基于高通量测序的坡豪湖湿地土壤细菌群落结构分析
2021-11-15罗奉奉李雪凤刘鑫泰高丽霞覃勇荣
罗奉奉,付 跃,李雪凤,刘鑫泰,高丽霞,覃勇荣,3*
(1.河池学院 化学与生物工程学院,广西 河池 546300;2.微生物及植物资源开发利用广西高校重点实验室,广西 河池 546300;3.桂西北经济植物生物技术研究中心,广西 河池 546300;4.华南农业大学 生命科学学院,广东 广州 510000)
湿地是介于陆地生态系统和水生生态系统之间的过渡生态系统,兼具陆地和水体的生态特性,其生物多样性非常丰富;土壤微生物作为湿地生态系统的重要生命体,对湿地物质循环、水土保持、生物多样性保护、生态净化等[1-4]起非常重要的作用;湿地的环境条件变化也会引起微生物群落多样性和生态功能的改变[5-7],因此,土壤微生物作为湿地土壤亚系统变化的预警和敏感指标[8],对评估湿地生态系统资源和功能具有重要意义。
近年来随着人为活动和环境变化的影响,众多学者开始关注湿地生态系统的研究,主要从水文、土壤、气候、污染物、植被等方面分析与微生物的相互关系,探索导致湿地生态稳定性变化的主要因素,这些研究认为在季节变化[9]、含水量[10,11]、土壤元素循环[12,13]、植被演替[14]、耕作方式[15,16]、重金属污染[17]等因素的影响下,湿地土壤微生物群落多样性呈现一定的特征性变化。
广西东兰坡豪湖国家湿地公园处在桂西北喀斯特石山地区的河池市东兰县境内,1992年在红水河中游的岩滩水电站蓄水封闸后,在东兰县长乐镇坡豪境内自然形成了湖泊湿地[18]。与其他湿地不同的是,坡豪湖湿地每年受岩滩水电站蓄放水的影响,湖泊周边土壤常处于不同程度的湖水浸润状态。在2014年12月,国家林业局正式批准广西东兰坡豪湖国家湿地公园开展试点建设[19],旨在更好地保护坡豪湖湿地的生物多样性。本研究根据喀斯特石山与红水河接壤形成的坡豪湖湿地的生境特点,结合Illumina高通量测序技术从微生物生态学角度探索了坡豪湖湿地土壤细菌群落的结构变化及其与土壤环境因子、下游水库蓄放水的关系,分析了细菌菌群在该湿地生态系统中的变化特征,旨在为坡豪湖湿地生态系统资源的保护管理和可持续利用提供理论依据。
1 材料与方法
1.1 研究区域概况
研究区域为广西河池市东兰坡豪湖国家湿地公园(24°25′02″~24°27′14″N,107°23′53″~107°32′40″E),属亚热带季风气候,夏长冬短,雨热同季,气候温和,光照充足,海拔217 m,年平均气温为19~21 ℃,年平均降雨量1563 mm;该公园处桂黔滇喀斯特石漠化防治生态功能区。
1.2 样品采集
岩滩水电站蓄放水情况:每年3~9月为放水期,9月~次年3月为蓄水期。选取坡豪湖水陆交替带(从最高水位向湖中靠近2 m位置)为试验地,在坡豪湖周边土壤区域按距离2~3 km选取样地,共选8个采样区,每个样地设置3个样方做平行,样方间隔20 m,样方大小50 cm×50 cm,按五点取样法随机取样,混合为1个土样,去除土壤中的动植物残体和碎石等杂物,用直径5 cm的土钻采集距土壤表层5~20 cm的土壤,每个样地土壤用无菌自封袋分装成两份,置于冰盒当天运回实验室。一份置于-80 ℃保存,用于土壤细菌群落分析;另一份自然风干,过100目筛,用于土壤理化性质的测定。根据广西的气候特征,于2017年4月开始采集土壤样品,为春季样;随后在7月、10月、次年1月分别采集夏、秋、冬季土壤样品。
1.3 土壤理化性质的测定
土壤含水量(SWC)的测定采用105 ℃烘干恒重法(HJ 613─2011);土壤pH值的测定采用电位法(HJ 962─2018);土壤有机质含量(SOC)的测定采用重铬酸钾氧化-分光光度法(HJ 615─2011);土壤全磷(TP)含量的测定采用碱熔-钼锑抗分光光度法(HJ 632─2011);土壤全氮(TN)含量的测定采用微量凯氏定氮法(HJ 717─2014);土壤速效磷(AP)含量的测定采用碳酸氢钠浸提-钼锑抗分光光度法(HJ 704─2014);土壤速效氮(AN)含量的测定采用纳氏比色分光光度法[20]。
1.4 土壤细菌16S rDNA的测序
采集土壤的总DNA用试剂盒(Power SoilTM DNA Isolation Kit, MOBIO, USA)提取,采用细菌通用引物338F(5'-ACTCCTACGGGAGGCAGCAG-3')和806R (5'-GGACTACHVGGGTWTCTAAT-3')对16S rDNA的V3~V4高变区进行扩增,扩增产物由上海美吉生物医药科技有限公司进行Illumina Miseq高通量测序。
1.5 数据处理
采用Excel 2013和SPSS 26.0软件对实验数据进行统计整理,显著水平确定为P<0.05。将有效序列相似性≥97%的序列划为同一分类操作单元(OTU);细菌的Alpha多样性分析采用Shannon 指数、辛普森(Simpson)指数、 Chao1指数,覆盖度(Coverage)则进行测序深度分析;对于Beta多样性分析,利用OTUs之间的系统发生关系,采用Anosim相似性分析探索不同分组样本间群落组成的相似性或差异性;采用RDP Classifier进行分类水平统计,用相关性热图进行理化指标和细菌群落关系的分析。
2 结果与分析
2.1 细菌群落多样性分析
2.1.1 序列统计和细菌Alpha多样性分析 经高通量测序,从坡豪湖湿地土壤8个样地共获得基因序列1200836条,平均长度为439.19 bp。表1显示了8个样地(YD1~YD8)的土壤细菌在不同季节的多样性指数(Shannon)和丰富度指数(Chao),由表1可知:土壤细菌群落多样性在大部分样地中以春季最为丰富;而样地YD1的土壤细菌多样性以冬季最丰富,该样地处于水陆交替带,鸭跖草、莲子草密度大,在冬季蓄水期该样地不会被淹没,因此植物生长旺盛,土壤有机质含量增加,有助于土壤微生物的生长和繁殖;样地YD4和YD5的土壤细菌群落多样性以夏季最丰富,这2个样地均处于农田旁,夏季处于放水期但雨水充沛,作物根系分泌旺盛,有机质等营养物质容易被土壤微生物利用,因此土壤细菌群落的多样性和丰度增加;此外,样品文库的覆盖率(Coverage)范围为95.4%~97.2%,能够反映土壤微生物群落结构与多样性的真实情况。
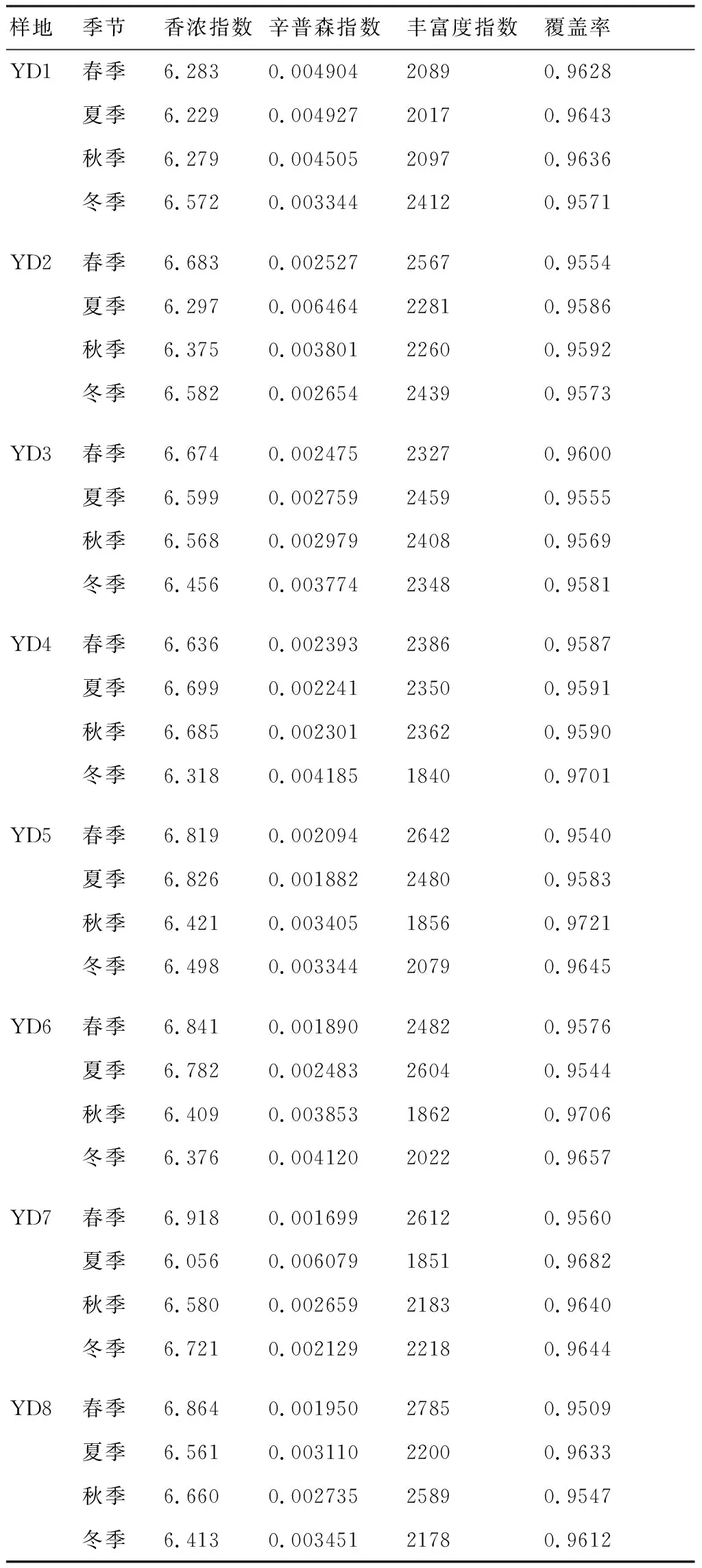
表1 坡豪湖湿地土壤细菌群落的丰度和多样性
2.1.2 Beta多样性分析 为了分析8种不同生境下的土壤细菌群落结构差异,在OTU水平上进行主成分分析,结果如图1所示,YD1在四季中(CYD1、XYD1、QYD1、DYD1)距离相近,说明土壤OTU组成差异不大,但区别于其他样地;夏季的YD2、YD7土壤OTU组成明显区别于其他样本;YD4在冬季的OTU组成区别于其在另外3个季节的;YD5在秋、冬季的土壤OTU组成明显不同于春、夏季,但QYD5与YD6在秋、冬季的土壤OTU组成相近,DYD5与DYD8的土壤OTU组成也相近。由此说明,土壤样本的细菌群落结构存在季节性差异,且在不同区域的样本间也有一定的差异性。

图中样地名称的首字母C、X、Q、D分别表示春季、夏季、秋季、冬季,下同。
2.2 细菌群落的物种组成分析
坡豪湖湿地土壤样品的4105个OTUs分属于38个门、92个纲、198个目、367个科、625个属。由图2可以看出,在门分类水平上,4个季节的土壤细菌群落组成有一定的相似性,但细菌类别占比有所差异,其中,变形菌门(Proteobacteria)、绿弯菌门(Chloroflexi)、放线菌门(Actinobacteria)和酸杆菌门(Acidobacteria)具有较高的占比,分别占26.83%~28.72%、15.85%~23.85%、13.80%~18.20%和13.31%~17.68%;而硝化螺菌门(Nitrospirae)和厚壁菌门(Firmicutes)所占比例分别只有5.28%~6.69%和3.02%~5.26%。变形菌门是坡豪湖湿地土壤的优势类群,在四季占比变化不大;绿弯菌门在春、夏季的占比(22.95%~23.84%)高于秋、冬季(15.86%~17.99%);厚壁菌门在秋、冬季的占比(4.54%~5.26%)高于春、夏季(3.01%~3.12%),说明季节变化对这两个细菌门的影响较大。
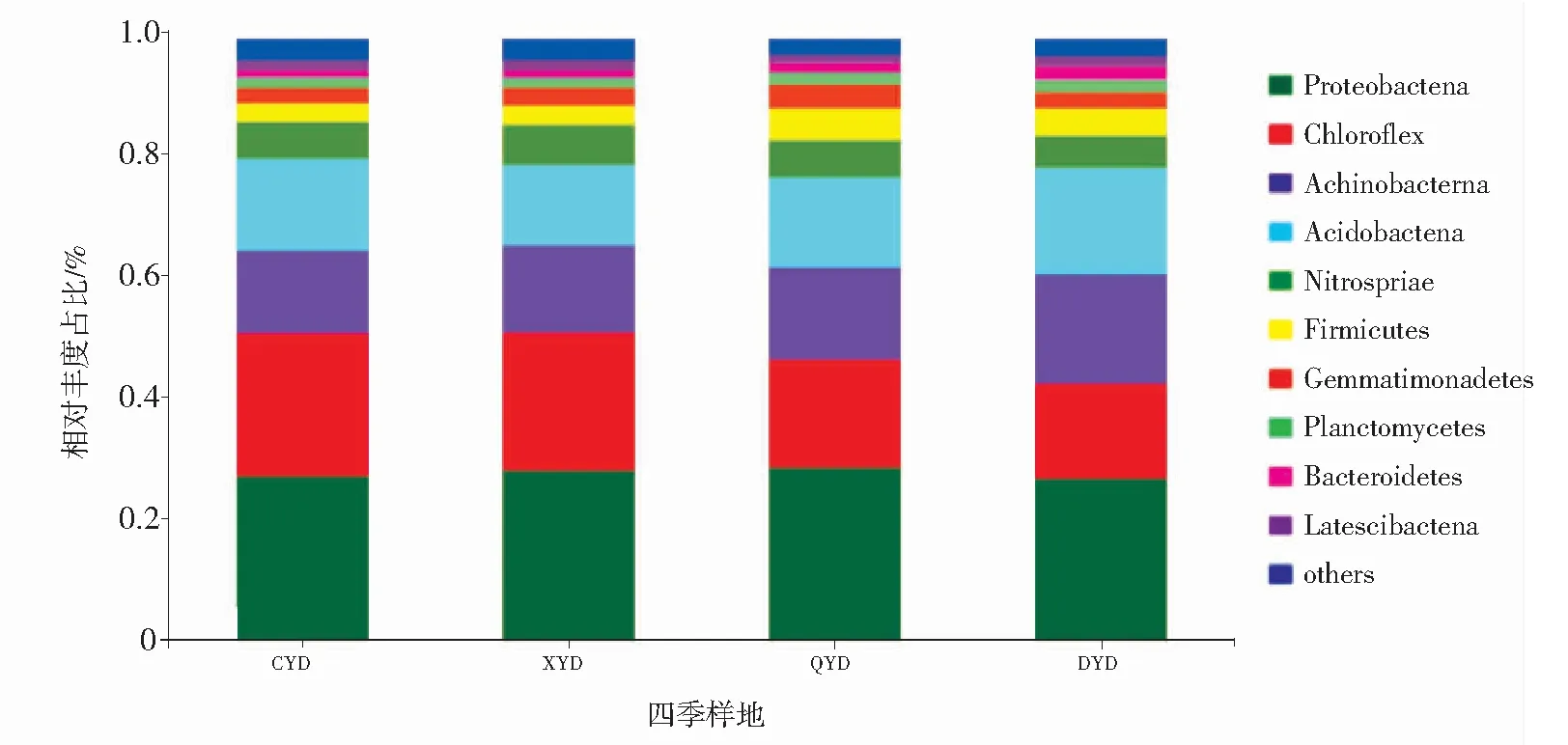
图2 坡豪湖湿地土壤细菌在门水平上的分类
为了进一步分析细菌群落结构在四季中的变化差异,在属分类学水平上进行分析,除平均丰度低于1%的部分合并为others外,共检测出29个属,如图3所示。酸杆菌门下的1个纲(norank c_Acidobacteria)的相对丰度最高,占比在冬季最大(12.64%);绿弯菌门的1个厌氧绳菌科(norank f_Anaerolineaceae)和玫瑰弯菌属(Roseiflexus)在春、夏季的占比大于秋、冬季;厚壁菌门下的芽孢杆菌属(Bacillus)在秋、冬季的占比(3.34%~2.92%)大于春、夏季(1.50%~1.60%);变形菌门下α-变形菌纲(Alphaproteobacteria)的黄色杆菌科(norank f_Xanthobacteraceae)在冬季的占比明显增大。
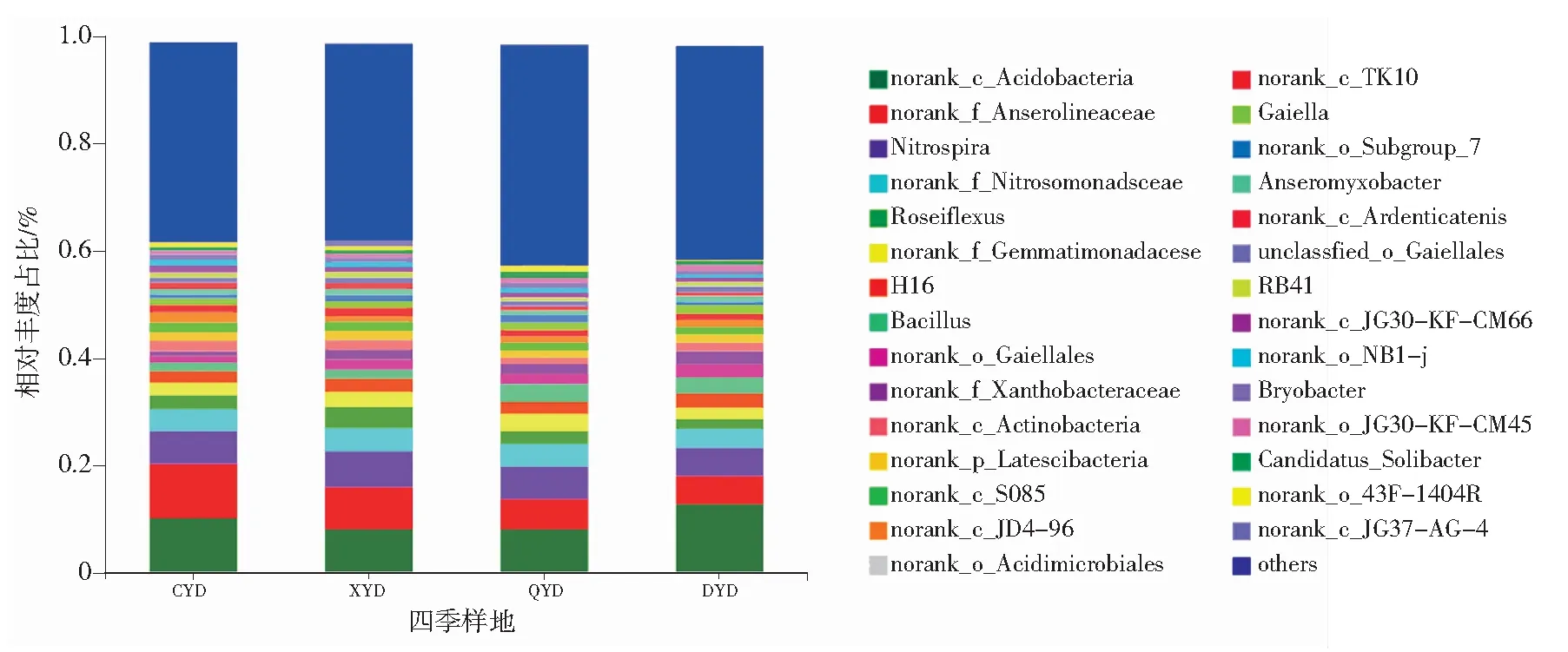
图3 坡豪湖湿地土壤细菌在属水平上的分类
对丰富度较高的6个细菌门类群进行季节性变化的比较分析,结果如图4所示。在春季,绿弯菌门(Chloroflexi)在YD5为优势菌群,相对丰度为31.22%,也高于其在其他季节中的相对丰度;该样地处于农庄附近,地势较高,受水库蓄放水的影响较小,主要受季节性水位变化的影响,春季雨水频繁,形成潮间带区,加之人为活动,影响了土壤的有机质含量,使得绿弯菌门的相对丰度较高。在夏季,变形菌门(Proteobacteria)在YD4、YD8为优势菌群,相对丰度分别为35.12%、39.06%;绿弯菌门在YD2、YD6为优势菌群,在夏季的相对丰度最大,分别为30.11%、28.52%。在秋季,酸杆菌门(Acidobacteria)在YD4、YD5的相对丰度最大,分别达20.65%、19.14%;厚壁菌门(Firmicutes)在YD2、YD8的相对丰度变化至最大,在YD2中从春季的1.45%升高到秋季的6.82%,在YD8中从夏季的2.34%升高到秋季的9.11%。在冬季,酸杆菌门在YD2、YD3的相对丰度最大,分别为23.48%和23.09%,为这两个样地的主要优势菌群;放线菌门(Actinobacteria)在YD5的相对丰度随季节变化而增大,在冬季最大,为23.29%;厚壁菌门在YD6的相对丰度以夏季最低,为0.96%,到冬季升至8.26%。

图4 坡豪湖湿地土壤优势细菌在四季的变化比较
2.3 细菌群落与土壤因子的相关性分析
由表2可知,坡豪湖湿地土壤呈弱碱性,季节性差别较大,在YD1、YD3、YD8中变化显著(P<0.05)。SPSS 26.0 ANOVA分析表明,土壤理化指标对不同季节和不同地势区域的样地有明显响应。从季节性层面看,SWC在秋、冬季显著大于春、夏季(P<0.05),因为秋冬季是下游水库蓄水期,大部分样地水位提升;TP含量在YD4内部季节波动性较大,可能与其处于农田附近,受人为因素干扰有关;大部分样地的TP含量在夏季低,因为夏季为放水期,加上降雨量增加,导致磷素流失;TN含量在冬季最高,且显著高于夏季的(P<0.05);多数样地的AP含量在秋季高,但YD4的AP含量在春季显著高于其他季节的(P<0.05),YD8的AP含量在冬季显著高于其他季节的(P<0.05);夏季在YD2中的AN含量远超其他样地的(P<0.05),达34.43 mg/kg;多数样地的SOM含量在冬季高,在夏季低,因为蓄水期使有机质积累[21],但在YD6中的SOM含量在春夏季较高。
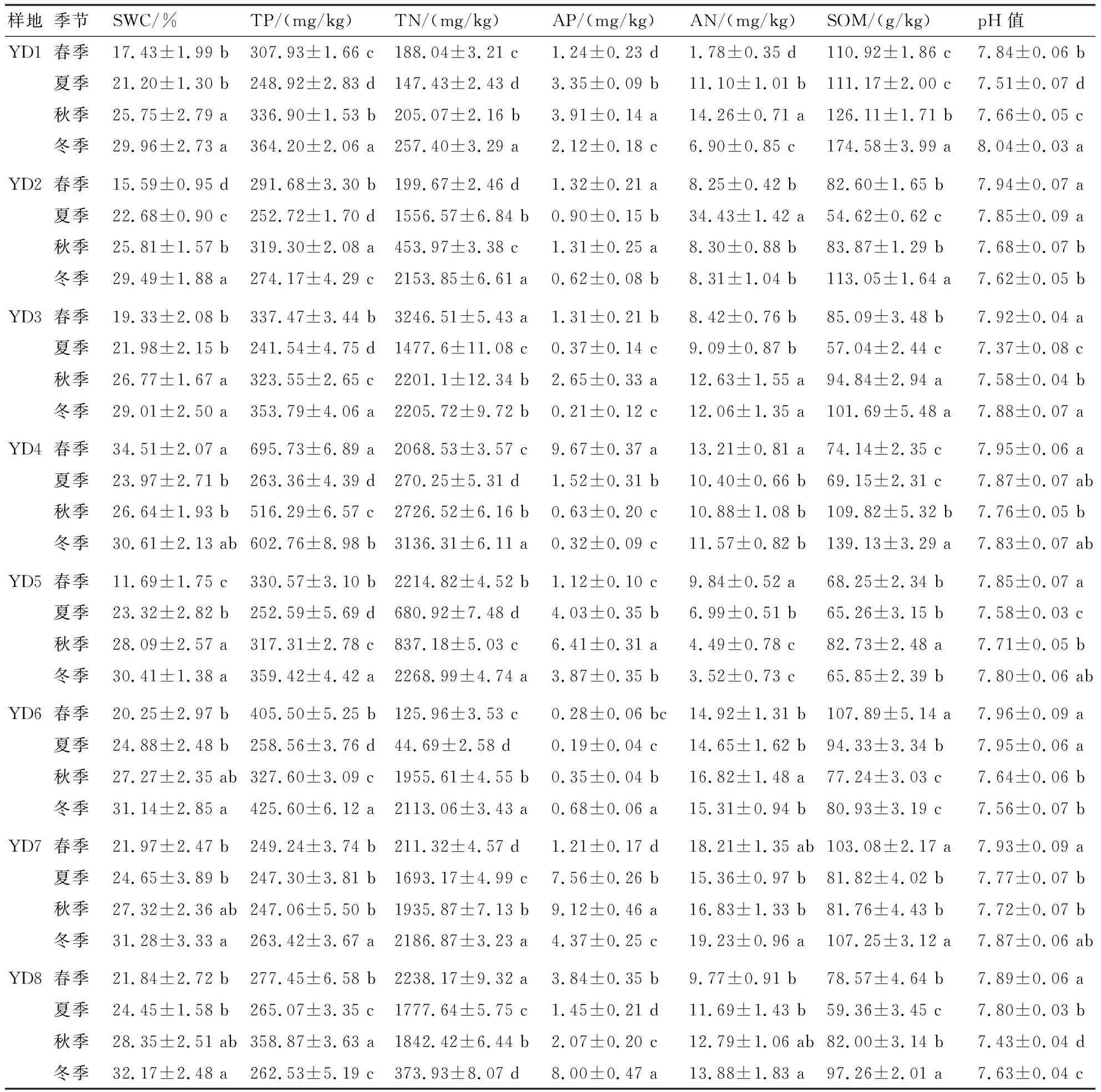
表2 坡豪湖湿地土壤理化指标的测定结果
Spearman相关性分析结果(表3、图5、图6)显示:SMC为主要影响因子,与多种细菌显著相关,其中与放线菌门(Actinobacteria)、绿弯菌门(Chloroflexi)、纤维杆菌门(Fibrobacteres)均呈极显著相关(P<0.01),其中SMC与放线菌门呈正相关,相关系数为0.605,与绿弯菌门呈负相关,相关系数为-0.695;pH、TN、SOM、AP也与一些细菌群落呈显著相关,其中pH与硝化刺菌门(Nitrospinae)呈极显著正相关(P<0.01),与厚壁菌门(Firmicutes)呈显著负相关(P<0.05);TN与拟杆菌门(Bacteroidetes)呈极显著正相关(P<0.01),与绿弯菌门呈显著负相关(P<0.05);SOM与螺旋体菌门(Saccharibacteria)呈显著负相关;AP与放线菌门呈显著正相关;TP、AN与细菌群落无显著相关性。

图5 坡豪湖湿地土壤细菌门分类群落与土壤化学指标的冗余分析结果
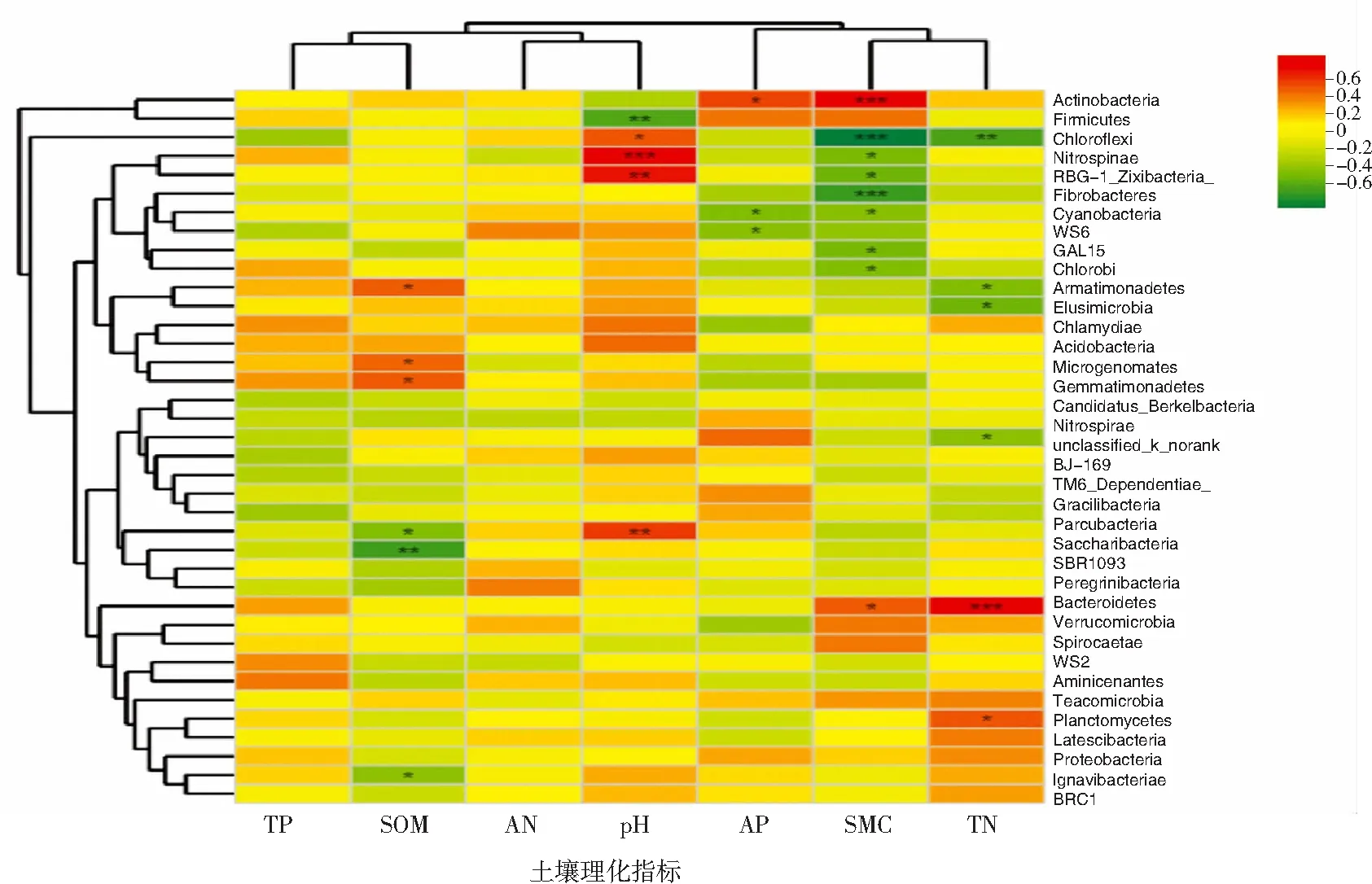
“*”表示0.01 在湿地生态系统中,微生物群落的多样性和相对丰度受温度、时间、空间和理化因子的多重影响。在分析微生物群落结构与环境因子间的相关性时,很难将某个环境因子独立分开,因为微生物群落的变化与多种因子耦合作用相关[22]。坡豪湖常年受下游电站蓄放水的影响,季节性蓄放水形成水分的反复迁移影响了湿地土壤的理化环境,继而对细菌群落的组成造成一定的影响。从细菌的多样性和丰富度结果看,本研究区的8个样地中土壤样本,春夏季为水库放水期,部分湖边土壤水位下降,但也有些区域地势低洼,加上春季雨水密集,形成长时间季节性淹水,如YD6在春季有机质含量最高,这与土壤的通透性较差、有机质积累有关,导致细菌多样性和丰富度较大。本研究发现,秋冬季的SOM含量较春夏季高,可能由秋冬季受蓄水期的影响所致。水分是影响土壤性质的最主要因子[10,23-24],样地水位高程不同导致土壤含水量、pH值、有机质含量等有明显的差异。细菌群落变化在水位影响较明显的样地中呈现差别,主要集中在靠近农田区域的YD4、YD5样地,虽然其部分区域地势较高,受蓄放水的影响较小,但也受人类活动的干扰,因此其土壤细菌群落的多样性和丰富度在夏季和秋季较大;YD2、YD8为桑树种植区域,受蓄放水和耕作的影响,其土壤细菌群落的多样性在春季最大;YD3为湖中岛,蓄放水使小岛边际的水位常处于动态变化,导致春夏季的土壤细菌多样性大于秋冬季,这与LIGI等[25]、XU等[26]、AMAUD等[27]的研究结果“在湿地生态系统中,处于不同水文条件下的土壤微生物群落结构存在着显著差异”一致。 在本研究的样地中,土壤主要的细菌群落有变形菌门、绿弯菌门、放线菌门、酸杆菌门、硝化螺菌门和厚壁菌门。其中变形菌门为相对丰度最高的类群,这与很多湿地土壤细菌群落研究[28-30]的结果一致,其中YD4和YD8在夏季变形菌门的相对丰度最大,这可能因为变形菌门与碳利用呈正相关[31],这两个区域为种植区,根系利用有机质活跃,加上耕作机械扰动促进变形菌门的碳矿化速率增大,促进了作物的生长。在属水平的物种组成中,绿弯菌门部分属的相对丰度在春夏季增大,究其原因,虽然春夏季为放水期,但春夏季雨水频繁,使部分地势处于水陆交替带,而光合自养微生物绿弯菌门常处于泥滩带,能提供大量的生产力[32],这也间接说明水分在样地中对绿弯菌门菌群的影响较大;厚壁菌门的芽孢杆菌属在秋冬季的相对丰度大于春夏季,其中在YD2、YD6、YD8中从春季到冬季丰度呈明显的增大趋势,可能与其形成可逆性强的芽孢有关,WANG等[32]在对海洋沉积物、淡水和潮间带沉积物微生物群落的研究中发现厚壁菌门的芽孢杆菌纲丰度较高,其产芽孢具有极强的环境适应能力。 相关性分析发现,土壤水分对绿弯菌门和放线菌门具有极显著的影响,绿弯菌门的相对丰度在YD5中春季最高,在YD2和YD6中夏季较高,因为这3类样地受季节性蓄放水的影响较大,这与物种组成的研究结果一致,再次证明绿弯菌门的相对丰度与土壤的水分含量相关。放线菌门的相对丰度在YD5中冬季增大,而该样地在冬季的土壤含水量较高,这与放线菌适于在干燥、高温环境下生长[33]相悖,因此其丰度增大可能还与该区域的人为因素影响有关,生活污水及垃圾的排放和堆积致使放线菌门菌群数量增加,有研究发现在污染石油、多环芳烃(PAHs)及有机物等环境中放线菌门的相对丰度增加[34-36],与其降解污染物的能力相关[37,38]。酸杆菌门的相对丰度在YD2、YD3中冬季升至最大,因为这2个样地受人为耕作和牲畜因素的干扰,土壤有机质容易被分解,这与相关研究[39-41]的结果“酸杆菌门喜欢低营养组环境”一致。有研究认为土壤酸杆菌门的丰度与pH值成反比[31];但Zhang等[44]认为大多数细菌门的相对丰度在近中性土壤中高于在酸性或碱性土壤中,其中酸杆菌门也属优势菌门,本研究结果则与其一致。由此看出,环境因素对湿地土壤微生物分布的影响很大,因坡豪湖湿地受水库的影响,其土壤细菌多样性呈季节性变化,维系物质循环,但同时湖边部分区域受耕作或污染因素的干扰,使得一些菌群的组成变化与人类活动因素相关。因此从保护湿地的生态功能考虑,应尽量控制和减少人为不利干扰的影响,以改善土壤微生物群落的结构和土壤结构。3 讨论与结论
