共同影响。内在因素主要指调控肌肉生长发育的基因,如生肌调节基因(myogenic regulatory factors,MRFs)在鱼体胚胎发育及孵化前促使肌原细胞分化、形成肌肉组织,在鱼体胚胎发育后期,使肌纤维体积增大,分化为不同类型肌纤维或肌肉组织[11]。外部因素主要指水生环境,如Gamperl等[12]研究水体温度和溶氧对大西洋鲑鱼的影响,发现温度为12 ℃、中度缺氧时会降低鲑鱼的生长速率。
水生环境与陆地环境有很大不同,水生环境极易受到自然和人为因素的破坏,温度升降、污水排放、放养密度增加等[13]均会使水中的溶氧量处于不断变化的状态。通常对鱼类生长发育来说,水中溶氧量保持在4 mg/L以上,鱼类能正常生长,而溶氧量低于2 mg/L时,鱼类出现生长发育迟缓现象[14]。例如当水体溶氧量为1.5 mg/L时,大西洋鲱鱼的生长速率降低了31%~89%[15]。即使是耐低氧的鱼类,处在溶氧量不断变化的水环境中,也会产生应激反应。应激反应是一个耗能过程,鱼类为了存活会抑制体内生长发育相关的代谢机制。Hou Zhishuai等[16]研究发现处在低氧环境虹鳟鱼的生长激素-胰岛素样生长因子(growth hormone-insulin-like growth factor,GHIGF)轴受到抑制,生长激素可以激活位于肝脏、肌肉和骨骼上的受体,进而触发胰岛素样生长因子系统,胰岛素样生长因子可充当中枢系统调节内分泌过程,并激活细胞内信号传导通路,抑制骨骼发育和肌肉细胞的生长、增殖和分化。低氧还会激活肿瘤坏死因子(tumor necrosis factor,TNF)的转录,新合成的TNF蛋白从肝细胞分泌并激活位于骨细胞膜上的受体,造成破骨细胞代谢紊乱,骨基质降解、钙动态异常,肌肉生长受到抑制。Wang等[17]研究发现低氧胁迫可通过干扰能量代谢机制如鱼类由有氧代谢转变为无氧糖酵解,而能量代谢紊乱使鱼类食欲不振、生长迟缓。综上所述,低氧环境中的鱼类,通过影响肌肉生长发育基因的表达以及抑制相关代谢机制,致使生长发育迟缓、体型偏小。
1.2 低氧胁迫对鱼类氧化应激的影响
脂质过氧化的中间产物自由基损害细胞膜结构、氧化含巯基的氨基酸,致使生物体细胞损伤、蛋白质合成降低[18]。脂质过氧化终产物(如丙二醛(malondialdehyde,MDA))常用来反映鱼类氧化应激的程度[19]。Wang Man等[20]研究发现低氧组((1.14±0.04)mg/L)黄颡鱼肝脏中MDA含量显著高于对照组((6.72±0.17)mg/L)组,这表明MDA含量越高,氧化应激程度越高。蛋白质氧化主要是活性氧直接攻击蛋白质或脂质过氧化间接诱导引起蛋白质结构的变化,鱼体内蛋白质氧化后活性巯基降低、羰基含量增加、蛋白质合成量降低。氧化应激不仅降低鱼体内蛋白质合成还会增强某些蛋白质的表达。Dawood等[21]研究发现低氧条件下,罗非鱼体内免疫球蛋白、热休克蛋白(heat-shock proteins,HSPs)表达量增加,用以缓解体内氧化损伤。
一般来讲,自身抗氧化体系以及外源添加剂均可缓解鱼类氧化损伤。体内抗氧化体系主要由抗氧化酶(如超氧化物歧化酶、谷胱甘肽过氧化物酶)和非酶类抗氧化剂(维生素、谷胱甘肽)组成[22]。超氧化物歧化酶是降解活性氧自由基的第一道防线,可将超氧阴离子自由基转化为过氧化氢并在过氧化氢酶的作用下生成水和氧气;谷胱甘肽过氧化物酶可清除胞内过氧化氢和有机过氧化物,以缓解氧化应激反应;抗氧化酶的活性可以用来表征鱼类氧化应激状态[23-25]。Nam等[26]研究红鲷时发现,短期缺氧红鲷体内超氧化物歧化酶和过氧化氢酶活性显著升高,这表明体内抗氧化酶有助于缓解氧化应激带来的损害。外源添加剂主要通过改变鱼类日粮组分来缓解氧化损伤。Abarike等[27]向日粮中添加中草药(黄芪、当归和胡杨),缓解了罗非鱼的低氧损伤。Li Huatao等[28]向日粮中添加含有蛋白质和多糖的桑叶提取物,减轻了鱼消化器官和呼吸器官中由缺氧和复氧引起的氧化损伤。由此可知,低氧胁迫致使鱼类产生氧化应激反应,损害体内脂质、蛋白质组成,而体内抗氧化体系以及外源添加剂可缓解氧化损伤,并伴随着抗氧化酶活力的增加以及免疫相关蛋白表达量的上调。
2 低氧胁迫对鱼体能量物质代谢的影响
2.1 糖代谢
短期或轻度低氧胁迫,糖类是最有效的能量供应物,主要以糖酵解、糖原分解以及糖异生方式产能。鱼类在应对低氧胁迫时,首先激活体内低氧诱导因子(hypoxia inducible factor,HIF)信号通路,其中HIF-1α、脯氨酸羟化酶(prolyl hydroxylase domain enzyme,PHD)、肿瘤抑制蛋白(von Hippel-Lindau tumor suppressor protein,pVHL)是HIF信号通路的关键信号分子。低氧条件下,PHD蛋白活性受到抑制,HIF-1α在细胞体内终止降解并聚集,随后与细胞核内的HIF-1β形成具有转录功能的异源二聚体[29],诱导下游低氧调控基因如葡萄糖转运子(glucose transporters,GLUTs)以及与糖酵解相关的酶类如己糖激酶(hexokinase,HK)、乳酸脱氢酶(lactate dehydrogenase,LDH)和丙酮酸激酶(pyruvate kinase,PK)[30]。低氧胁迫还会激活腺苷酸活化蛋白激酶(adenosine 5’-monophosphate (AMP)-activated protein kinase,AMPK)信号通路(图1)[31],通过AMPK信号通路感知肌肉细胞能量变化,增加葡萄糖摄取、糖酵解、糖异生作用。糖酵解过程中产生大量乳酸,可通过单羧酸转运蛋白(monocarboxylate transporter,MCT)途径转运到血液中,避免肌肉酸中毒。研究表明,AMPK在调控动物能量代谢和激活HIF-1转录活性方面具有十分重要的作用[32-33]。
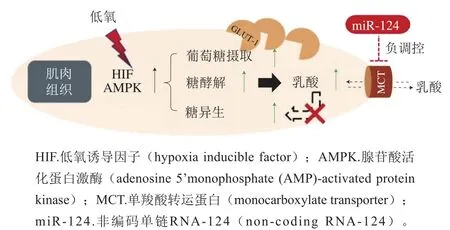
图1 大口黑鲈肌肉中糖代谢及乳酸转运[31]Fig. 1 Glucose metabolism and lactic acid transport in the muscle of largemouth bass[31]
低氧胁迫会使鱼类代谢方式从有氧代谢转变为无氧糖酵解,即葡萄糖在无氧条件下分解为丙酮酸,丙酮酸在乳酸脱氢酶的作用下生成乳酸并产生少量能量[34-35]。低氧胁迫会增强鱼类糖原分解和抑制糖原合成,以满足机体基本能量需求,并保持血糖水平稳定。短期低氧胁迫会导致糖原和血糖含量降低[36],长期低氧胁迫使鱼类消耗大量能量,糖异生作用增加、体内葡萄糖含量升高[37]。研究发现,低氧胁迫大黄鱼6、24、48、96 h后,与糖酵解有关的酶如HK、磷酸果糖激酶(phosphofructokinase,PFK)以及与糖异生相关的酶如磷酸烯醇式丙酮酸羧激酶(phosphoenolpyruvate carboxykinase,PEPCK)、葡萄糖-6-磷酸酶(glucose-6-phosphatase,G-6-Pase)等均显著上调[38]。
2.2 脂代谢
鱼类在长期或急性缺氧的情况下,脂质代谢便会成为主要的能量代谢方式,且以脂质的分解代谢为主。研究发现大口黑鲈在低氧应激8 h后,甘油三酯(triglyceride,TG)、游离脂肪酸(nonesterified fatty acid,NEFA)含量显著增加,参与脂解的酶类如脂肪酶(lipase,LPS)和脂蛋白脂肪酶(lipoprotein lipase,LPL)活性先升高后降低[39]。这可能由于鱼类低氧胁迫时,TG在LPS和LPL作用下进一步分解为NEFA,NEFA含量的增加抑制TG的分解[40]。
低氧胁迫通过激活过氧化物酶体增殖剂激活受体(peroxisome proliferators-activated receptors,PPARs)信号途径。PPARs包括PPARA、PPAR β/δ、PPARγ,其中PPARA可参与调节脂质代谢和维持葡萄糖稳态。如图2[41]所示,一方面,急性低氧增加大口黑鲈肝脏中脂肪分解代谢相关酶类活性(LPS和LPL),同时显著提高脂肪动员和脂肪酸转运有关基因的表达水平,包括PPARA、溶质载体家族成员27成员6(solute carrier family member 27 member 6,SLC27A6)。LPS和LPL在肝脏中将TG分解为NEFA,PPARA及SLC27A6将NEFA释放到血液中,继而被机体其他组织氧化供能。另一方面,急性低氧胁迫会显著降低肉碱棕榈酰转移酶1(carnitine palmitoyltransferase 1,CPT-1)的活性,CPT-1酶是脂肪酸β-氧化的关键速率限制酶,说明低氧胁迫会抑制脂肪酸的β-氧化[42]。

图2 急性缺氧对大口黑鲈肝脏脂质代谢相关酶和基因的影响[41]Fig. 2 Effect of acute hypoxia on enzymes and genes associated with lipid metabolism in liver of largemouth bass[41]
2.3 蛋白质代谢
鱼类在严重缺氧时,无法通过氧化磷酸化产能,为了维持体内能量平衡,可以通过转氨基(如谷氨酸、精氨酸和脯氨酸)作用将一些特定氨基酸转化为丙氨酸并生成ATP,还可以通过抑制非必须基因的翻译以及蛋白质的加工以节约能耗[43]。Tian Changxu等[44]研究发现,多鳞鱚经过4 h缺氧处理后,tRNA连接酶、核糖体蛋白(ribosomal protein,Rp)的表达量降低。tRNA连接酶是调节翻译过程的关键酶,核糖体蛋白是核糖体的主要成分,调控细胞内蛋白质的合成,同时蛋白质在内质网中的加工和输出也受到抑制,这表明低氧会抑制基因的翻译和蛋白质合成、加工的过程[45]。
低氧胁迫对鱼类蛋白质代谢具有一定影响。鱼类应对低氧环境时,会促进蛋白质的分解,且减少蛋白质的合成。在胃肠道消化酶的作用下,蛋白质被分解成游离氨基酸及小分子肽(二肽、三肽)。游离氨基酸在转氨酶的作用下进行转氨基和脱氨基反应,产生的代谢物如丙酮酸可进一步参与糖代谢过程。Moraes等[46]发现低氧胁迫刀鲚3 h后,体内游离氨基酸水平显著降低。Genin等[47]发现作为蛋白质分子伴侣的HSPs,在缺氧时基因表达高度上调。董云伟等[48]研究发现热休克蛋白不仅可以降解鱼体内变性蛋白质、防止细胞自溶,还可用来评估水产养殖中鱼类的健康状况。
各土地利用现状图按照第二次土地利用调查分类标准分为二级地类,分别为:耕地、园地、林地、草地、林地、交通运输用地、水域及水利设施用地、城镇村及工矿用地、其他用地。在以往的研究基础上,同时估计研究区面积及景观破碎程度,基于相关软件平台将土地利用矢量数据输出为100m×100m的栅格数据,作为生境质量的评价单元。
3 低氧胁迫对鱼肉品质的影响
鱼肉品质主要包括食用品质、加工品质以及营养品质等。低氧胁迫使鱼类糖、脂质、蛋白质等物质能量代谢发生显著变化,物质能量代谢转变过程中产生的中间物质及代谢产物,如相关酶类和蛋白以及乳酸等,会致使鱼肉色泽、嫩度、持水性下降并产生不良风味。有关低氧胁迫过程中鱼肉品质与糖、脂质、蛋白质物质代谢关系如表1所示。

表1 低氧胁迫下鱼肉品质和糖、脂质、蛋白质物质代谢关系Table 1 Relationship between fish meat quality and glucose, lipid and protein metabolism under hypoxia stress
3.1 低氧对鱼肉食用品质的影响
3.1.1 色泽
色泽是影响消费者购买欲的直接因素,也是判断肉质是否劣变的重要标准。肌肉的色泽主要与血红素蛋白中肌红蛋白的形态和含量有关。鱼类在缺氧时,会加速脂质分解并促进脂质过氧化,而脂质过氧化的初级和次级产物对肌红蛋白的存在形式有显著影响[54-55]。铁存在于血红蛋白和肌红蛋白的血红素中,而转铁蛋白有助于维持细胞内铁的稳态。Chen Boxiang等[49]研究发现,低氧条件(0.5 mg/mL)会增加团头鲂肌红蛋白表达量。鱼肉中红肌和白肌的比例是影响肉色的另一因素,急性和慢性低氧均会影响红肌和白肌的代谢及占比,这在虹鳟中已经得到验证[56]。目前,最常用的肉色测定方法是用色彩色差仪测定L*、a*、b*值,其中L*值表征亮度,a*值表征红度,b*值表征黄度。Lefèvre等[57]对虹鳟屠宰前进行短期低氧处理,发现鱼肉亮度和黄度显著降低。
3.1.2 嫩度
肉的嫩度是评价鱼肉食用品质的另一重要指标。低氧胁迫对鱼肉嫩度的影响与缺氧程度相关:轻度低氧胁迫时,鱼体内活性氧增加有助于蛋白质降解,从而提高鱼肉嫩度;重度低氧胁迫时,蛋白质分子内或分子间发生交联聚合,抑制蛋白质的降解,降低鱼肉嫩度。Ciaramella等[50]研究发现,轻度低氧胁迫下,斑点叉尾鮰的α-肌动蛋白3以及肌球蛋白α-1表达量下降,证明轻度低氧胁迫可使结构性蛋白降解,提高鱼肉嫩度;重度低氧胁迫下,与能量代谢有关的AMP脱氨酶表达量升高,而AMP脱氨酶可与肌原纤维蛋白结合,抑制肌原纤维蛋白的降解,降低鱼肉嫩度。
3.1.3 风味
肉的风味可作为评判肉是否新鲜的标准,通常包括气味和滋味。气味主要是由小分子的醛、醇、酮、酯等挥发性物质组成,低氧胁迫促使蛋白质氧化以及脂质过氧化,蛋白质氧化过程中产生的酮类以及脂质过氧化过程中产生的醛、酮对鱼的气味有显著不良影响。滋味主要由游离氨基酸、核苷酸、有机酸等非挥发性物质构成,低氧胁迫会促使鱼类糖代谢发生转变,无氧糖酵解能力增强产生大量乳酸,而氨基酸与乳酸等有机酸结合会在肉中产生酸味使肉质发生劣变[58]。
3.2 低氧对鱼肉加工品质的影响
肉类加工品质主要包括硬度、pH、系水力等。低氧胁迫一方面显著增加氨基酸分解,抑制肌肉胶原蛋白合成,胶原蛋白合成降低可能会改善鱼肉硬度。另一方面,肌细胞生成素(myogenin,MyoG)表达量显著增加,而MyoG的表达量与肌肉硬度呈正相关[51]。总之,低氧胁迫使鱼肉硬度增加,这可能因为低氧条件下MyoG对鱼肉硬度的影响更大。
pH值下降主要是糖酵解过程中鱼肉乳酸的积累所致,Pédron等[52]研究发现葡萄牙利马河口中的比目鱼在低氧条件下会增强肌肉中糖酵解能力(如乳酸脱氢酶的活性),抑制有氧呼吸过程(如柠檬酸合成酶、细胞色素氧化酶)。pH值下降会促使细胞膜离子通道发生改变,细胞内汁液流失、肌红蛋白和肌原纤维蛋白含量降低,鱼肉的持水性、色泽、剪切力发生显著劣变,这在虹鳟中已经得到验证[59]。
持水性是评估肉质的重要指标,通常用蒸煮损失、滴水损失表征。鱼肉的系水力受自身因素以及外部环境的影响。内在因素如细胞体积的变化、蛋白质氧化以及pH值下降均会影响鱼肉的持水性。Wulff等[53]发现低氧条件下,虹鳟肌肉中参与细胞骨架调节的微管蛋白a链、肌球蛋白结合蛋白h以及参与细胞体积调节的膜突蛋白的基因上调,且有学者发现缺氧会导致细胞肿胀,进而造成水分流失[60]。蛋白质氧化会抑制钙激活中性蛋白酶(Calpains)活性,降低肌肉持水力[61]。pH值下降主要由于肌肉中乳酸的积累,Bjrnevik等[62]研究发现低氧胁迫下鳕鱼持水力显著下降。外部环境的影响包括日粮影响等,Wu Pei等[63]研究发现鱼类日粮中缺乏组氨酸时,鱼肉剪切力、pH值降低,而蒸煮损失增加。综上可知,低氧环境中鱼体内糖代谢以及蛋白质结构的变化,使鱼肉硬度增加、pH值降低以及持水性下降,影响肉的多汁性、嫩度和色泽,降低鱼类加工品质。
3.3 低氧对鱼肉营养品质的影响
营养品质一般包括粗蛋白、粗脂肪、维生素、矿物质等。鱼类长期处于低氧环境,脂代谢发生改变,抑制脂肪合成同时促进脂肪分解,脂肪分解产物TG可以经过一系列转化途径参与糖酵解过程为机体供能,脂肪分解使体内粗脂肪含量降低。Abdel-Tawwab等[64]将罗非鱼在低氧(0.5~1.0 mg/L)养殖12 周,与常氧组相比(6.0~6.5 mg/L),鱼体内总脂肪含量下降,这表明低氧可能影响鱼体内营养物质的沉积,进而对鱼肉品质产生影响。但也有学者发现低密度养殖的黄花鱼在低氧处理后其肝脏代谢产物如葡萄糖、糖原、甘油三酯等与未经低氧处理组相比并无显著差异[65]。低氧胁迫还会促使脂质过氧化,而脂质过氧化产物如丙二醛、4-羟基壬烯酸与生物分子结合改变细胞膜的流动性和通透性,损伤细胞结构和功能,导致营养物质流失。鱼类在急性缺氧时,谷氨酸可通过转氨基作用,产生少量能量,鱼体内蛋白质表达降低。Bosworth等[66]研究斑马鱼的蛋白质组学,发现急性缺氧会使蛋白质的丰度降低,这表明低氧可能会影响蛋白质的整体表达,鱼体内蛋白质含量降低。低氧胁迫还会导致蛋白质氧化,必需氨基酸降解,鱼体内蛋白质含量降低。综上可知,低氧环境鱼体内脂肪以及蛋白质含量下降均会影响鱼肉营养品质。
4 结 语
鱼类在养殖、运输等过程中不可避免遭受低氧胁迫,低氧胁迫调控鱼类生长激素-胰岛素样生长因子并抑制能量代谢机制,供自身生长发育。鱼类在低氧环境中产生氧化应激反应、能量代谢异常,氧化应激使鱼体内脂质过氧化、蛋白质氧化,降低鱼肉食用、加工、营养品质,而糖、脂、蛋白质代谢异常,进一步促进肉质劣变。目前,国内外学者对鱼类应激方面做了大量研究,但都集中在应激对鱼类存活率、生理生化、缓解方法的变化机制,未来研究应致力于将内在机制变化与鱼肉品质相结合,全面建立低氧胁迫对鱼肉品质影响的内在调控网络,保证鱼肉品质,提高水产经济。
