耦合CBEM的脉冲神经元模型建模与仿真
2021-11-13张嘉瑛贺兴时于青林
张嘉瑛,贺兴时,于青林
(1.西安工程大学 理学院, 陕西 西安 710048;2.汤普森河大学 数学与统计学系, 加拿大 甘露市 V2C 0C8)
0 引 言
脉冲神经网络[1](spiking neural network,SNN)被认为是第三代神经网络,有着广泛的研究价值。神经元细胞是神经系统能够处理复杂信息的物质基础[2-4],这些神经元之间的相互连接,形成神经系统各种复杂的结构和功能。脉冲神经元的主要生物物理模型有Hodgkin Huxley模型[5]、Lzhikevich模型[6]、LIF模型[7]、SRM模型[8]等。这些模型可以解释真实神经元的巨大动态库[9],但对于神经编码和计算的理论分析来说,不够清晰简单,激发了对神经脉冲响应的简化模型的需求。
神经编码的研究目的是揭示感觉信息是如何在神经脉冲反应中编码。编码需要了解刺激和脉冲序列之间的统计关系,以及进行这种转换的生物物理机制。PARK等从一系列阶段描述编码过程,即线性滤波、非线性变换开始,并以噪声或有条件的泊松脉冲结束,形成泊松GLM等模型[10]。然而,级联式描述性统计模型与生物物理解释模型之间存在很大差距。文献[11-12]认为在实际神经元中,刺激是非线性的,而非线性是由兴奋性和抑制性突触输入之间的相互作用引起的。这些突触输入非线性地依赖于刺激,输入反过来驱动电导变化,从而改变控制膜电位的非线性动力学。2011—2014年,MENG等介绍了标准泊松GLM的准生物物理解释,即具有相同和相反兴奋性和抑制性调节约束电导模型的等价性。放松这些约束,以获得一个更灵活和更现实的基于电导的模型,独立调整兴奋性和抑制性输入[13-14]。2014年,LATIMER正式提出了基于电导的编码模型(conductance-based neural encoding model,CBEM)[15]。文献[16-18]在生物物理框架中明确定义模型组件,并直接将模型预测与单个细胞的激发和抑制调节进行比较。CBEM在预测新刺激的视网膜脉冲反应方面优于标准GLM。这些差异突出了CBEM的能力,揭示了感官神经元可以在自然环境中进行计算。2019年,LATIMER等通过来自猕猴视网膜神经节的细胞内记录,证明了CBEM可以适合细胞外的脉冲序列数据,然后用于预测兴奋性和抑制性的突触电流[19]。由简单神经元组成的神经网络,其功能并非单个神经元功能的简单叠加。以上所建立的脉冲神经元模型均为单神经元模型,而生物体细胞是相互影响,相互耦合的。
为了考虑生物细胞序列的实际特性,缩小数学模型与生物物理模型之间的差距,本文对以上CBEM进行耦合,构建耦合脉冲神经元模型,并进行仿真实验观察细胞脉冲序列的变化。耦合的CBEM具有良好的点过程似然,使得其更易于拟合细胞外的数据,能够更多地考虑来自其他细胞的信息。该模型不仅具有独立的兴奋和抑制电导,而且根据不同脉冲序列间的相关关系,增加了耦合滤波器参数,获得了更多的信息进行序列拟合,构成一个更灵活、在生物物理上更可信的模型。
1 CBEM神经元细胞模型
因级联式统计模型(GLM模型[10])与生物物理的可解释模型间逐渐存在的巨大差距,LATIMER等提出了CBEM,详细地描述了数学模型和对应的生物物理模型之间的联系,并取得了较好的仿真结果[20]。
CBEM与泊松GLM相比,增加了基线参数b,如式(1)所示:
(1)

(2)
式中:Vt表示膜电位。将该模型转化为一个典型的LIF(leaky integrate and fire)模型,可以替换fr用“硬”的阈值函数,在从零到正无穷设置某个阈值跳跃的膜电位,将k设为指数衰减的过滤器参数,并设置脉冲后过滤器h为δ函数,进而导致瞬时出现峰值后膜电位重置。GLM膜电位是输入的线性函数,因此,2个模型都无法捕捉到大多数真实神经元突触输入的非线性。
由常微分方程控制的膜电位Vt的神经元模型为
gi(Vt-Ei)
(3)
式中:gl为漏导;ge(t)和gi(t)为兴奋性和抑制性突触电导;El、Ee和Ei分别为漏性、兴奋性和抑制性反向电位。式(3)要满足2个条件:ge(t)+gi(t)=c,c为常数;ge(t)Ee+gi(t)Ei是刺激的线性函数。根据以上条件,构造了满足这些条件的CBEM。通过将兴奋性电导和抑制性电导建模为刺激的仿射函数,由正负相反的线性滤波器驱动,即
(4)
式中:kc为线性电导滤波器;be、bi分别为兴奋性与抑制性电导的基线参数。总电导是定值g=gl+be+bi,因此将膜电位式(4)改写为

(5)
式中:b=beEe+biEi;V0=b/g。瞬时膜电位的表达式为
Vt=k·xt+V0
(6)
式中:k相当于线性卷积kc的指数衰减滤波,
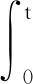
(7)
CBEM示意图如图1所示。

图1 基于电导的模型CBEM示意图Fig.1 Schematic diagram of conductauce based neural encoding model(CBEM)
图1中,CBEM的参数由刺激滤波器组成。刺激滤波器分为抑制滤波器和兴奋滤波器2个部分,以便更好地拟合对不同刺激类型的选择性过滤作用。脉冲后的过滤器h获得对脉冲历史信息,以及在没有输入的情况下确定基线发射率的基线b。对这些滤波器的输出求和,并且通过一个非线性函数fr,得到非齐次泊松脉冲过程的条件强度。
脉冲CBEM模型为
(8)
式中:ke和ki分别是驱动兴奋性电导和抑制性电导的线性滤波器;fg是保证电导非负的软整流非线性函数。加入对脉冲历史数据的考虑,并加入线性自回归项,和GLM的历史一致,得到膜电位方程,即
(9)

(10)
式中:μ是一个“软”脉冲阈值;α和β分别决定了非线性函数的斜率和锐度。
2 耦合CBEM神经元细胞模型
CBEM只考虑了单个神经元对刺激的脉冲响应过程。通过对滤波器进行耦合,加入神经元间的相互信息,可以扩展到多个神经元之间耦合的模型。图2为2个神经元耦合的示意图。
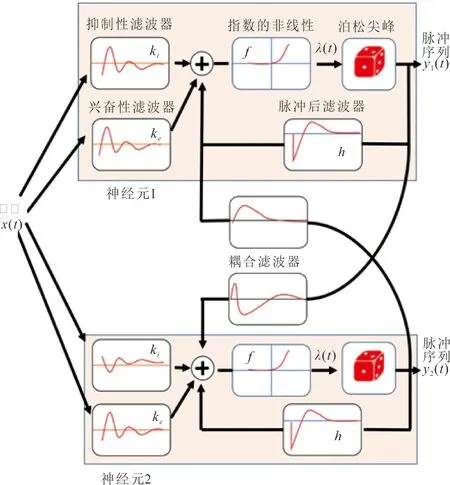
图2 互相耦合的2个神经元模型Fig.2 Coupling of two neuron models
图2在图1的基础上由单神经元模型变化为耦合神经元模型。此时的滤波器不仅是兴奋性滤波器与抑制性滤波器,还根据不同脉冲序列之间的相关关系,增加了耦合滤波器以及脉冲后滤波器,通过对滤波器的输出求和并通过非线性函数fr,得到了非齐次泊松脉冲过程的条件强度。

(11)

由MENSI等提出生物物理驱动输出的非线性函数,将膜电位转换为脉冲率,膜电位与脉冲率的关系如式(12)[21]:
(12)
式中:μ是一个“软”脉冲阈值;α和β分别决定了非线性函数的斜率和锐度。得到脉冲率的表达式为
(13)
用最大似然进行模型拟合,并计算模型参数。
3 仿真实验与分析
3.1 数据预处理
引用数据源为文献[22]发布的神经节细胞脉冲序列数据;数据组根据测试细胞神经元的种类分为on中心型及off中心型等2种。
展示的数据使用数据组中提供的第3个神经元细胞,其状态被标记为on-cell。对数据统计信息进行计算,求解到每个时间块(time bin)大小为8.3 ms,不同时间的刺激序列为144 051个,总时间长度为20 min。脉冲响应序列的个数为49 954,其平均脉冲率为41.6 Hz。
图3为原始数据在每个时间块内的可视化图,展示了对每个时间块内的脉冲次数的统计结果。这个数据作为比对细胞脉冲序列的脉冲率真值。

图3 划分的时间块内的脉冲次数Fig.3 The number of spikes in each time block
3.2 可视化结果
对4个细胞的脉冲序列两两之间进行了互相关计算,得到了互相关关系图,见图4。脉冲序列是一种二元的时间序列,从互相关关系图中可以得知2个时间序列之间的相关程度。图4中,在某些时间点上,不同脉冲序列之间存在相关联系,显示了不同神经元脉冲序列之间有着时间上的某种关联。模拟比较所提出的模型,模型中的相关关系由不同神经元间的脉冲历史滤波器实现,这些耦合滤波器和抑制、兴奋的滤波器一起构建设计矩阵。使用设计矩阵进行拟合,提出的模型与其他模型的脉冲率预测可视化结果。

(a) 细胞(1,1) (b) 细胞(1,2) (c) 细胞(1,3) (d) 细胞(1,4)
将提出的耦合CBEM与GLM(历史滤波器)[23]、单神经元CBEM[20],泊松GLM[10]、NP-GLM[24-25]等4种模型进行脉冲率预测对比,结果如图5所示。

(a) 细胞1 (b) 细胞2
从图5可以看出,提出的耦合CBEM神经元模型取得了更好的脉冲拟合效果,模型拟合预测的脉冲率更加符合实际的脉冲率。相比于GLM(历史滤波器)与泊松GLM,耦合CBEM对高频的脉冲响应拟合效果更好,在较高的脉冲率时较为明显,且在脉冲率较低时保持了很好的拟合效果;相比于NP-GLM,耦合CBEM在低频的脉冲响应拟合效果更好,而且高频时也更接近于脉冲率真值;相比于单神经元CBEM,耦合CBEM采用了多个神经元的联合信息,耦合后对拟合效果产生了很大的影响。
3.3 结果分析
对模型的实验验证采用了单脉冲(single-spike)信息[26]和AIC信息准则[27]等2个量化指标。单脉冲信息[26]定量比较这些模型更详细地考虑时间为单一脉冲的简单情况,即计算单个脉冲传递的平均信息I1:
(14)
式中:r(t)为瞬时脉冲率;T为单个观察窗口的时间。单个脉冲所携带的信息应该与脉冲速率相关。速率作为时间的函数,给出了脉冲序列的“单一体”统计的完整描述。表1为使用Matlab计算的关于几种模型的单脉冲信息比较结果,单位为比特/脉冲(bits/spike)。

表1 单脉冲信息比较Tab.1 Comparison of single-spike information
从表1可以看出:相对于其他4种模型,提出的耦合CBEM 4组实验的单脉冲信息都有所提升。虽然在细胞3中耦合CBEM仅次于NP-GLM,但是其他实验数据的增长非常明显,表明单个脉冲传递的平均信息增多。耦合CBEM在细胞序列的拟合方面,携带了更多的信息。不同神经元耦合对脉冲序列有较大的影响。耦合CBEM使用数据的增加容易导致模型编码信息随之增多,所以计算模型的AIC(akaike information criterion)值[18]要考虑模型的复杂程度。
AIC信息准则[27]是衡量统计模型拟合优良性的一种标准。该准则建立在熵的概念基础上,可以权衡所估计模型的复杂度和此模型拟合数据的优良性。在一般的情况下,AIC可以表示为
AIC=2k-2lnL
(15)
式中:k为参数的数量;L为似然函数。AIC准则鼓励数据拟合的优良性同时惩罚模型的过度拟合,所以优先考虑的模型应是AIC值最小的那一个。表2为几种模型AIC值的计算结果。

表2 AIC计算结果Tab.2 Calculation results of AIC
耦合CBEM是简化的单个CBEM加入其他细胞历史信息,在此基础上增加耦合滤波器,简化了模型的相对复杂度。由表2可以看出:耦合的CBEM相对于其他模型AIC值都有所降低。虽然细胞3的实验中复杂度略高于NP-GLM,但是其他几组实验中仍然低于其他模型,且相对于单个CBEM的模型复杂度大大降低,获得了较好的拟合效果。
4 结 语
本文提出了一种脉冲神经元建模方法,考虑不同脉冲神经序列数据间存在的相关关系,加入耦合滤波器对CBEM进行改进。实验结果表明,提出的耦合CBEM对脉冲序列的拟合效果更好。因耦合滤波器,信息得以在不同脉冲神经元中传递,使得模型承载了更多的信息,在整体上降低了模型复杂度。加入了耦合滤波器,仍保留了CBEM在生物物理上的可解释性。以后的工作将进一步探索大规模脉冲神经网络中脉冲神经元间耦合机制,研究相应的高效优化方法。
