水稻分蘖期叶片淀粉和蔗糖代谢途径的转录组动态分析
2021-11-16刘天奇张雪晴张时煜高红秀金正勋张忠臣姜振峰
谢 威,刘天奇,张雪晴,张时煜,高红秀,金正勋,张忠臣,姜振峰
(东北农业大学 农学院,哈尔滨 150030)
水稻是最重要的粮食作物之一,是全国60%以上人口的主要食物来源[1]。水稻分蘖期是由秧田向本田过渡的生长关键期,这一时期的生长对水稻的总产量有重要影响[2-6]。张洪程等[7-9]研究表明,水稻从拔节一直到成熟期的干物质积累量对产量产生正向影响。另外,水稻分蘖期的栽培调控会导致稻米中直链淀粉含量下降,从而提高稻米品质[10]。
淀粉是稻米中最重要的能量储存物质[1],其合成涉及一系列酶,包括颗粒结合淀粉合酶(GBSS)、可溶性淀粉合酶(SSS)、分支酶(SBE)、脱分支酶(DBE)、异淀粉酶(ISA)等,对应基因有20多个[1,11]。淀粉合酶已鉴定出5个亚家族,包括颗粒结合淀粉合酶(GBSS)、淀粉合酶Ⅰ(SSⅠ)、淀粉合酶Ⅱ(SSⅡ)、淀粉合酶Ⅲ(SSⅢ)和淀粉合酶Ⅳ(SSⅣ)。直链淀粉的合成必须经过GBSS的催化,Dian等[12]研究水稻淀粉合成时发现基因GBSSⅡ。水稻储藏器官中的直链淀粉含量主要由GBSSⅠ控制[13],水稻营养器官中的瞬时淀粉含量由GBSSⅡ调控[14]。SSⅠ与支链淀粉的极短链合成有关,SSⅢ与长B1和B2链合成有关[11]。Commuri等[15]在研究玉米时发现,SSⅠ对支链淀粉亲和力强,直链淀粉次之,对糖原的亲和力最低。SSⅡa参与水稻A + B1链的合成[11],为一种主效基因,调控稻米的糊化温度[16],Umemoto等[17]将SSⅡa定位在水稻的第6号染色体短臂上的ALK位点。有研究指出,GBSSⅠ除有催化直链淀粉合成的功能外,对支链淀粉的合成也有一定影响[1]。ISA1、ISA2和ISA3是水稻异淀粉酶(ISA)的3种同工型。ISA1在水稻的根茎叶中未发现表达,在水稻早期胚乳发育中进行表达[18],而ISA2在水稻叶片与胚乳中进行表达,ISA3则主要在水稻叶片中进行表达[19]。
有研究发现与水稻产量相关的因素对水稻的糖代谢有影响[20]。有报道称,参与蔗糖代谢的基因有OsINV3和OsINV2等[21],在水稻中已鉴定出5个蔗糖共运体基因家族成员,分别为OsSUT1、OsSUT2、OsSUT3、OsSUT4和OsSUT5[22]。
尽管与淀粉合成相关基因的功能有诸多报道,但在水稻分蘖期叶片中行使功能的淀粉合成相关基因的报道还很少,且水稻淀粉合成相关基因的转录调控关系尚不清楚。本研究对分蘖期‘五优稻4号’(‘WYD4’)的叶片进行转录组分析,为揭示淀粉合成相关基因的转录调控和水稻分蘖期的栽培调控提供分子水平的依据。
1 材料与方法
1.1 水稻材料准备
试验在哈尔滨东北农业大学盆栽场进行。首先将水稻品种‘五优稻4号’(‘WYD4’)种子用30%次氯酸钠灭菌30 min后放入锥形瓶中,然后用0.6%的硝酸在室温下处理以打破休眠状态,移入30 ℃培养箱催芽,在温室中播种。三叶一心期移栽至0.24 m2的白盆中,插秧规格为每穴3株苗,插秧方式为10 cm×10 cm。
1.2 栽培管理
盆栽土壤的pH为7.52,基础肥力为全氮 1.51 g/kg、速效氮158.76 mg/kg、速效磷50.65 mg/kg、速效钾177.68 mg/kg、有机质3.11%。基肥为纯氮72 kg/hm2、纯磷90 kg/hm2和纯钾72 kg/hm2,移栽后7 d施加分蘖肥,分蘖肥为纯氮54 kg/hm2,水分管理为常规管理。在分蘖期氮素处理后第7天、第14天和第21天设置3次生物重复,以确保试验的可靠性。
1.3 样品制备
在分蘖期氮素处理后第7天、第14天和第21天拍照,分别收集每株苗顶部第一片完全展开的叶片并保存在已标记Nt7d,Nt14d,Nt21d的试管中,置于液氮中冻存0.5 h后在干冰保存下快递至浙江安诺优达生物科技有限公司,进行RNA提取和转录组分析。利用Qubit 3.0荧光仪(Life technologies,CA,USA)进行 RNA纯度和浓度的测定,并用Agilent 2100 RNA Nano 6000检测试剂盒(Agilent technologies,CA,USA)对 RNA样品的完整性和浓度进行分析。聚类分析(Clustering)与测序使用 HiSeq PE Cluster Kit v4-cBot-HS(Illumia)试剂在 cBot 上完成。随后利用 HiSeq 测序平台获得150 bp的双端测序Reads数据。最后通过GffCompare将RNA-seq数据的转录组与参考转录组进行比对,结合HTSeqv 0.6.0 Reads计数(http://www-huber.embl.de/users/anders/HTSeq/doc/overview)计算FPKM(每千碱基每米的片段映射读数),从而估计每个样本中基因的表达水平。
1.4 差异基因表达分析
利用DESeq2方法检测在基因表达水平基础上的差异表达基因,得到P值,然后根据Benjamini 和 Hochberg 方法校正控制假阳性。差异基因筛选主要参考差异倍数值(FoldChange)以及q值(padj值,矫正之后的P值)作为相关指标,通常选取|lb FoldChange|≥1和q< 0.05的差异基因作为显著差异基因。基因功能描述参考The Rice Annotation Project Database(rapdb.dna.affrc.go.jp)。
1.5 功能富集分析
首先,通过 GO(Gene ontology,http://geneontology.org/)功能显著性富集分析确定差异表达基因行使的主要生物学功能。根据计算注释到每个GO条目中的差异表达基因数目,应用超几何检验,找出与整个基因组背景相比,差异表达基因显著富集的GO 条目,其筛选标准为q≤ 0.05。其次,利用KEGG(Kyoto encyclopedia of genes and genomes,http://www.kegg.jp/)找出差异表达基因中显著性富集的Pathway。对KEGG中每个Pathway应用超几何检验进行富集分析,找出差异表达基因中显著性富集的Pathway,并对每个比较组进行q值的KEGG条目分析和作图。
1.6 蛋白互作网络
利用 STRING 蛋白质互作数据库(http://string-db.org/)将目标基因集直接映射到水稻的蛋白互作网络。然后将蛋白互作网络数据文件导入 Cytoscape 软件,并根据目标基因集中的基因属性进行互作网络的可视化编辑。
1.7 数据处理与分析
表和图的数据分析与处理利用WPS Office(11.1.0.9339)及Adobe Photoshop(13.0 20120315)完成。
2 结果与分析
2.1 分蘖期WYD4植株表型
从第7天、第14天和第21天WYD4的分蘖期表型可以看出,WYD4的群体长势越来越旺盛,冠层覆盖度越来越高(图1)。这个结果为采集分蘖期WYD4叶片进行转录组分析提供了表型基础。
2.2 转录组测序数据统计及质量评估
通过对第7天、第14天和第21天的9个分蘖期WYD4叶片样本进行转录组测序,将Raw data经数据过滤后分别产生45 454 356、42 789 826、47 737 496、41 771 146、42 587 134、40 932 800、44 273 324、43 704 008和45 662 926个Clean reads,且Q30为92.67%~93.31%。将过滤后的测序序列进行基因组定位分析,9个样本分别有 97.02%、96.94%、96.76%、96.83%、97.01%、96.90%、96.46%、96.61%和96.68%的Clean reads能够比对到水稻基因组上(表1)。这些结果表明转录组测序数据产量及质量都比较高,可用于进一步分析。

表1 测序数据统计与质量检测
2.3 分蘖期水稻叶片差异基因的表达
WYD4叶片在分蘖期共产生12 815个差异表达基因(DEGs,Differential expression genes),通过第14天与第7天的对比,有3 962个上调差异表达基因,有3 246下调差异表达基因;通过第21天与第14天的对比,有3 374个上调差异表达基因,有5 198个下调差异表达基因;通过第21天与第7天的对比,有3 062个上调差异表达基因,有3 953个下调差异表达基因。在第21天时达到差异表达基因数目的一个高峰,且下调差异表达基因数目远远超过上调差异表达基因数目(图2-A),这说明WYD4分蘖期的生长调控机制是动态变化的,且主要作用在后期。
为了展示不同时间下WYD4分蘖期DEGs表达模式的异同,进行层次聚类分析并绘制Venn图和Volcano图,层次聚类分析展示分蘖期WYD4的DEGs在不同的时间点的变化模式(图2-B~2-D),韦恩图分析显示,有7 152基因在两个时期差异表达,有1 414个基因在WYD4整个分蘖期都发生差异表达(图2-H),从火山图可以清晰地看出WYD4在不同时间点差异基因的表达倍数和显著性变化情况(图2-E~2-G)。这些结果展示分蘖期WYD4叶片差异基因的变化情况,为揭示水稻分蘖期的生长调控机制提供了转录组水平的参考。
2.4 GO分析
为了分析分蘖期差异表达基因的功能,对WYD4分蘖期叶片进行GO注释和GO富集分析。结合(Binding)和催化活性(Catalytic activity)是DEGs在分子功能(Molecular function)方面注释最多的条目,细胞组分(Cellular component)是DEGs在细胞组分(Cell part)方面注释最多的条目,代谢过程(Metabolic process)和细胞过程(Cellular process)是DEGs在生物过程(Biological process)方面注释最多的条目(图3)。通过对WYD4分蘖期叶片3个时间点的DEGs进行GO分析发现,水稻分蘖中期(7~14 d)与前期(0~7 d)的DEGs主要富集在生物调节(Biological regulation)、胞质部分(Cytoplasmic part)和蛋白丝氨酸/苏氨酸激酶活性(Protein serine/threonine kinase activity),水稻分蘖后期(14~21 d)与中期的DEGs主要富集在激酶活性(Kinase activity)、细胞组分(Cell part)和生物调节(Biological regulation),水稻分蘖后期与前期的DEGs主要富集在氧化还原酶(Oxidoreductase activity)、细胞组分(Cell part)和刺激反应(Response to stimulus)(图4)。
这些结果表明细胞组分(Cellular component)是分蘖期 WYD4叶片中DEGs主要集中的部分,这可能是因为分蘖期属于水稻营养生长最旺盛的阶段,是株型构建的关键时期,参与该时期细胞形成的差异表达基因较多。
2.5 KEGG富集分析
通过KEGG富集分析WYD4分蘖期叶片,发现不同时间点的DEGs主要集中在淀粉与蔗糖代谢(Starch and sucrose metabolism)、氨基糖与核苷酸糖代谢(Amino sugar and nucleotide sugar metabolism)、碳代谢(Carbon metabolism)和核糖体(Ribosome)等途径(图5)。其中淀粉与蔗糖代谢途径在WYD4整个分蘖期都达到显著富集水平,这说明淀粉与蔗糖代谢途径在水稻的分蘖期尤为重要。
2.6 分蘖期WYD4淀粉与蔗糖的代谢途径关键差异表达基因
通过对淀粉与蔗糖的代谢途径的差异表达基因在分蘖期WYD4水稻叶片中的表达情况进行分析发现,有6个基因在3个时间点都差异表达,分别是PGI2(LOC_Os06g14510)、OsAGPS1(LOC_Os09g12660)、GBSS1(LOC_Os06g04200)、glgP(LOC_Os03g55090)、ISA2(LOC_Os05g32710)和ISA3(LOC_Os09g29404)(图6)。
这些基因随着时间推移,都呈先升高后降低的表达模式,表明这些关键基因在水稻的分蘖中期起到重要的调控作用。
2.7 淀粉与蔗糖途径相关的转录因子
为了详细了解淀粉与蔗糖途径在WYD4分蘖期转录表达,分析了WYD4分蘖期叶片差异表达超过2倍的转录因子。结果发现,差异表达倍数超过2倍的转录因子共有24个,且全部为上调表达,分别来自ERF、NAC、HD-ZIP、FAR1、BES1、HB-other、C2H2、WRKY和MIKC等9个家族,其中ERF和NAC家族的转录因子在蔗糖与淀粉途径中表达基因数目最多,都为6个。FAR1家族的SSⅠ(LOC_Os06g06560)差异表达倍数最大,在第14天时差异表达达到9.7倍(表2)。进一步分析差异表达超过2倍的转录因子在水稻分蘖期随时间推移的变化情况,发现GN1_2_3(LOC_Os07g32600)、GN4(LOC_Os07g38930)、GN4(LOC_Os07g43940)、GN5_6(LOC_Os08g23720)、GN5_6(LOC_Os09g09980)、AMY2A(LOC_Os06g49970)、BGLU4(LOC_Os01g67220)、BGLU6(LOC_Os03g11420)、BGLU19(LOC_Os05g30250)、BGLU28(LOC_Os08g39870)、BGLU31(LOC_Os09g33680)BGLU1(LOC_Os01g32364)、malZ(LOC_Os06g46284)、SSⅠ(LOC_Os06g06560)、E3.2.1.2(LOC_Os03g04770)、RSUS1(LOC_Os03g28330)、BGLU16(LOC_Os04g43390)、GH9A3(LOC_Os03g52630)主要在水稻的分蘖前期起作用,E2.4.1.14(LOC_Os10g40324)、XK5(LOC_Os05g44760)、E2.4.1.14(LOC_Os06g42824)、E3.2.1.2(LOC_Os10g41550)主要在水稻分蘖中后期起作用,SSⅡC(LOC_Os10g30156)、scrK(LOC_Os01g66940)在水稻分蘖全期起作用。这说明在淀粉与蔗糖的代谢途径中,大部分的转录因子都在水稻分蘖的前期起作用,这可能为水稻的栽培调控提供新思路。
2.8 淀粉与蔗糖代谢途径关键转录因子SSⅠ(LOC_Os06g06560)的互作蛋白预测
鉴于转录因子分析结果,淀粉与蔗糖代谢途径的转录因子多在分蘖前期起作用,因此对前期的转录因子进行互作蛋白的预测,发现ERF家族共有203个互作蛋白;NAC家族共有691个互作蛋白;FAR1家族共有103个互作蛋白;BES1家族共有36个互作蛋白;HB-other家族共有401个互作蛋白;C2H2共有98个互作蛋白;WRKY家族共有145个互作蛋白(表3),但是基于FAR1家族的SSⅠ(LOC_Os06g06560)在水稻分蘖前期差异表达倍数为9.7倍,所以对SSⅠ(LOC_Os06g06560)进行互作蛋白分析,结果发现64个蛋白与SSⅠ(LOC_Os06g06560)互作,这些蛋白大多数与淀粉与蔗糖代谢途径相关,且都在前期差异表达,同时部分蛋白还在本试验的整个分蘖期差异表达。这些结果说明SSⅠ(LOC_Os06g06560)在水稻分蘖期对淀粉与蔗糖代谢途径起到极为重要的调控作用(表4)。
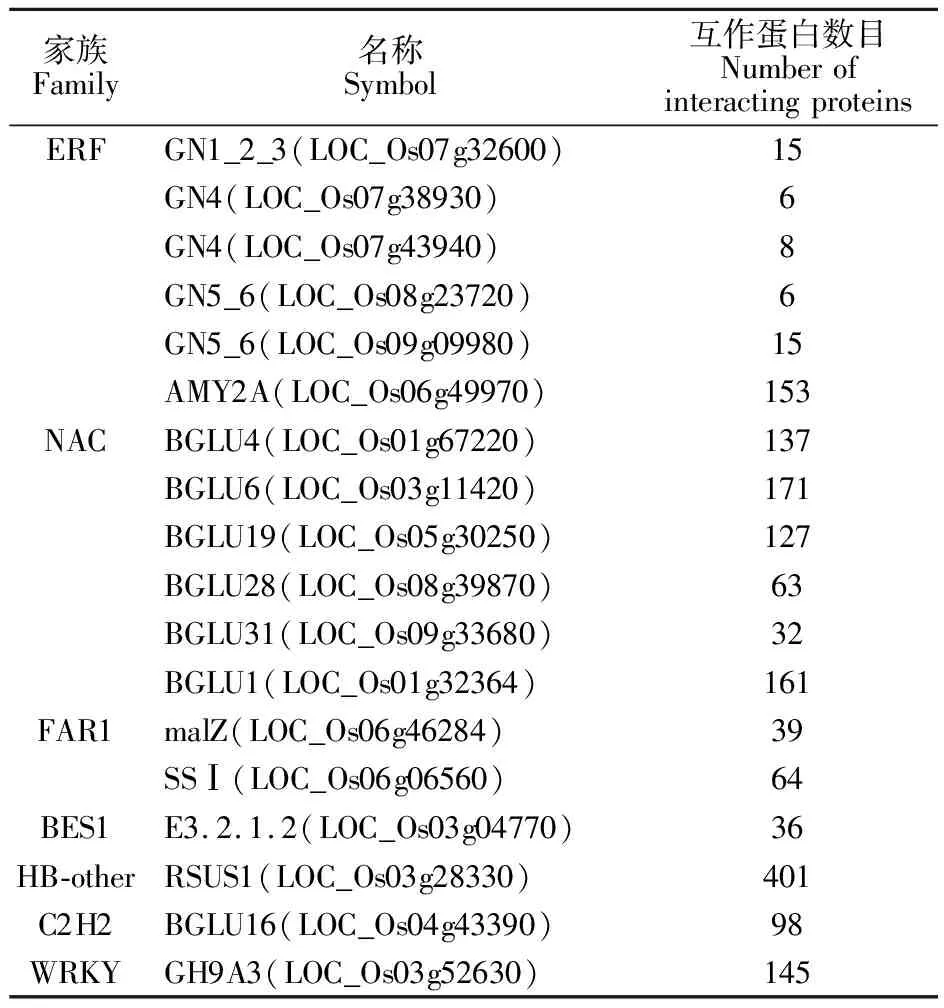
表3 转录因子的互作蛋白

表4 SSⅠ(LOC_Os06g06560)的互作蛋白
3 讨 论
直链淀粉和支链淀粉含量构成水稻淀粉含量,淀粉含量明显影响稻米RVA谱特征[23],GBSSⅠ是直链淀粉合成的关键基因,抑制GBSSⅠ的表达会导致直链淀粉含量的降低[24];ISA1能够决定水稻籽粒中的淀粉结构[25-26];ISA3是控制淀粉颗粒降解的基因[27]。已有研究报道GBSSⅠ、ISA1和ISA3在灌浆期的表达呈先升高后降低的单峰曲线变化情况[19,28-32],本研究发现,GBSSⅠ、ISA1和ISA3在分蘖期的水稻叶片也呈先升高后降低的单峰曲线变化趋势;另外,本研究中PGI2(LOC_Os06g14510)、OsAGPS1(LOC_Os09g12660)和glgP(LOC_Os03g55090)在水稻叶片的整个分蘖期表达也达到显著差异水平,并呈现单峰曲线变化趋势,但这几个基因在分蘖期淀粉蔗糖途径中的分子功能还有待于进一步 完善。
在支链淀粉合成过程中,SSⅠ行使着重要的功能。SSⅠ是可溶性淀粉合酶(soluble starch synthase,SS)4个亚家族中唯一没有同工型的酶[33],Yasunori等[34]发现SSⅠ和SSⅢ是水稻胚乳中主要的SS酶,且SSⅠ活性明显高于SSⅢ活性,SSⅠ活性高达总SS活性的70%左右。SSⅠ位于水稻的6号染色体上,由15个外显子和14个内含子构成[35]。吴洪恺等[36]用糯稻为背景构建近等基因系,发现SSⅠ对稻米的RVA谱有显著影响。康翠芳等[37]通过裂区试验发现SSⅠ基因和SSⅢ-1基因间存在互作效应,同时发现这两个基因间的互作效应对GT产生极显著影响,而对其他RVA谱特征值也产生显著影响。杨博文等[38]通过回交重组自交系发现SSⅢ-1和SSⅠ、SSⅠ和PUL的基因互作效应在GT有极显著的影响外,对于其他所有的RVA谱特征值都产生显著影响,同时SSⅠ、SSⅢ-1和PUL间的3基因互作效应也对GT有极显著影响。Nakamura等[39]在体外进行酶促反应时发现SS酶中的SSⅠ与BEs酶存在相互作用。Crofts等[40]进行淀粉合成相关酶分析时发现水稻胚乳中的蛋白复合物存在SSⅠ、SSⅡa、BEⅡb、ISA、PUL和Phol。Chen等[41]发现SSⅠ和PUL,SSⅠ和BEs间存在蛋白互作,且因水稻品种不同蛋白互作模式也不相同。陈雅玲等[1]通过STRING 网站预测SSs家族可能与BEs家族和DBEs家族存在互作关系,但没有试验进行证实。邱献锟等[42]发现SS酶在水稻灌浆期活性表现的越早越高,越容易促进蔗糖与淀粉的转化,从而使淀粉含量升高。本研究对分蘖期WYD4的叶片进行转录组分析,发现淀粉与蔗糖途径在分蘖期就已经呈现显著的差异,而SSⅠ作为FAR1家族的转录因子在分蘖前期差异表达倍数极大,达到9.7倍。并预测与64个蛋白基因存在互作关系,其中包括BEs家族中的BEⅠ(LOC_Os06g51084)、ISA家族的ISA1(LOC_Os08g40930),这与前人结果相同,而SSⅢ-1(LOC_Os04g53310)、SSⅡa(LOC_Os06g12450)和BEⅡb(LOC_Os02g32660)并没有在本研究的结果中出现。综合本研究和前人的研究结果[1,37-44],绘出淀粉合成途径关键酶互作网络图(图7),这些结果对于解析参与淀粉合成的基因间的表达调控存在重要指导意义,对通过基因网络调控改良稻米品质和定向水稻育种具有重要意义,并对用通过水稻精准栽培调控的方法提高水稻产量的同时改良稻米品质指明方向。
4 结 论
水稻分蘖期淀粉与蔗糖途径的6个基因PGI2、OsAGPS1、GBSS1、glgP、ISA2、ISA3以及转录因子SSⅠ在分蘖期不同时间点的表达是有显著差异的,这一点可以为水稻分蘖形成和群体构建提供时间上的能量来源,也为水稻栽培过程中精准调控提供转录组水平的参考依据。
