蓝舌病在全球的流行及分布
2021-11-11罗倩敏韩佃刚杨云庆叶玲玲尹尚莲李瑶瑶何昌霖董仙兰
罗倩敏,韩佃刚,杨云庆,董 俊,祝 贺,叶玲玲,张 冲,尹尚莲,李瑶瑶,何昌霖,董仙兰,艾 军
(昆明海关技术中心,云南昆明 650200)
蓝舌病(bluetongue,BT)是由呼肠孤病毒科环状病毒属蓝舌病病毒(bluetongue virus,BTV)引起的一种烈性传染病,其发病严重程度取决于宿主和BTV 血清型[1]。该病的临床特点是发热、面部水肿、流涎、口腔黏膜溃疡、舌发绀,流浓涕以及蹄叶和蹄冠发炎。绵羊对该病最易感,病死率一般为2%~30%,有时可达70%;牛为亚临床感染,症状不明显或无症状,是BTV 的携带者[2-3]。
20 世纪初,BT 首先在非洲流行,1902 年南非最早详细记录了BT;1905 年证实BT 由病毒引起,1944 年证实库蠓是BT 的传播媒介;1924 年BT 侵入塞浦路斯,至1977 年连续发生多起疫情;20 世纪50 年代BT 相继传播至美洲、欧洲、大洋洲;到20 世纪60 年代,BT 已在全球范围内流行,导致大量感染动物死亡,每年经济损失达百亿美元。2006 年,欧洲出现了毒力更强的BTV-8 型,引发严重疫情,波及大半个欧洲,导致大量绵羊死亡,牛也出现明显症状甚至死亡。据不完全统计,2006—2008 年,BTV-8 感染动物86 806 头,造成数十亿欧元的损失[3-4]。
1979 年,我国云南省师宗县首次发生BT 疫情,并从患畜体内分离到BTV 毒株。其后,湖北、安徽、四川和山西等地也先后报道发生BT。我国分离到BTV 血清型有16 种,包括BTV-1~5、7、9、11、12、14~17、21、24 和29[3-5]。本文回溯了BT 的全球流行及分布情况,分析了流行因素,以期为BT防控提供参考。
1 病原学特征及传播媒介
1.1 病原学特征
BTV 由10 个节段(Seg-1~10)的双链RNA构成,其病毒粒子对称结构为二十面体,长度约80 nm[6]。BTV 病毒粒子由外衣壳、内衣壳两个蛋白质层组成,其中内衣壳包含病毒蛋白(VP1、VP4、VP6)、非结构蛋白(NS1、NS2、NS3、NS3a、NS4)和病毒基因组。其中病毒基因组由10 个dsRNA 片段及其编码基因组成,分别为Seg-1/VP1、Seg-2/VP2、Seg-3/VP3、Seg-4/VP4、Seg-5/NS1、Seg-6/VP5、Seg-7/VP7、Seg-8/NS2、Seg-9/VP6、NS4、Seg-10/NS3和NS3a[7]。其中:Seg-2/VP2变异程度最高,可诱导被感染动物产生特异性中和抗体,对病毒血清型(serotype)起决定性作用;Seg3/VP3将BTV 毒株分为两种地域型(topotype),其中中国以及南亚、东南亚和澳大利亚的BTV 毒株为东方型(Eastern),美洲、非洲、欧洲以及中东的BTV 毒株为西方型(Western)[8];Seg-7 和Seg-10 将BTV 毒株分为数种地域亚型[5]。目前全球已报道的BTV 血清型有29 种(表1)[2]。

表1 全球已知BTV 血清型分离情况
1.2 传播媒介
BT 是虫媒传染病,不会在动物之间直接传染,主要通过媒介库蠓传播。传播BT 的库蠓遍及温带和热带气候区域,主要分布在北纬40°至南纬35°。不同BTV 血清型的分布通常由库蠓的种类及栖息地决定[9]。主要病媒库蠓及分布(图1):非洲的C.imicola,欧洲的C.obsoletus、C.imicola、C.dewulfi和C.pulicaris,北美的C.sonorensis,南美的C.insignis,东亚及澳大利亚的C.brevitarsis、C.wadai,亚洲的C.brevitarsis、C.wadai和C.fulvus[9]。
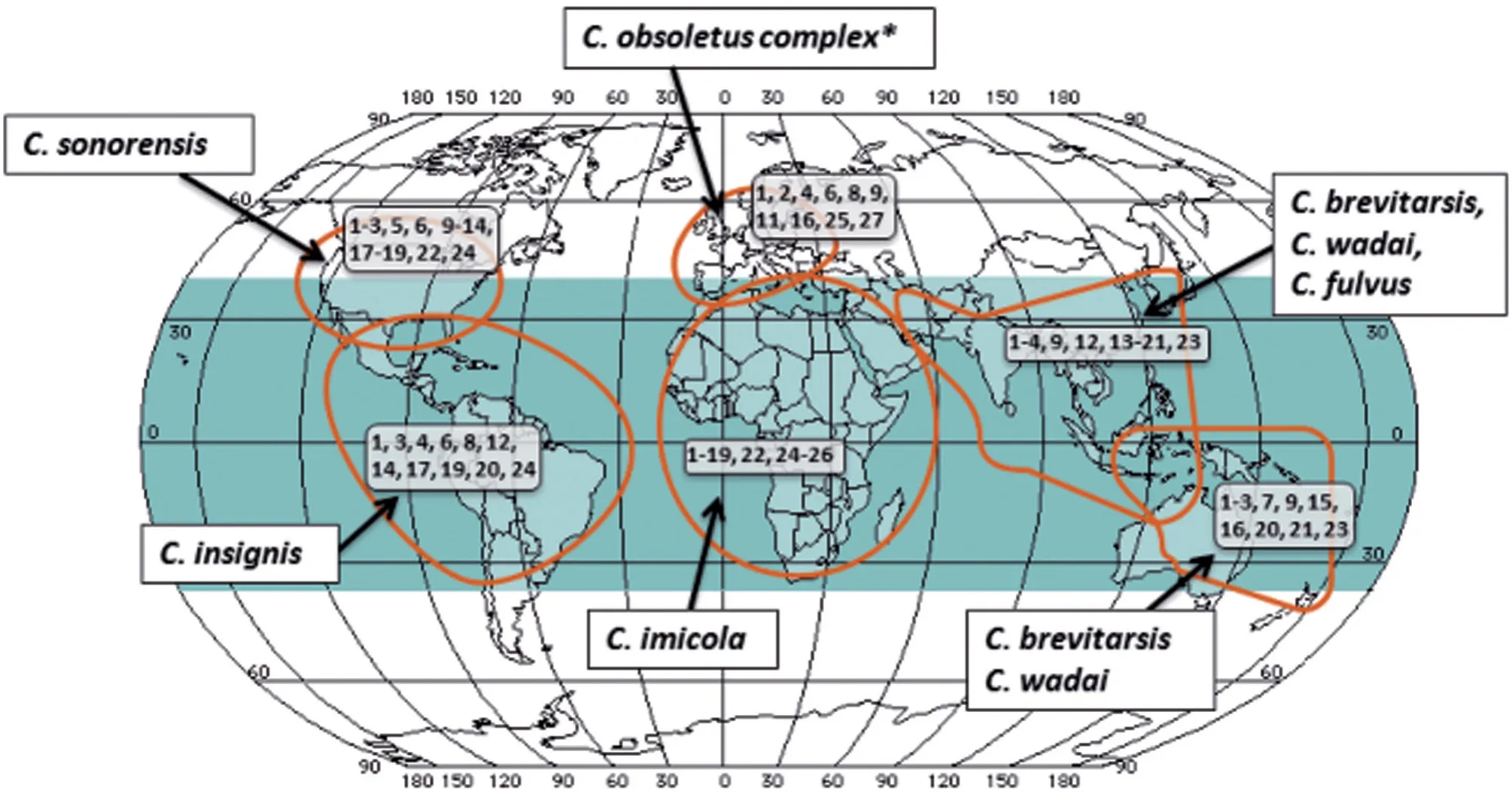
图1 BTV 血清型的大致地理分布及主要病媒[9]
2 全球流行及分布
2.1 非洲
BT 在非洲已存在100 多年,第一个关于BT的官方报告起源于18 世纪晚期南非的开普省[10]。目前29 种BTV 中有27 种已在非洲不同地区报道。BTV-2 侵入北非(阿尔及利亚、利比亚、摩洛哥和坦桑尼亚)已在2004 年得到确认[11]。Mahmoud等[12]在调查利比亚BTV 流行率和血清型分布时,检测到6 种BTV 血清型抗体,包括BTV-1~4、9和26。Drif 等[13]在摩洛哥开展了一项血清学调查,在骆驼中检测到6 种血清型,包括BTV-1、4、6、8、14 和16。
Toye 等[14]在肯尼亚西部的犊牛中检测到9 种BTV 血清型,其中BTV-7、15、16 和19 为单一感染,BTV-1、12,BTV-3、24,BTV-22、24 为混合感染。Elfatih 等[15]在苏丹中部的2 个哨牛群中检测到16种BTV 血清型中和抗体,包括BTV-1~10、12、14、17、20~22,发现苏丹中部BT 发病率与雨季和病媒种群有关,仅在7—12 月检测到。博茨瓦纳、莱索托、马达加斯加、纳米比亚、南非和津巴布韦都报告了绵羊和山羊的BT 疫情[16]。在南非分离到24 种血清型BTV-1~24[16],2013 年在南非羊驼中检测到BTV-29[17],2016 年在毛里塔尼亚的牛和单峰骆驼中发现了BTV-26 抗体[18]。1987 年,通过血清中和试验从埃及牛羊血清中鉴定出BTV-1、4、10、12 和16,2016 年发现BTV-3[19]。
这些研究表明,非洲作为BT 源发地,已存在27 种BTV 血清型。它们如何形成,如何向外传播,尚无更深入溯源报道。
2.2 欧洲
1924 年塞浦路斯首次发现BTV,随后引发多起BT 疫情,主要为BTV-3 和BTV-4 血清型,1977 年超过13%的绵羊感染BTV-4,2016 年9 月首次发现BTV-8[20]。
2006 年,荷兰马斯特里赫特地区首次检测到BTV-8,随后该血清型在西欧和中欧地区引发大规模疫情,直到2009 年才得到控制;2007 年,西班牙、葡萄牙和法国发现BTV-1;2008 年,荷兰和德国发现BTV-6,瑞士发现BTV-25;2009 年,比利时发现BTV-11;2011 年,俄罗斯发现BTV-14;2012 年,波兰发现BTV-14,这种血清型一直存在到2014 年;2012 年,意大利在撒丁岛发现BTV-4,并于2014—2017 年蔓延至整个东欧地区,波及17 个国家;2014 年,法国在科西嘉山羊中发现BTV-27[21-22]。
BTV-8 在2009—2014 年期间未在欧洲发现,2015 年在法国中部再次出现,至今仍在持续流行[18]。北欧地区BTV 血清型有6 种,包括BTV-1、6、8、11、14 和25[23]。1998—2012 年在南欧地区流行的血清型至少有6 种,包括BTV-1、2、4、8、9 和16[24]。自1902 年首次报道BT 以来至2012 年,西班牙已经历了6 次BT 暴发,由5 种血清型(BTV-1、2、4、8 和10)引起,每次暴发的严重程度和后果各不相同[25]。法属圭亚那分离到BTV-1、2、6、10、12、13、17 和24 共8 种血清型[26]。BTV 从1924 年登陆欧洲以来,已引发多次疫情。迄今,欧洲是全球BTV传播流行最大的疫区,也是疫情重灾区。截至2014 年9 月3 日,欧洲的BT 疫区(图2)分别位于意大利、西班牙、葡萄牙、马耳他、法国(科西嘉岛)、希腊、保加利亚和罗马尼亚[27]。
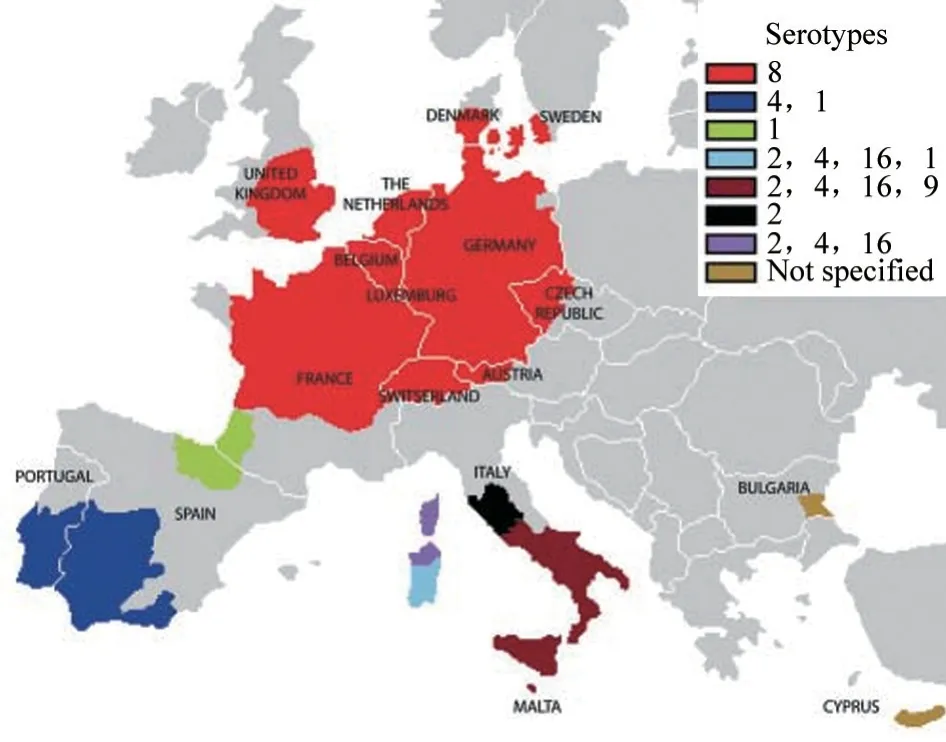
图2 欧洲各BTV 血清型疫区分布[27]
BTV 从非洲传入地中海盆地和欧洲有两条途径(图3)[28]:①东部从埃及、利比亚等地出发,可能通过牲畜交易,或以病畜携带,或以附着交通载体等方式,横越地中海,从土耳其、希腊陆路进入欧洲;②西部经北非突尼斯、阿尔及利亚、摩洛哥传入,以同样方式跨过地中海,侵入西西里、撒丁岛、塞浦路斯、科西嘉、克里特岛等地中海群岛,然后进入南欧国家。
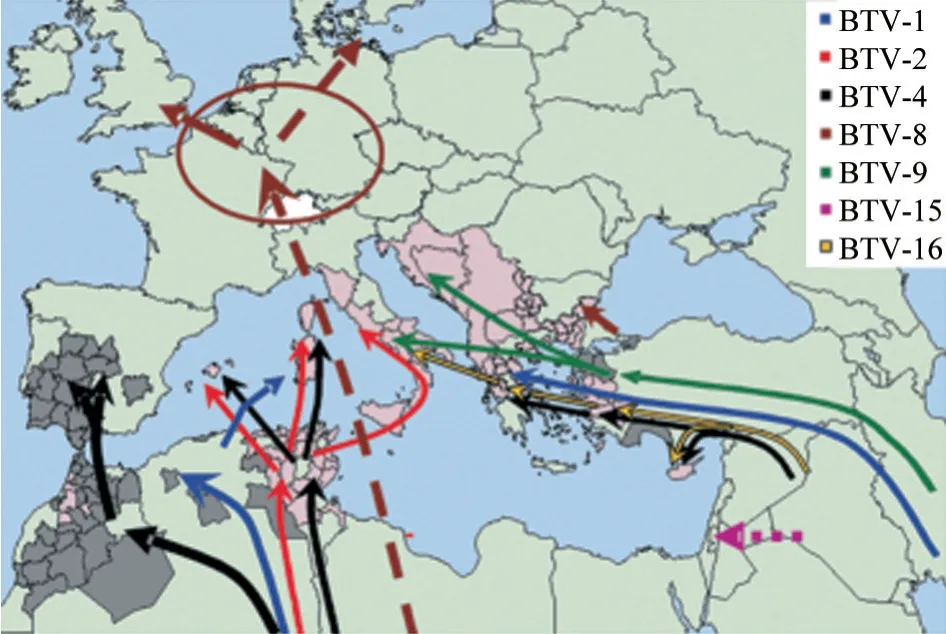
图3 欧洲各BTV 血清型传入散播途径[28]
2.3 美洲
20 世纪90 年代初,有关人员对加勒比、中美洲、南美洲地区的BTV 进行了广泛研究,确定了特立尼达和多巴哥(TTO)牛羊中的BTV 抗体高阳性率(79%),并确定了流行血清型为BTV-1、3、4、6、8、12 和17[1]。Brown-Joseph 等[1]在特立尼达和多巴哥进口牛中检测到BTV-1、2、3、5、12和17,并首次报告了BTV-2 和BTV-5 的流行。自1999 年以来,美国在东南部已经分离出11 种外来血清型(BTV-1、3、5、6、9、12、14、18、19、22 和24)[29]。
4 种血清型(BTV-10、11、13、17)长期以来在美国大部分地区流行[29]。Da 等[26]首次报告了巴西存在BTV-2、9、21 和26 的血清学证据。在中美洲和加勒比地区,20 世纪80 年代分离到BTV-1、3、4、6、8、12 和17,20 世纪90 年代分离到BTV-10、11、13、14、19 和22[30]。部分BTV 血清型传入美洲的途径:北美洲→加勒比/中美洲→南美洲(巴西、阿根廷)[31]。在南美洲、中美洲和加勒比地区的BTV 血清型流行情况见表2。

表2 在美洲地区BTV 流行血清型分布情况[31]
2.4 大洋洲
研究表明,澳大利亚的BTV 起源于东南亚和美拉尼西亚地区,从北部地区或西澳大利亚进入[32]。澳大利亚1975 年分离出BTV-20,1979 年分离出BTV-1 和BTV-21,1982—1986 年检测到BTV-3、9、15、16 和23 共5 种血清型,2007 年分离出BTV-7,2008 年分离出BTV-2,2015 年分离出BTV-5 和BTV-12[33]。澳大利亚12 种血清型BTV-1、2、3、5、7、9、12、15、16、20、21 和23 主要流行于北部地区,BTV-1、2、15 和21 流行于东部地区(图4)[32,34]。
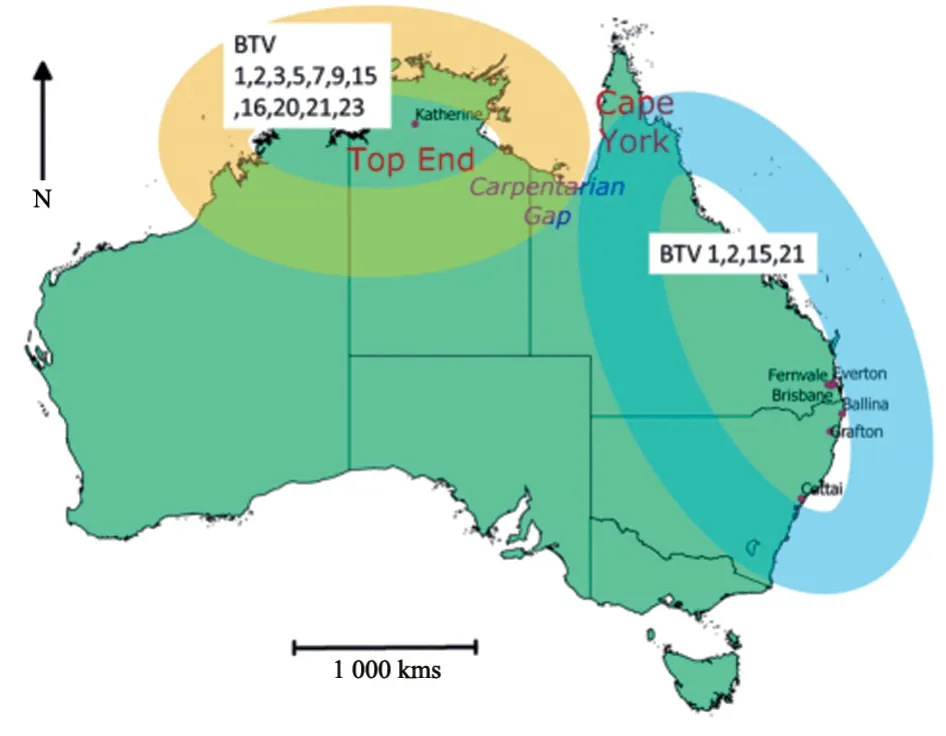
图4 澳大利亚BTV 血清型分布[34]
2.5 亚洲
传入亚洲的途径:南亚的印度首次记载BTV流行,其后在东亚、中亚、中东等地有BTV 流行的报道。
2.5.1 南亚 印度在20 世纪60—70 年代为了杂交改良本地绵羊品种,从美国和南非引进绵羊品种。1964 年印度次大陆首次报道了BT。1967—1970 年,印度从进口的无角短毛羊、兰布莱绵羊、俄罗斯美利奴羊和考力代羊中报告发现了BTV。1974 年,安得拉邦报告多塞特品种羊出现了严重的BT。自1981 年以来,印度每年都有BT 疫情报告。这些数据表明,牲畜的国际贸易可能在西方BTV 毒株传入印度方面发挥了重要作用[35]。印度已分离出23种血清型,包括BTV-1~21、23 和24[2]。Rupner等[36]通过系统综述和meta 分析,介绍了印度农场的BT 流行情况。其在2001—2018 年已发表文章的409 篇文章中筛选出71 篇,样本涉及绵羊14 048 只、山羊14 696 只、牛5 218 头、水牛2 653 头、骆驼2 062 只、大额牛222 头;分析显示,绵羊中的BT 血清阳性率为39%,山羊中为43%,牛中为31%,水牛中为35%,骆驼中为21%,大额牛中为66%。
2.5.2 中东 在以色列,自2006 年以来,多种BTV 血清型(BTV-2、4、5、6、8、12、15、16和24)在羊牛中共同传播。2008 年以色列首次报道出现BTV-8(ISR2008/13),证实它是通过从北欧进口受感染动物而传入的[20]。在黎巴嫩流行6种血清型,分别为BTV-1、4、6、8、16 和24[37]。
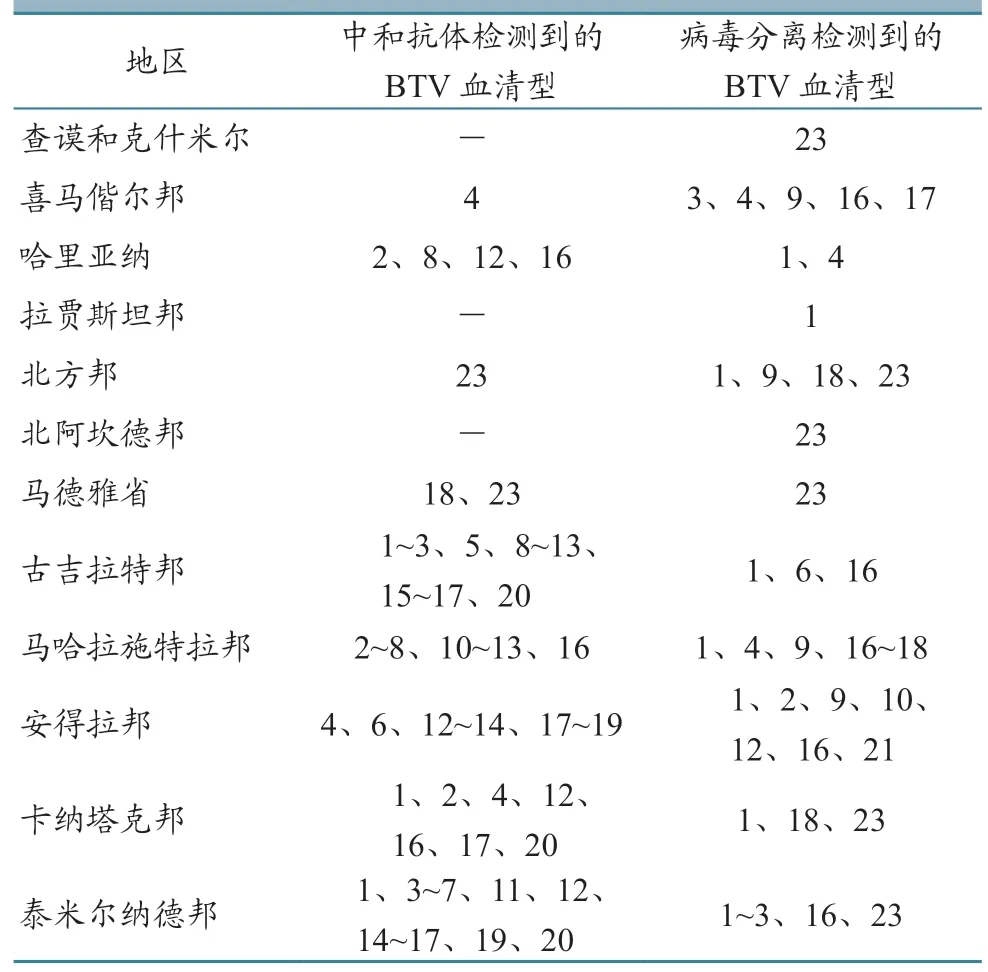
表3 印度BTV 流行情况[35]
2.5.3 东南亚 在东南亚已分离到的9 种BTV 血清型,分别为BTV-1、2、3、7、9、12、16、21和23。从印度尼西亚哨牛中分离出8 种血清型,包 括BTV-1、3、7、9、12、16、21 和23;从 马来西亚哨牛中分离出BTV-1、2、3、9、16 和23血清型[38]。Douangngeun 等[39]对老挝5 个省进行BTV 检测,在1 089 份水牛、牛和山羊样本中,发现96.7%为BTV 阳性。
2.5.4 东亚 1979 年我国首次在云南省师宗县绵羊中记录了BT 暴发,其后湖北、安徽、四川和山西等地相继暴发BT,并陆续分离到BTV-1~4、12、15 和16 型毒株,其中BTV-1 和16 已在我国广泛流行,且引起的绵羊临床症状较为明显[5]。我国目前已经分离到16 个BTV 血清型,包括BTV-1~5、7、9、11、12、14~17、21、24 和29(表4)[3]。由表4 可见:我国云南省存在11 种BTV 血清型,说明已有多种血清型BTV 侵入云南;广东省分离到6 种BTV 血清型,广东拥有外运港口,可能导致BTV 的传入与扩散;新疆、内蒙古的牧区与境外蒙古国、中亚国家陆地相连,当地牧业以饲养绵羊为主,且一直保留游牧习惯,很容易导致牛羊感染并传播BTV。在没有BTV 检出报告的地区,不能说明没有BTV 及病媒库蠓的存在,还需要进一步扩大监测范围,掌握BT 流行动态。
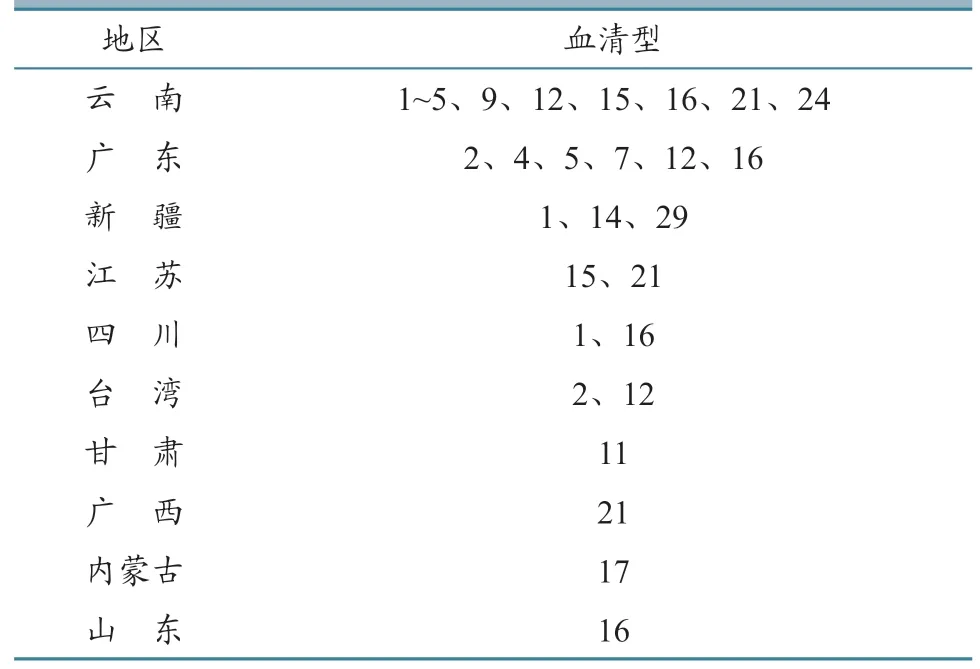
表4 BTV 在我国的分布情况
3 近年来流行形势
3.1 持续在全球流行,欧洲是重灾区
根据OIE-WAHIS 记录统计,2015—2020 年,BT 呈持续全球流行态势,但主要疫区分布在欧洲。2016 年,欧洲暴发次数多达4 234 起,随后呈逐年下降趋势,而非洲、亚洲、美洲暴发次数每年合计约几百起(图5)。2015—2020 年,欧洲累计因BT 疫情死亡牛羊25 050 头(只),其中2017 年单次死亡数高达10 714 头(只),而亚洲、非洲、美洲累计死亡牛羊分别为16 883、2 268、122 头(只)。
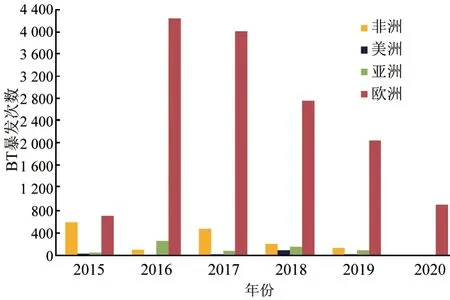
图5 2015—2020 年全球BTV 暴发次数统计
3.2 流行血清型复杂
根据OIE-WAHIS 数据可知,全球范围内因BT 疫情而造成的牛羊死亡,是由多种BTV 血清型毒株感染所致,没有出现单一BTV 血清型引发的大规模疫情。从表5 可见:2015—2020 年,非洲每年有1~9 种BTV 血清型流行,以BTV-1~4 型为主;欧洲有3~5 种,以BTV-1~4 型和8 型为主;亚洲有3~7 种,以BTV-2~4、8、16 型为主;美洲多达14种,但与其他洲不同的是BTV-4型流行较少。

表5 2015—2020 年全球BTV 血清型流行分布
4 流行因素
BT 在全球持续流行已有100 多年,主要有四大因素:易感动物移动、易感动物聚集、BTV 基因重配和传播媒介强大。
4.1 易感动物移动
BTV 需要宿主、病媒不停移动,才能接近新的易感动物完成传播。野生反刍动物随季节、水、草生长变化往返迁徙,群体数量庞大;BTV 借助病媒库蠓,伴随牛羊同行,形成一个生态圈,延续了各自物种。畜牧业BT 流行史表明,BT 传播与反刍动物的国际交易紧密相关。BT 源发于南非,当地畜种对其并不敏感,感染后症状轻微,而欧洲畜种改良后代对BTV 极为易感,首例BT 病畜记录就是欧洲引入的美利奴绵羊。2006 年研究[22]证实,在法国病畜体内分离到的BTV 与非洲南部毒株高度同源,表明它来自非洲;据统计[40],2009—2013 年,非洲94%的出口牛精液来自南部非洲,这些精液若有被感染,就可能传播给受精母牛;印度在BT 患畜体内分离到BTV 西方毒株,认为牲畜的国际贸易可能在西方BTV 毒株传入印度方面发挥了重要作用[35];研究[20]认为,2008 年以色列暴发BTV-8 疫情,也是通过从北欧进口受感染动物而造成的。牛羊交易为BTV 传播不断地提供新的易感动物群体。
4.2 易感动物聚集
BTV 的大范围传播并形成疫情,与易感动物聚集密切相关。从时间纵轴看,BT 流行始于20 世纪初欧洲工业革命时代,资本的高速扩张,带动了农业革命,使传统养殖业走向工厂化、集约化、规模化和商品化。在20 世纪中后期,大农业发展聚集了巨大数量的BT 易感动物群体,导致BTV 跟随宿主集合流动,开始了大面积流行。
4.3 血清型基因重配
BTV 是由双链RNA 组成的病毒,变异频率高。基因组分节段RNA 病毒所产生的基因重配,对病毒的免疫原性、感染特性、致病力以及传播力等都有很大影响[40]。RNA 病毒不同毒株之间能进行基因重组,产生变异毒株。研究表明基因重配是导致BTV 遗传多样性的主要原因:1987 年,美国报道BTV-10 和BTV-17 发生重配现象;2008 年,意大利报道M5 节段与BTV-2 疫苗株发生重配的BTV-16,随后发现BTV 基因重配在欧洲大陆广泛发生;印度研究发现,BTV-21 和BTV-10 发生了重配现象;2014 年发现广东汕头市BTV-7 分离株有部分节段属于Western 型,与非洲BTV 毒株有亲缘关系[41];李卓然等[5]在云南省哨兵动物中分离获得了7 种血清型,其中BTV-2、12、16 的Seg-7 属于Eastern 型,与南非和荷兰的BTV 毒株Seg-7 的亲缘关系最近。
4.4 强大的传播媒介
BT 是非接触性的虫媒传染病,库蠓是其主要媒介。库蠓栖息在温暖潮湿的低洼处或杂灌树丛处,多以吸食哺乳动物血液为生,在日出前和日落后活动频繁;专门吸食牛羊血液并能传播BTV 的库蠓有50 多种,其中少数能在野外作为有效载体[42]。它们能敏锐感应到易感动物的存在,可通过主动飞行、被动风吹、搭载运输载体等形式传播,在风中可飘行300 km 左右。任何封锁隔离手段都无法彻底阻断库蠓对易感动物的接近,以及传播BTV。
5 结语
进入21 世纪后,受全球气候变暖、畜产品贸易等因素影响,BT 流行发生变化,逐渐向地球高纬度、大范围蔓延。近年来,BT 在北欧、中欧、北美以及亚洲等多个地区暴发,这些区域以前都被认为没有BT 发生风险,现在却都遭受到BT 威胁。
BTV 虽然不感染人类,却能给人类生产活动带来巨大损失。面对BT 全球流行的严峻形势,需要采取强有力的防控措施:禁止传染源输出/输入,控制易感动物流动,改造低洼天然牧场,建设无规定动物疫病生物安全隔离区,实施有效的预防接种制度,建立强有力的动物防疫法规体系。
