猪饲粮能源结构体系研究进展
2021-11-18孙曙光姜海龙
孙曙光,严 俊,周 锴,姜 丹,姜海龙
(吉林农业大学动物科技学院,长春动物生产与产品质量安全教育部重点实验室,吉林长春 130118)
近年来,随着畜禽能量营养体系的建成与完善,该研究领域知识理念与技术创新全面升级。基于此,猪饲粮能源结构体系作为其重要分支之一便值得深入探讨。动物能量营养研究旨在摸清能量在动物机体中的供求关系,其中“供”是指饲粮能源物质的有效供能,“求”是指动物机体在生产生活中对能量的代谢需求。随着现代科技的不断进步和畜牧研究理论的不断深入,以单一能量指标为基础的现行猪饲养标准逐渐无法满足需求。因此,需要对不同饲粮能源物质的独立效应、组合效应和互作效应进一步研究归纳,建立并完善饲粮能源结构体系,从而更精准地调整能量供求关系、更高效地利用饲料资源及降低环境污染,加快畜牧业技术进步,真正促进经济增长[1]。为此,本文在回顾三大能源物质碳水化合物、脂肪、蛋白质代谢供能规律的基础上,对其独立效应和组合效应展开综述,为饲粮能源结构体系的进一步研究提供一定理论依据。
1 供能物质——碳水化合物
碳水化合物是由碳、氢、氧3 种元素组成,因其含有与水相同的氢氧比例,故称作碳水化合物。在饲料中可将其分为两大类:一是动物机体可吸收并利用的有效碳水化合物,如淀粉(葡萄糖)等;二是机体无法直接消化的无效碳水化合物,如纤维素等。其中,纤维素具有促进肠道蠕动、增强饱腹感、调节肠道菌群等生理作用。在供能方面,纤维主要是先在后肠经肠道菌群发酵并产生挥发性脂肪酸(乳酸、丙酸等),然后在肝脏中经过糖异生转化为葡萄糖参与供能,其中乙酸、丁酸可转化为乙酰辅酶A 进而氧化供能[2]。因此,葡萄糖作为碳水化合物的主要代谢产物参与机体供能。
1.1 葡萄糖的代谢供能规律 饲粮中的可消化碳水化合物在消化道各类消化酶(如淀粉酶等)的作用下以葡萄糖的形式吸收入血[3]。葡萄糖的主要代谢供能途径如图1 所示。葡萄糖是机体最常用的供能物质,在有氧条件下,一个葡萄糖分子彻底氧化成水和二氧化碳,共生成30 个三磷酸腺苷(ATP)分子。而其中有26 个ATP 分子是通过氧化磷酸化作用生成的(糖酵解产生的NADH 通过甘油磷酸穿梭途径转运),若NADH 通过苹果酸-天冬氨酸穿梭途径转运,则多生成2 个ATP 分子,总计32 个[4]。在无氧条件下,一分子葡萄糖生成乳酸仅产生2 个ATP分子。除了生成ATP 供能,少量葡萄糖经戊糖磷酸途径产生的NADPH 可以作为氢负离子供体参与机体还原性生物合成(如脂肪酸及胆固醇的生物合成),产生的磷酸核糖可以为细胞提供不同结构糖分子(如在核酸的生物合成中提供核糖)。葡萄糖在机体细胞中的储存形式为糖原,糖原是动物机体最易动员的能量贮存物质[5]。

图1 葡萄糖代谢供能途径[6]
1.2 葡萄糖的独立效应 葡萄糖是神经细胞的重要能量来源,更是大脑神经系统的主要供能物质[7]。Schönfeld等[8]研究表明,脑细胞很少利用脂肪酸氧化供能,主要原因为脂肪酸的β氧化消耗了更多氧气,增大了神经元缺氧风险;并且脂肪酸氧化生成ATP 的速率较慢,无法满足神经元连续快速的放电消耗。当然,在动物严重饥饿或血糖水平过低的情况下,脑细胞也可利用脂肪酸代谢生成的酮体作为临时供能物质。与神经元的能量快速消耗原理相同,肌肉的能量供应也主要依赖于葡萄糖。肌细胞利用肌糖原分解产生的葡萄糖作为剧烈运动中的主要能量来源。肝脏中的肝糖原可用于维持机体血糖稳定,并对机体某些有毒化学物质起到解毒作用,从而增强机体免疫力[9]。对于繁殖母猪而言,葡萄糖既是胎儿早期发育的主要供能物质[10],又是乳腺乳糖代谢的唯一前体物质。Sato 等[11]研究表明,葡萄糖在猪卵母细胞核成熟过程中起到重要作用。
2 储能物质——脂类
脂类包括油脂和类脂。油脂又称甘油三酯或三酰甘油,一般将在常温下呈液态的油脂称为油,呈固态的称为脂肪。类脂则包括磷脂、糖脂和胆固醇及其脂。类脂是生物膜的重要组成部分,它对脂溶性维生素(A、D、E、K)的吸收以及钙、磷代谢等均起着重要作用。在供能方面,油脂在各能源物质中能值最高、热增耗最小、净能产生率最高[12]。因此,油脂作为机体最重要的储能物质,拥有不可替代的作用。
2.1 油脂的代谢供能规律 饲粮中的可消化油脂在胆汁的作用下乳化,并在消化道内各类脂肪酶的作用下分解后吸收入血[13]。油脂的主要代谢供能途径如图2 所示。1 个三酰甘油分子由3 个脂肪酸分子和1 个甘油分子组成,其中甘油仅提供5% 的能量,剩余95% 的能量贮存在脂肪酸链中。以1 分子软脂酸(十六碳)彻底氧化为例,总计可产生108 个ATP 分子。已有研究表明,脂肪的有效能约为碳水化合物的2.25 倍[14]。由此可见,脂肪的有效能大大高于碳水化合物,且当动物处于严重饥饿或患有糖尿病的情况下,体内的草酰乙酸主要用于糖异生过程,导致脂肪酸氧化生成的乙酰辅酶A 无法进入柠檬酸循环,仅可产生酮体,严重时会导致“酸中毒”(由于产生大量乙酰乙酸和D-β-羟丁酸)和“酮血症”(由于产生大量丙酮)。

图2 脂肪代谢供能途径[6]
2.2 油脂的独立效应 脂肪酸是心脏、肾脏等的主要能量来源[8],其原因可能是脂肪酸链中贮存能量较高,可以为较缓慢的持续能量消耗提供保障。同样由于脂肪酸的氧化过程缓慢且持续,所以油脂的热增耗较低。Espinosa 等[15]研究表明,在饲粮中添加适量油脂,可有效提高断奶仔猪的抗热应激能力,并提高饲料报酬。饲粮中添加适当油脂可提高育肥猪生长速度和饲料利用率,有效缩短育肥期,而育肥阶段油脂的适宜添加量为2%~5%,过量则会导致胴体品质下降[16]。除此之外,不同来源的油脂会产生不同的肥育效果,这可能与它们不同的能量值相关。Liu 等[17]研究结果证明,含有较多饱和脂肪酸的油脂为育肥猪提供的能量可能比含有较多不饱和脂肪酸的油脂更多。在基因表达方面,不同来源油脂会对育肥猪脂肪组织和肝脏脂质代谢相关基因的转录产生影响[18]。因此,饲粮能源结构体系不单单是指三大能源物质之间的平衡,也是指各能源物质内部不同来源组成之间的平衡。Jin 等[19]研究表明,在繁殖母猪妊娠后期和哺乳期饲粮中添加油脂,可提高初乳和常乳中的乳脂含量,进而提高哺乳仔猪的生长性能和断奶成活率。并且在母猪饲粮中添加油脂可以改变仔猪脂肪酸组成和肝基因表达[20]。在猪生产中,饲料中添加油脂还可以减少粉尘的产生,改善畜舍环境[8]。
3 结构物质——蛋白质
蛋白质是一种复杂的有机化合物,它是由氨基酸以脱水缩合方式组成的多肽链经过盘曲折叠形成的具有一定空间结构的物质[21]。蛋白质主要作为一种结构物质为动物机体的生长发育和组织器官的修复提供建筑材料。除此之外,一部分功能性氨基酸还起着调节维持、生长、繁殖和免疫等关键代谢途径的作用[22]。
3.1 蛋白质的代谢供能规律 饲粮中的可消化蛋白质在胃酸与胃蛋白酶的共同作用下变性并分解,在肠道经胰蛋白酶等的作用下进一步分解为氨基酸与肽并吸收入血[23]。在机体正常生理条件下,摄入的氨基酸中仅有小部分以碳链形式进行氧化供能或转化为葡萄糖,其余大部分将进入氨基酸再循环并参与机体的生长发育。蛋白质的主要代谢途径如图3 所示。氨基酸在体内的再循环频繁且复杂,且需脱去氨基生成尿素排出体外,这些过程中大量能量以热的形式散失,这可能是蛋白质相对于碳水化合物和油脂热增耗较高的原因。机体摄入的多余氨基酸无法被存储,只能通过分解代谢排出体外。因此,氨基酸的合理供给尤为重要。
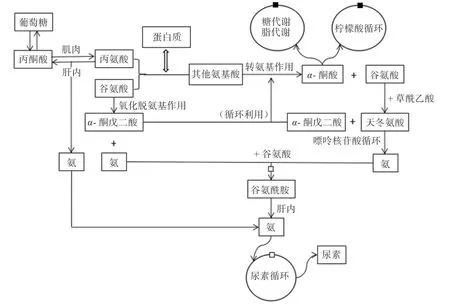
图3 蛋白质代谢供能途径[6]
3.2 蛋白质的独立效应 氨基酸是动物机体蛋白质合成的底物,在猪生长发育(骨、肉、皮、脂)和器官的组成与修复中均起到不可替代的作用,其中赖氨酸是猪饲粮中的第一限制性氨基酸[24]。在调控猪肉品质方面,氨基酸可作为信号分子发挥作用[25]。对繁殖母猪而言,氨基酸对母猪妊娠期内胎儿、胎盘和子宫的生长发育尤为重要[26]。其中精氨酸成为妊娠母猪的条件性必需氨基酸和限制性氨基酸,原因是精氨酸作为含氮活性物质(一氧化氮、多胺、肌酸等)的合成前体,可通过提高机体抗氧化能力、调节孕体血管密度与直径等改善孕体妊娠内环境[27]。传统观念一般将蛋白质独立于能量体系之外,主要原因是蛋白质供能较少,但其实蛋白质在供能方面的独立效应也是不可忽视的。Der Schoor 等[28]研究表明,猪肠道上皮细胞会直接利用蛋白质分解代谢产生的氨基酸氧化供能,其中谷氨酸和天冬氨酸是肠道上皮细胞重要的能量来源。动物机体在血糖过低的情况下,由于无法将油脂大量转化为葡萄糖,便会利用氨基酸转化为葡萄糖或直接氧化供能,这可能也是肠道上皮细胞(尤其是后肠)利用氨基酸供能的原因。
4 饲粮能源结构体系
4.1 物质与能量的联系 动物机体的物质代谢和能量代谢其实是一个过程的2 种表现[29],营养物质被动物机体吸收,在体内互相转化,从而达到机体此刻所需的物质形态,转化过程伴随着能量的吸收、释放、传递和损失。能量依附于能源物质而存在,伴随着能源物质的转化而流动。能量与能源物质之间是统一的,更是不可分割的。传统观念一般仅利用单一的能量指标(代谢能、净能等)或将能量看成单独的营养素,究其原因可能是认为能源物质之间可相互转化,只要能量供应充足,动物机体的需求即可得到满足。但实际上能源物质之间的转化是互相制衡的,在转化过程中更是伴随着能量的消耗和浪费。现行猪营养标准(NRC)便主要依托能量指标,以消化能、代谢能为基础,结合蛋白质和氨基酸共同形成一个较为完善的体系,为生猪产业的整体稳定发展提供了保障。但由上文可知,不同能源物质有不同的供能途径和独立效应,若将其笼统的折合为能量值来计算,则具有一定的局限性。因此,优化猪饲粮能源结构,建立多维度能量需求体系变得尤为重要[1]。赵胜军等[30]通过建立猪可代谢葡萄糖模型,期望构建猪可代谢营养评定体系,结合“能量载体物质”将物质与能量在一定程度上关联起来。这与本文所述问题异曲同工。可见,要想在现行单一能量指标的基础上寻求突破,需要重点关注各能源物质代谢供能途径,并理清其互作、组合效应,将物质与能量有机结合,并进行整体优化思考。
4.2 三大能源物质的组合效应 三大能源物质的部分代谢供能模型(猪生长模型)如图4 所示,该模型便可较为完整的体现碳水化合物、脂肪和蛋白质之间的转化与利用[31]。该模型以三大能源物质为起始,将它们在猪体内的代谢产物以乙酰辅酶A 作为枢纽联系起来,代谢物在体内可以通过不同途径互相转化,转化过程伴随着能量的吸收与释放。这其中代谢物除转化为乙酰辅酶A 分解供能外,它们之间的互相转化过程大多需要吸收、消耗能量,这个过程会使转化效率降低。而由于各能源物质具有不同的独立效应,在饲粮能源结构不均衡的情况下其互相转化又是不可避免的。由该模型也可以看出,三大能源物质之间互相转化、相互影响,整体达到一种平衡的状态,从而共同为猪的生长提供能量。改变其中一种或几种能源物质供应或组成,该体系会整体改变,从而达到不同的平衡效果,进而直接影响猪的生长生理状态。因此,优化饲粮能源物质组合配比对于提高能源物质利用效率、改善猪的生长生理状态至关重要。

图4 猪生长代谢模型[31]
国内外不少研究已经通过改变猪饲粮能源结构发现并解决了一些问题,但传统研究更多是点对点的探究,并没有从饲粮能源结构的组合、互作效应上入手,形成系统化的体系。Benz 等[32]研究表明,不同油脂水平的饲粮会改变育肥猪生长性能、胴体性状和胴体脂肪品质。Schenck 等[33]发现,添加油脂会显著降低仔猪的日增重。Lawrence 等[34]设计了不同油脂水平饲粮,并通过改变玉米和豆粕含量保持饲粮等能等氮,其研究发现随着油脂的增加会显著降低断奶仔猪采食量,断奶仔猪(约27 日龄)前2 周日增重也会随饲粮脂肪含量的增加线性下降,能量利用率也呈下降趋势。以上研究虽然实质是改变了猪饲粮能源结构,但仅是对添加的物质进行了点对点的研究,并没有从动物机体整体的能量代谢角度思考其背后的逻辑与机制。因此,秦贵信[1]提出了饲粮能源结构体系可作为畜禽能量营养体系的重要发展方向,需系统研究不同饲料能量营养源在畜禽体内能量代谢的独立效应、累加效应以及互作效应,从而得到饲粮能源结构的优化模式和结构性需求。
在此基础上,刘飞飞[35]在保持仔猪饲粮等能等氮的基础上,调整饲粮碳水化合物/ 油脂比分别为11.7:1.0(LF 组)、1.5:1.0(MF 组)和1.0:6.9(HF 组),结果表明,改变能源物质的组成显著影响了仔猪的生长性能。其中LF 组仔猪的生长性能显著优于MF、HF 组;与LF 组相比,MF 和HF 组氮沉积显著下降。究其原因可能是在不同能源结构下,机体不同供能平衡对猪生长生理状态产生了影响。在试验饲粮等能等氮的条件下,添加过高的油脂无法满足机体对碳水化合物的需求,就会有大量的蛋白质转化为糖类或直接参与到供能途径中(如图3),导致蛋白质沉积率降低。
Han 等[36]通过对生长猪进行呼吸测热、碳氮平衡等试验,证明饲粮能源结构的调整改变了营养物质氧化供能比例;不仅如此,环境温度的改变与饲粮能源结构之间有一定互作效应;随着环境温度的降低,脂肪氧化供能量显著增加,碳水化合物氧化供能量显著降低,氮沉积率有降低趋势,造成此结果的原因可能是环境温度的改变影响了机体对能源物质的需求,从而改变了能源物质氧化供能比例。
5 小 结
上述研究充分证实了饲粮能源结构在猪生产生活中的重要价值,为继续构建猪饲粮能源结构体系奠定了基础。但饲粮能源结构体系的研究目前还处于起步阶段,只有一些点状的试验作为其理论支撑,未能形成一个完整体系。今后,为了完善猪饲粮能源结构体系,还需进行不同生长阶段、不同环境因素、不同遗传基础等方向的饲粮能源结构研究,根据猪机体的不同条件,进行多维度、多角度的饲粮营养配比,实现能量营养供求的更精准对接,为我国猪能量营养的系统化研究提供一条切实可行的道路。
