弓形虫分泌蛋白转运机制的研究进展
2021-11-11李爽,刘群
李 爽,刘 群
(中国农业大学动物医学院 国家动物寄生原虫实验室,北京 海淀 100193)
弓形虫(Toxoplasmagondii)是一种重要的人兽共患机会性病原,专性细胞内寄生,几乎感染所有的温血动物及人类,危害严重。弓形虫在中间宿主体内主要有2种存在形式,即速殖子和缓殖子,缓殖子外被囊壁,以包囊形式存在[1]。速殖子的入侵、增殖和逸出是弓形虫传播必不可少的过程,任何一个步骤出现异常都不能使其正常寄生。
弓形虫的顺利入侵、增殖和逸出主要依赖于其3种独特的分泌细胞器,即微线体、棒状体和致密颗粒。虫体入侵时,首先产生微线体蛋白(Microneme proteins,MICs)和棒状体颈部蛋白(Rhoptry neck proteins,RONs)将弓形虫锚定在宿主细胞膜上,随后穿过细胞膜进入宿主细胞,释放棒状体蛋白(Rhoptry proteins,ROPs)和致密颗粒蛋白(Dense granular proteins,GRAs),形成纳虫空泡(Parasitophorous vacuole,PV)。ROPs可调控宿主细胞的免疫应答促使宿主细胞提供有利环境维持自身增殖和生存;部分GRAs被分泌到PV参与形成纳米微管-囊泡网络,摄入生长必需养分并调控自身抗原的暴露,也可以分泌到宿主细胞,调节宿主基因表达,触发宿主免疫反应[2]。
这些分泌蛋白是维持虫体正常生命活动和传播的必要分子,其中有些是弓形虫重要的毒力因子,因此阐明弓形虫分泌细胞器从头合成的分子机制和分泌蛋白加工转运途径是近数十年来的研究热点,加上最近开发的众多分子生物学研究手段,促进了弓形虫作为顶复门原虫蛋白质运输和细胞器生物发生模型的研究。因此,本文将对分泌蛋白的合成、分类和运输的研究进展做一综述。
游客们对三峡地区历史文化的认知主要是巴蜀文化和巫文化。原始的巴人部落、舞蹈表演和神秘的巫文化祭祀活动构成了游客们对三峡地区古代文明的体验,帮助游客更好地理解遥远的历史文化。一位广西的年轻游客描述了印象深刻的巫文化祭祀表演:
1 弓形虫分泌系统亚细胞结构
弓形虫具有一个内质网(Endoplasmic reticulum,ER)与核膜相连。高尔基体(Golgi)由3~5个折叠的腔室堆积而成,向外分泌延伸[3]。分泌蛋白在核糖体合成后有序的从内质网运输到高尔基体,再从高尔基体流向内体样的腔室,包括早期内体(Early endosome,EE)和晚期内体(Late endosome,LE),经过蛋白酶水解修饰,形成棒状体和微线体的前体蛋白[4]。弓形虫的LE附近还存在一个充满大量蛋白酶的植物样液泡(Plant-like vacuole,PLV)[5],其中组织蛋白酶L(Cathepsin L,CPL)介导了微线前体蛋白的水解成熟。这些现象表明,弓形虫具有独特的内体系统(Endosomal-like compartments,ELC),与弓形虫增殖过程中顶端分泌细胞器的生物发生和蛋白功能的发挥直接相关。
2.4 两组患者术后排尿情况比较 术后6个月研究组患者排尿潴留、排尿困难、尿失禁、尿急及尿频的发生率均显著低于对照组患者,差异有统计学意义(χ2=7.350,P<0.05)。见表5。
2 弓形虫分泌蛋白的生物合成
除了这些分泌蛋白自身的保守基序和结构域在蛋白转运过程中起作用外,还有很多调控因子发挥着核心功能。目前,鉴定出部分弓形虫蛋白分泌过程中囊泡出芽、运输和融合机制的相关因子。主要包括以下几种。
Rab蛋白:Rabs是最大的小G蛋白家族,在囊泡运输中起分子开关作用,是囊泡从供体到受体定向运输所必需的[18]。弓形虫只存在一组基本的Rab蛋白,TgRab5A定位于EE,Rab5A和Rab5C过表达导致微线体和棒状体转运缺陷,ROPs被重新运输到PV中。此外,过表达TgRab11A或TgRab11B都可以导致虫体分裂异常,新合成的内膜复合体(Inner membrane complex,IMC)从高尔基体向子代细胞的运输和分泌受阻,表明TgRab11A和TgRab11B调节组成性分泌[19]。
2.3 致密颗粒蛋白 致密颗粒为球形致密细胞器,直径约200 nm,不同感染阶段虫体中的致密颗粒数量不同[16]。
2.2 棒状体蛋白 棒状体的位置与微线体几乎一致[10],共有8~12个,其生物合成由高尔基体的囊泡出芽驱动,在TGN和正在分裂的虫体顶端之间形成大的囊泡,不断发育成熟[11],但其成熟途径和机制尚不完全清楚。
ROPs的分选机制与MICs非常相似,可溶性ROPs可携带多种不同的靶向信号[12]。ROP2和ROP4都具有一个跨膜结构域,基于酪氨酸的分选信号(YXXΦ基序)有助于将蛋白质分类到棒状体[13]。任何一个基序发生缺失或突变都会阻断蛋白向成熟棒状体的转运,导致它们在类似多泡体(Multivesicular bodies,MVB)的大囊泡中积累[14]。这些数据表明,ROPs从TGN到早期内体可能以复合物的形式运输,依赖于不同蛋白的分类信号。事实上,后来鉴定TgSORTLR是ROPs转运的关键受体,条件性敲除后导致蛋白定位错误,成熟异常[15]。
本文以理论计算和数值模拟相结合的方法,得出CFG桩复合地基的公式计算沉降量和有限元分析结果相符。首先对CFG桩复合地基的承载性状进行了理论研究,再利用MIDAS有限元软件,分析了褥垫层厚度、置换率对桩复合地基的承载特性的影响。主要结论如下:
吴浈长时间在江西省卫生系统工作。他曾任江西省卫生厅医教科技处干部;1989年-2000年,任江西省卫生厅药政管理局副局长、局长。
综上所述,我们在组织衔接教学时,一定要把握住学生学习过程中所暴露的问题,要针对性地给出解决方案,这样才能让我们的教学更有效率.
3 弓形虫蛋白分泌转运的关键因子
2.1 微线体蛋白 微线体位于弓形虫顶部100~150 μm的位置,表达多种MICs,在分泌途径早期组装成复合体,不同蛋白之间的相互作用是它们从反式高尔基体网(Trans-golgi network,TGN)靶向到微线体的必要手段[6]。
GRAs与形成宿主细胞器隔离管状结构(Host organelle sequestering tubular structures H.O.S.T.)和膜状纳米管腔网络结构(Membranous nanotubular intravacuolar network,IVN)有关,前者调节宿主内体和溶酶体向PV的传递,后者连接PV内的虫体和PV膜[17]。与高等生物的真核细胞不同,弓形虫中从未观察到未成熟的致密颗粒。目前发现引起GRAs转运异常的蛋白还很少,并且新合成的GRAs从TGN转运到致密颗粒的分子机制目前尚不清楚,需要更多的研究来阐明其分泌途径。
TgMIC6是一种跨膜蛋白,也是TgMIC4和TgMIC1的转运蛋白,这种联系通过MIC6的特征性生长表皮因子(Epidermal growth factor,EGF)样结构域介导[7],与TgMIC1的类半乳糖凝集素结构域(Galactose-like agglutinin domain)相互作用,其中SYHYY和EIEYE两个酪氨酸的基序可以对该复合物进行正确的分类[8]。TgMIC2与TgM2AP在弓形虫表面以复合物的形式分泌,有助于滑移运动和对宿主细胞附着。M2AP前肽发挥分类作用,决定了分泌蛋白的靶向性[9]。复合物的每个组成部分都具有不同的分类信号,它们分泌方式和功能的差异直接与分泌细胞器生物合成过程中的不同转运途径有关,但这些不同的MICs是否依赖于不同的分类机制仍有待确定。
可溶性N-乙基马来酰亚胺敏感因子附着蛋白受体蛋白(SNAREs):定位于内体系统(ELC)和高尔基体(Golgi)之间,促进囊泡融合传递货物蛋白。3种突触蛋白synaptobrevin/VAMP、syntaxin和SNAP-25形成复合物共同构成突触小泡与质膜融合机制的核心[20]。弓形虫存在23种SNARE样蛋白,其中Stx6定位于TGN、Golgi和ELC之间以及虫体表面。过表达后虫体发育畸形,ELC扩张,Golgi破碎,IMC成熟异常,并破坏蛋白从ELC到高尔基体的运输[21]。
网格蛋白衔接复合物(Clathrin adaptor complexes):在真核细胞中有6种AP复合物,功能不同[22]。AP-1在TGN到内体的顺行和逆行中起作用,破坏TgAP1 μ1亚基导致MICs在TGN水平上的分类错误,使可溶性MIC3重新进入PV,而跨膜MIC8主要保留在TGN中。此外,TgAP1可以通过激活TGN中的ROPs转运来调节棒状体的生物发生,缺失TgAPμ1导致成熟的棒状体破碎,未成熟的ROPs被重新运输到PV或残体中[23]。因此,TgAP1可能在pro-ROP从TGN中运出的过程和未成熟棒状体向成熟棒状体的转变过程中发挥作用[24]。
囊泡分选蛋白(Vacuole protein sorting,VPS):酵母和哺乳动物细胞中,内溶酶体系统包含2个同源的系留复合体:内体Class C core vacuole/endosome tethering(CORVET)和Homotypic fusion and vacuole protein sorting(vacuolar HOPS),这些复合物对于内体从早期到晚期的转变、溶酶体生物发生和内溶酶体运输途径至关重要[25]。TgSORTLR也称VPS10,定位于高尔基体-内体相关区室内,具有2个关键的结构域。N末端TgSORTLR内腔结构域与ROPs和MICs特异性相互作用,异位表达导致自身蛋白以及棒状体和微线体蛋白均定位错误;而胞质尾部结构域则参与蛋白质的顺行和逆行运输。因此,TgSORTLR是弓形虫分泌细胞器生物发生和蛋白转运的关键。TgVps11是 HOPS和CORVET装配的支架蛋白,与囊泡和未成熟分泌细胞器有关[15]。随后,研究人员在弓形虫中又发现了多个VPS蛋白,包括VPS4、VPS18、VPS39和VPS9等,它们均参与调节分泌细胞器的生物发生[26]。
动力蛋白相关蛋白B(Dynamin-related protein,DrpB):动力蛋白是一种大的GTPase,通过充当机械酶来切断囊泡之间的联系,参与细胞器分裂或囊泡运输[27]。弓形虫中存在3个动力相关蛋白,分别是DrpA、DrpB和DrpC,其中TgDrpB定位在Golgi附近,通过调节分泌途径中的囊泡运输促进分泌细胞器的形成,条件性敲除DrpB导致成熟的子代虫体微线体和棒状体丢失[28]。
4 展望
弓形虫利用其内体系统发挥分泌转运蛋白的功能,使MICs和ROPs高效有序的沿着TGN到ELC的路线通向顶端分泌细胞器,其中多种蛋白因子参与调控,从而确保蛋白精确无误的靶向运输。HOPS复合体参与了分泌蛋白从TGN到未成熟细胞器的顺行运输和融合,Vps10与AP-1、AP-2、网格蛋白、Vps9和逆转录酶复合物结合,是货物蛋白顺行和逆行转运的关键受体(图1)。然而,现在还需要更多的研究来证明弓形虫早期内体向晚期内体的转化机制,分泌囊泡的分类途径,以及参与分泌蛋白转运成熟的相关蛋白及其作用机制。此外,GRA蛋白对弓形虫的毒力和生存至关重要,调节致密颗粒生物发生和蛋白分泌的机制仍然是弓形虫分泌蛋白研究的重点难点,还未有探索。因此,对于揭示弓形虫分泌蛋白的运输成熟仍需大量研究。
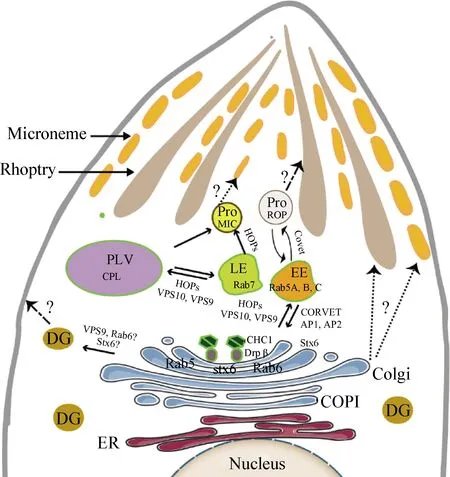
图1 弓形虫分泌蛋白转运模式
