传统牦牛酸奶源高产胞外多糖乳酸菌特性及发酵性能
2021-11-23降初祝玛陈炼红
降初祝玛,陈炼红,张 岩
(西南民族大学食品科学与技术学院,四川成都 610041)
传统的牦牛奶是藏区农牧民必不可少的饮品,牦牛奶中含有多种营养物质,且含量高于普通牛奶,牦牛奶经发酵后,具有抗氧化、降胆固醇等保健功能[1],营养物质也随之发生变化,具有较为丰富的微生物菌相,是获取具有优良发酵性能乳酸菌的主要来源[2]。乳酸菌(Lactic acid bacteria, LAB)是公认的安全的食品级微生物[3],其次级代谢产物胞外多糖(Exopolysaccharide, EPS)是乳酸菌在生长过程代谢中分泌到细胞壁外或渗入培养基中的一种多糖及其混合物[4],本身具有保护菌体[5]、抑制癌细胞增殖[6]、调节免疫能力等生理功能[7]。此外,研究表明EPS乳酸菌对酸奶流变学特性[8],组织状态、质地和黏稠度等有益[9−10]。
近年来,胞外多糖乳酸菌可用于食品工业和医药领域,具有潜在的开发价值,其发酵性能也是研究的热点[11]。目前我国对牦牛乳源高产胞外多糖乳酸菌研究不多,局限于对其筛选鉴定和益生功能与耐受力的探讨,如唐超群等[12]从牧民自制的牦牛乳制品(奶渣)中分离得到胞外多糖乳酸菌。王翔宇等[13]研究了传统发酵牦牛乳中2 株高产胞外多糖乳酸菌在模拟消化道中耐受力。从未对川西高原发酵牦牛酸乳中筛选出的高产胞外多糖乳酸菌进行其菌株特性和发酵性能的研究。
本实验运用培养高产胞外多糖乳酸菌菌株在其生长达到最佳收获时间,获得菌体,对菌株进行耐不良环境能力的实验,再利用菌体发酵酸奶,测定酸奶的发酵性能。本研究对于开发青藏高原极端环境条件下的发酵剂种质资源,生产功能性乳制品、丰富现有乳制品种类、提高乳制品的附加值将具有重要的意义。
1 材料与方法
1.1 材料与仪器
6 株高产EPS 乳酸菌 均分离自四川藏区传统牦牛酸奶,经过16S rDNA 测序鉴定,其鉴定结果和来源地如表1。由西南民族大学食品科学与技术学院保藏。

表1 菌株来源Table 1 Source of strains
巴氏杀菌鲜奶、蔗糖 购买于沃尔玛超市;M17 固体培养基、MRS 肉汤培养基 青岛海博生物技术有限公司;氯化钠、牛胆盐、盐酸、十六烷、二甲苯、氢氧化钠、三氯乙酸、邻苯二胺 均为分析纯,成都康迪生物技术有限公司;酚酞指示剂、双乙酰标准液、碘液、PBS 磷酸缓冲盐溶液 成都创诺飞生物科技有限公司。
PYX﹣DHS﹣50X65﹣BS﹣Ⅱ型隔水式电热恒温培养箱 北京中科兰泽科技有限公司;PHS-3C 型pH 计 上海雷磁仪器厂;HC-2062 型高速离心机安徽中科中佳科学仪器有限公司;UV-2100 型分光光度计 尤尼柯仪器有限公司;HH-数显恒温水浴锅
国华电器有限公司;PL303 型分析天平 梅特勒托利多仪器有限公司;TA-XT.Plus 质构分析仪 英国Stable Micro Systems 公司。
1.2 实验方法
1.2.1 菌株特性
1.2.1.1 菌株的生长曲线 将冷冻保存的六株高产胞外多糖乳酸菌菌株进行解冻,混匀后吸取100 μL菌液置于5 mL 液体培养基中传三代进行活化,使得菌种充分恢复活性,最后按照3%的接种量接种在MRS 液体培养基,培养。
乳酸菌生长曲线的测定:将充分活化的菌株按3%的接种量接入5 mL MRS 液体培养基中,37 ℃厌氧培养24 h,在0 h 开始取样,每隔2 h 取样,用没有接过菌种的MRS 液体培养基作为空白调零,测定OD600值,三组平行求平均值[14]。
1.2.1.2 菌株的耐酸性 用盐酸调节MRS 液体培养基pH 为1.5、2.5、3.5、4.5、5.5,接上3%的菌株培养液,37 ℃厌氧培养,在24 h 时取样,用没有接过菌液的对应pH 的MRS 液体培养基作为空白调零,测定OD600值,三组平行求平均值[15]。
1.2.1.3 菌株的耐胆盐 按菌株3%的接种量分别接到牛胆盐浓度分别为0.05%、0.1%、0.5%、1.0%、1.5%已灭菌MRS 液体培养基中,37 ℃厌氧培养,在24 h 时取样,用没有接过菌液相对应的牛胆盐浓度MRS 液体培养基作为空白调零,测定OD600值,三组平行求平均值[16]。
1.2.1.4 菌株的耐渗透压 按菌株3%的接种量分别接到NaCl 质量分数分别为3%、4%、5%、6%、7%已灭菌 MRS 液体培养基中,37 ℃厌氧培养,在24 h 时取样,用没有接过菌液相对应 NaCl 质量分数MRS 液体培养基作为空白调零,测定OD600值,三组平行求平均值[17]。
1.2.1.5 菌株的细胞表面疏水性 PBS(磷酸盐缓冲液):0.2 mol/L Na2HPO4·12H2O 18.5 mL+ 0.2 mol/L NaH2PO4·2H2O 81.5 mL,用蒸馏水稀释1 倍,PBS pH=6.2±0.2。
菌体重悬液的制备:将六株乳酸菌的菌株培养液分别在6000 r/min,4 ℃条件下离心10 min,得到菌体,用PBS 洗涤菌体2 次,10 mL/次[18]。PBS 为空白对照,并用PBS 调整菌液的浓度,用分光光度计测定OD600值为0.6±0.02。
菌体表面疏水性的测定:取每管为5 mL 的菌体重悬液到试管中,再往试管中分别加入200 μL 的十六烷和二甲苯,对照组不加,该两相体系通过涡旋彻底混合1 min,静置15 min 分层。取水相,PBS 为空白对照,测量OD600值并记录。每组平行做5 管,分别记录OD 值,并进行3 次独立的实验[17]。通过以下公式计算:
表面疏水性(%)=(对照组OD600−实验组 OD600)/对照组 OD600×100
1.2.2 菌株的发酵性能
1.2.2.1 凝乳时间 用符合国家标准的生鲜牛奶,加入白砂糖,高压均质化,巴氏灭菌30 min,冷却到42 ℃,按照3%比例接种六株乳酸菌的菌体,在42 ℃温度下发酵酸奶至凝乳状态(当发酵到液体全部转化成凝固状,轻微摇晃不改变固体形状时,停止发酵),确定发酵时间,酸奶凝乳后,4 ℃冷藏保存[19]。
1.2.2.2 持水力测定 取发酵酸奶样品10 g 装入离心管中,在5500 r/min 转速下,低温离心30 min,倒掉上清液,倒置10 min,让上清液充分流出,最后立即称量沉淀的质量[20]。计算公式如下:
持水力(%)=离心后沉淀质量/样品质量×100
1.2.2.3 酸化能力 将六株乳酸菌的菌体培养液分别按3%的比例接种到牛奶中,在42 ℃条件下发酵。在发酵过程中2 h 取一次样,测定酸奶的滴定酸度。
1.2.2.4 后酸化能力 六株乳酸菌的菌体培养液分别按3%的比例接种到牛奶后,经42 ℃条件发酵至凝固后放入4 ℃环境中贮藏。测定酸奶在冷藏1、3、5、10、15 d 后的滴定酸度,根据酸奶在贮藏期间滴定酸度的增加量来判断后酸化能力的高低。
1.2.2.5 冷藏期间活菌数的变化 采用稀释平板计数法,在无菌条件下,分别取1 mL 4 ℃冷藏1、3、5、10、15 d 的酸奶样品,在9 mL 的0.9%的NaCl溶液中进行10 倍的梯度稀释,使其混匀后,分别吸取1 mL 10-5、10-6、10-7的样品稀释液于灭菌的培养皿中,倒入20 mL 已灭菌并冷却至45~50 ℃的MRS琼脂培养基,将样品稀释液涂布于凝固后的MRS 琼脂培养基。等培养皿中培养基冷却凝固后,将培养皿倒置放入(36±1)℃恒温培养箱,培养18~24 h。测定酸奶冷藏期间活菌数的变化[19]。
1.2.2.6 发酵酸奶的乙醛含量测定 将8%的TCA溶液与酸奶样品按比例混匀,在5500 r/min 转速下,低温离心10 min,取5 mL 过滤后的上清液,加入1 mL浓度为1%的 NaHSO3溶液振荡摇匀,在室温环境下放置60 min,然后加入0.5 mL 的1%淀粉溶液,用0.1 mol/L 的碘标准溶液进行滴定,快接近终点时换0.01 mol/L 的碘标准溶液滴定到为初滴定终点,(溶液变为淡蓝色,30 s 不褪色),不计数。再加入10 mL的1 mol/L NaHCO3,摇匀到溶液颜色消失,最后用0.01 mol/L 的碘标准溶液滴定到终点,记录消耗的碘标准溶液的体积,同时做空白对照试验[21],三组平行求平均值。
1.2.2.7 发酵酸奶的双乙酰含量测定 参考吕嘉枥等[22]的方法,用0.5%的邻苯二胺与不同浓度的双乙酰标准溶液反应生成2,3-二甲基并吡嗪,在波长为325 nm时有最大吸收值,可对双乙酰含量进行定量测定。三组平行求平均值。
1.2.2.8 酸奶的质构(TPA)测定 质构仪测定样品,重复测定3 次,求其平均值。质构仪测定设置参数:探头:A/BE35;测试距离:25.00 mm;测定前探头速度:2.00 mm/s;测定时探头速度:2.00 mm/s;测定后探头速度:2.00 mm/s;感应力:Auto-10.0 g[23−24]。
1.3 数据处理
采用Microsoft Office Excel 2013 和SPSS 17.0对数据进行统计分析,利用单因素方差分析对数据进行显著性分析,每个试验做三个重复,显著差异水平取P<0.05。
2 结果分析
2.1 生长曲线
由图1 可知,六株乳酸菌菌株的生长在前2 h 内生长量均没有明显变化,经过短时间的迟缓期后,从4 h 开始到了对数生长期,随后菌数呈指数上升。218、276、266 在14 h 进入对数生长稳定期,271、285、231 在16 h 进入对数生长稳定期。其中编号218 的菌株生长在14 h 时OD 值为2.188,是六株菌株中生长量最高的。

图1 乳酸菌在生长期间OD 值的变化曲线Fig.1 The curve of OD value of lactic acid bacteria during growth
2.2 耐受性实验结果分析
2.2.1 耐酸实验的结果分析 人体正常胃酸pH 在3.0 左右,空腹与食用酸性食品时pH 达1.5,而食用碱性食物时,其pH 一般为4.0~5.0[25]。因此,具有较强耐酸能力的乳酸菌才能够顺利通过胃酸屏障,并到达肠道内发挥其益生作用。由表2 可知,随着介质pH 的增大,6 株胞外多糖乳酸菌生长繁殖能力增强,来自酸的抑制作用减弱。当培养介质pH 在2.5~5.5时,同种乳酸菌OD 值有显著差异(P<0.05),说明pH对菌株的生长影响较大;当培养介质pH 在1.5 时,菌株生长被抑制,但仍能够存活和生长,其中编号为276 的菌株OD 值最大(P<0.05),说明该菌株具有良好的耐酸能力。
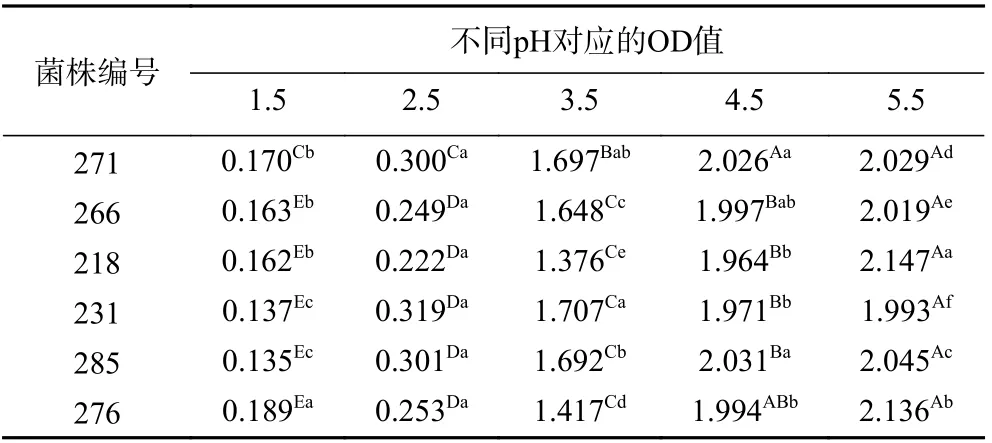
表2 乳酸菌在不同pH 下OD 值的变化Table 2 The change of OD value of lactic acid bacteria under different pH
2.2.2 耐胆盐的结果分析 人体小肠内胆盐的含量一般在0.03%~0.3%范围内波动[26],食物通过肠道的时间一般是4~16 h,乳酸菌的耐胆盐能力也是乳酸菌能否在肠道存活的重要条件。由表3 可知,六株菌株在胆盐浓度为0.05%~0.5%时,生长状态均良好,说明六株菌株在正常人体的胆盐范围内可以存活;六株菌株均在0.5%的浓度下生长达到最高值,其中编号266、218、231 的菌株生长量最优(P<0.05),表现出对高浓度胆盐的耐受性较强;胆盐浓度在1%~1.5%时,六株菌株生长受到抑制,说明这六株菌株不耐高胆盐环境。

表3 乳酸菌在不同胆盐浓度下OD 值的变化Table 3 The change of OD value of lactic acid bacteria under different bile salt concentration
2.2.3 耐渗透压的结果分析 NaCl 在人体内的含量约为1%~4%[27],高盐会导致高渗透压,当渗透压浓度突然变化时,会影响到乳酸菌的生长代谢,严重时会导致乳酸菌的死亡。由表4 可知,6 株乳酸菌OD 值均随着NaCl 浓度的增大而降低。6 株菌株在较低NaCl 浓度(3%、4%)时,生长状态均良好,表明均具有较好的耐渗透压的能力;当添加5% NaCl 浓度时,285 菌株的生长量为最高值(P<0.05),表明菌株285 对高渗透压的耐受性较强。六株菌株在较高NaCl 浓度(6%、7%)时,六株菌株生长均被抑制,菌体处于低生存状态,基本不生长繁殖。

表4 乳酸菌在不同NaCl 浓度下OD 值的变化Table 4 The change of OD value of lactic acid bacteria under different NaCl concentration
2.2.4 细菌表面疏水性的结果分析 对肠道黏附能力是乳酸菌在人体内发挥益生作用的前提,而细菌的表面疏水性与黏附能力呈正相关,因此根据疏水性的大小可筛选出潜在的高粘附性菌株[28]。由图2 可知,通过十六烷和二甲苯处理,不同菌株间的疏水性差异显著(P<0.05),菌株285 和218 在两种有机试剂中的疏水性均显著(P<0.05)高于其他菌株,且与十六烷的结合效果较好。其中编号266 的菌株在十六烷中表面疏水性最低为2.3%。六株乳酸菌的菌株表面疏水性的大小排序为:285>218>231>276>271>266。

图2 乳酸菌的表面疏水性Fig.2 The surface hydrophobicity of lactic acid bacteria
2.3 发酵性能
2.3.1 凝乳时间的结果分析 在生产酸奶的过程中,凝乳时间太短不利于酸奶中芳香物质的形成,使酸奶不香,同时还可能使酸奶凝乳状态不佳;而凝乳时间太长不利于酸奶的工厂生产,成本过高,同时还容易导致产品的污染。六株乳酸菌的菌株发酵酸奶都在6~8 h 内凝固。
2.3.2 持水力的结果分析 酸奶离心后所截留的水分称为持水力(WHC),多为凝胶中大分子物质结合较紧密的水分[29],WHC 越高,该体系中的大分子结合水分子的作用力越强,稳定性也越好。由图3 可知,1~5 d 内,六株菌株的持水力缓慢上升,其中218、231、276 在第5 d 开始迅速上升,在第15 d 达到最高值。菌株218 发酵的酸奶持水力最强,达到了60.98%。这可能是由于产EPS 的乳酸菌发酵乳中含有微孔,EPS 在这些微孔里使蛋白质网络形成更厚更浓密的蛋白质凝胶通道,这种结构更有利于防止发酵乳乳清析出[30],从而增加了发酵乳的持水力。

图3 冷藏期间持水力的变化Fig.3 The change in water holding capacity during cold storage
2.3.3 酸化能力的结果分析 产酸能力是乳酸菌的重要性能,产酸快的乳酸菌有利于生产时发酵时间的缩短,可以提升生产效率,但当产酸过多,酸度增加会破坏酸奶的滋味和凝乳状态,而且高浓度的酸会阻碍乳酸菌的生长,也会影响酸奶的组织结构和品质。由图4 可知,在整个发酵时期,六种酸奶的酸度增长较快,发酵期结束时,6 种酸奶的酸度均已达到70 °T,均符合国标GB 19302-2010 中的对发酵乳中酸度要求(>70 °T)[31],其中编号218 的菌株的酸度最高,为83 °T。

图4 发酵期间滴定酸度的变化Fig.4 The changes in titration of acidity during fermentation
2.3.4 后酸化能力的结果分析 后酸化是指酸奶在4 ℃条件下低温贮藏,乳酸菌仍具有活力,继续生长繁殖,酸度会发生变化[32]。酸奶酸度增加过高会导致产品口感、品质下降。由图5 可知,在酸乳贮藏的1~10 d 期间里,6 株菌株发酵的酸乳皆呈现出酸度升高的现象,编号276、266 的菌株酸度增加明显;在贮存后期,编号276、266 的菌株滴定酸度趋于平缓,说明这两株菌株没有增强发酵乳的后酸化的能力。其余4 株的酸度迅速上升,可能是利用胞外多糖发酵产生乳酸,使得酸度继续升高。

图5 冷藏期间滴定酸度的变化Fig.5 The changes in titration of acidity during the cold storage
2.3.5 冷藏期间活菌数变化的结果分析 酸奶中含有大量活菌是体现酸奶具有益生功能的指标。由图6可知,在冷藏期间,6 株菌株发酵的酸奶的活菌数均大于107CFU/mL,符合发酵剂的标准(>106CFU/mL);其中菌株271 发酵的酸乳活菌数整体呈现下降趋势;其余5 种发酵乳的活菌数随着冷藏时间的延长,总体上呈现先上升后下降的趋势,在第10 d 到最大值,以编号276 的菌株最高为3.22×106CFU/mL,其次为编号218,为3.15×106CFU/mL。冷藏前期,乳酸菌仍在生长代谢,导致活菌数增加;在贮藏过程中,新陈代谢产生的杂物积攒以及高浓度的酸都可能抑制活菌的生长。

图6 冷藏期间活菌数的变化Fig.6 The changes in viable counts during cold storage
2.3.6 发酵酸奶的产香能力的结果分析 乙醛和双乙酰是构成酸乳典型风味的重要化合物,可以依据二者的含量判断菌株的产香能力。此外,形成香味物质的高峰一般在发酵结束后,需要12~24 h 才能完成其产香过程[33],所以本实验样品分别冷藏12、24、36 h时测定二者的含量。试验所得双乙酰标准曲线的方程为y=0.0313x+0.0009,R2=0.9991,线性良好。二者测定结果见图7 和图8。

图7 冷藏期间乙醛含量的变化Fig.7 The changes in acetaldehyde content during cold storage

图8 冷藏期间双乙酰含量的变化Fig.8 The changes in diacetyl content during cold storage
由图7 可知,六株菌株发酵的酸奶在贮藏期间乙醛含量的变化总体呈上升趋势,范畴在18~30 μg/mL 之间;其中编号276 的菌株发酵的酸奶中乙醛的含量最高,在27~29 μg/mL 之间;其次为编号231 菌株,在24~26 μg/mL 之间。编号271 的菌株发酵的酸奶在贮藏期间内乙醛含量的变化很稳定;并且产生的乙醛含量符合刘宁宁等[34]研究的产香最佳浓度:5~30 μg/mL。
由图8 可知,六株菌株贮藏期间双乙酰含量的变化范畴在1.58~3.73 μg/mL 之间;编号266、218的菌株发酵的酸奶在贮藏期间双乙酰的含量先下降后上升,可能是由于产品中乳酸与胞外多糖的综合作用所引起的;其余4 株的乙醛含量变化总体呈下降趋势,可能是因为双乙酰为散发性物质,随着贮藏时间的延长会挥发掉一部分。有研究表明,产香味物质能力强的菌株, 产双乙酰的能力较弱,因此建议选择双乙酰产量较小的酸奶发酵菌株[35]。
因此,综合比较乙醛和双乙酰含量,产香能力最强的为编号为231 的菌株。
2.3.7 发酵酸奶的质构特性 质构性质包括硬度、稠度、凝聚性、黏度指数等,对于酸奶品质的评价有着极其重要的作用[36]。由图9 可知,六株菌株发酵酸奶的测定质构结果分析,编号218 的菌株发酵酸奶的硬度为70.54 g,稠度为266.51 gsec,凝聚性为12.54 g,黏度指数为20.89 gsec,是六株菌株中质构测定结果是最好的;编号231 的菌株发酵的酸奶其次;编号271 的菌株发酵的酸奶的硬度为34.27 g,稠度为158.43 gsec,凝聚性为10.81 g,黏度指数为8.97 gsec,是测定质构结果是最低的。综合六株菌株发酵的酸奶测定质构结果从高到低排序:218>231>285>276>266>271。

图9 发酵完成后的酸奶质构Fig.9 The yogurt texture after fermentation
3 结论
菌株的生长曲线表明,6 株菌株分别在14 h(218、276、266)、16 h(271、285、231)进入对数生长稳定期,其中编号218 的菌株生长量最高。
耐受性实验结果表明,6 株乳酸菌菌株的耐酸能力、耐胆盐能力、耐渗透压能力和疏水性各不相同,其中菌株276 对酸性环境的耐受性最好(P<0.05),菌株266、218、231 的耐胆盐的能力最强,285 对高渗透压的耐受性较强,菌株285 和218 的疏水性较高,且与十六烷的结合效果较好(P<0.05)。
发酵试验结果表明,6 株菌株所发酵酸奶均在6~8 h 内凝固,冷藏期间,活菌数均保持在107CFU/mL以上,均符合发酵剂的标准,其中菌株276 的最高为3.22×106CFU/mL;菌株218 发酵制得的酸奶质构特性较好,持水力和酸化能力较其余菌株均最强,分别为60.98%和83 °T;菌株276、266 抗后酸化能力较好;菌株231 产香性能最优,乙醛含量为24~26 μg/mL,双乙酰含量为1.58~3.73 μg/mL,能够明显改善酸奶的风味。
通过综合比较6 株高产胞外多糖乳酸菌特性及发酵性能,菌株218 为一株性能良好、具有良好稳定性的菌株,可作为乳酸菌发酵剂且具有一定的应用潜力。
