川西高原垂穗披碱草根际土壤放线菌分离与鉴定
2021-11-08许斌洋杨旭赵德浚杨财容刘松青
许斌洋,杨旭,赵德浚,杨财容,刘松青
(成都师范学院 化学与生命科学学院,四川成都 611130)
放线菌是土壤微生物中的一个主要类群,其数量、种类及活性会影响土壤生态系统的结构和功能,进而影响植物的生长。与此同时,放线菌还是一类具有抑菌、抗肿瘤、酶抑制等重要活性的微生物资源,具有巨大的应用前景[1]。如临床上常用的氨基糖苷类、四环素类、糖肽类及大环内酯类抗生素等均来自放线菌[1-3]。放线菌还可以合成多种有用的代谢产物,如维生素、淀粉酶、纤维素酶和氨基酸等,与人类的生产生活关系紧密[4]。随着放线菌分离研究的不断深入,大量常见菌株已被反复研究,因此从中发现新化合物的概率逐渐降低。为了更高效地发现新的微生物资源,人们逐渐把目光投向少有研究的特殊生态环境中,如海洋、动物共生、植物内生、根际土壤以及极端环境等,从而寻找具有较大利用价值的放线菌资源[5-9]。
青藏高原平均海拔4 000 m,地势高亢,生态条件复杂,这决定了青藏高原放线菌资源具有其独特性。本研究实验材料选择川西高原的垂穗披碱草根际土壤,目前对该地区较为常见的垂穗披碱草根际土壤放线菌的系统研究较少。而垂穗披碱草又是川西高原的优势草种,分布广泛,可能是由于放线菌产生活性物质,与垂穗披碱草互利共生,相互影响、相互作用。通过对放线菌的形态特征观察、生理生化鉴定、16S rRNA基因扩增比对以及系统发育树的构建,筛选出拮抗病原菌的放线菌,为垂穗披碱草牧草的生长生产提供理论依据,这对川西高原畜牧业的发展和生态环境保护具有重要意义。
1 材料与方法
1.1 材料
1.1.1 土壤样品
实验材料的土壤样品共6份,分别采自川西高原的查真梁子(经纬度为N32°67'25''、E102°11'59'',海拔4 345 m)、红原草原(经纬度为N32°34'22'、E102°23'12',海拔3 920 m)、红原草甸(经纬度为 N32° 90'18'、E101° 15'53'',海 拔 3 595 m)、安曲乡(经纬度为 N32° 66'21'、E102° 26'17',海拔3 455 m)、查理乡(经纬度为N32°76'34'、E102°05'49',海拔3 299 m)以及马塘沟(经纬度为N31° 53'62'、E102° 37'43',海拔 3 100 m),编号依次为1、2、3、4、5、6。采集土样均为0~12 cm的表层土样,分别用无菌袋装好放于实验室4 ℃冰箱保存。
1.1.2 培养基
改良高氏一号培养基、l/10BeIlllette agar培养基、腐殖酸培养基、改良淀粉脯氨酸培养基、低营养矿物盐培养基以及高氏一号培养基。
1.2 仪器
LDZX-50KBS型立式压力蒸汽灭菌锅,上海申安医疗器械厂;BCD-248WBSV型立式冷藏箱,青岛海尔股份有限公司;PYX-DHS型隔水式电热恒温培养箱,上海跃进医疗器械有限公司;SJ-CJ-1F型超净工作台,苏州苏洁净化设备有限公司;JA3003精密电子天平, 上海良平仪器仪表有限公司;SKY-100C型恒温培养振荡器,上海苏坤实业有限公司。
1.3 方法
1.3.1 菌种分离
采用土壤悬浮液稀释涂布平板法分离菌种。从混匀的各备用土样中随机称取1 g自然风干的垂穗披碱草根际土壤,在研钵中研磨,通过100 ℃高温1 h烘干,在准备好的带有磁珠的99 mL无菌水中低速摇床(170 r/min,30 ℃)处理1 h,再微波处理30 s混匀后得到稀释100倍的土壤悬浮液。取1 mL稀释100倍的土壤悬浮液加入到9 mL无菌水的试管中,混合摇匀得到稀释1 000倍的土壤悬浮液。取1 mL稀释1 000倍的土壤悬浮液同样加入到9 mL无菌水的试管中,混合摇匀后得到稀释10 000倍的土壤悬浮液。取摇匀后的土样悬液0.2 mL,涂布于分离平板上,28℃培养2周,得到单菌落。
1.3.2 菌种纯化和保藏
通过28 ℃温度条件下在15 d的培养皿上挑选单个菌落,在改良高氏一号培养基平板上接种,用平板划线法纯化菌株,并挑取纯菌株的单菌落接种于改良的高氏一号培养基的斜面上作为菌种的初步保藏。将纯化后的菌株用高氏一号液体培养基在28 ℃,摇床中转速170 r/min下培养3~5 d,配制好浓度的50%的甘油,将甘油与菌液按体积比1∶1(V∶V)混合,保藏于-80 ℃冰箱中。
1.3.3 放线菌的形态学鉴定
将保藏于-80 ℃的菌株活化,在改良高氏一号培养基进行插片培养法进行观察,并记录肉眼可观察的放线菌菌落特征,如基内菌丝颜色、气生菌丝颜色、菌落表面含水状态以及色素产生情况等。
1.3.4 放线菌对病原菌的抑菌活性的测定
1.3.4.1 菌悬液的制备
将病原菌大肠杆菌、金黄色葡萄球菌、铜绿假单胞杆菌、枯草芽孢杆菌4株菌接种于牛肉膏蛋白胨培养基,37 ℃活化24 h,然后液体培养得到菌悬液。
1.3.4.2 抑菌活性的测定
每个培养皿加入100 μL病原菌的菌悬液,均匀涂布,通过牛津杯法[10],每皿内竖放3个牛津杯,每个牛津杯加入200 μL放线菌的样品制备液(每个重复3次)。
1.3.5 生理生化鉴定
(1)唯一碳源的利用:鉴定实验菌株对不同碳源的利用情况。(2)明胶液化试验:测定菌种产生蛋白酶的能力。(3)硝酸盐还原试验:测定菌种对硝酸盐的还原能力。(4)纤维素分解试验:测定菌种产生纤维素酶的能力。(5)硫化氢的产生:检测菌株生长过程中是否产生H2S[11-13]。
1.3.6 DNA的提取及16S rRNA基因PCR扩增
采用细菌DNA抽提试剂盒(上海生工生物股份有限公司)提取基因组DNA,扩增引物为通用引物,扩增产物进行测序,在NCBI上进行序列比对。
2 结果与分析
2.1 形态观察及培养特征
从川西高原的6个土壤样品中分离5株放线菌,分别命名为YX1、YX2、YX3、YX4、YX5,观察菌落形态和菌丝形态,可以发现5株菌落均为圆形,有皱褶,其表面干燥,菌落周围有辐射。菌株的菌丝粗细不一,长短不同,分支多而且孢子丝不易观察,结果如表1。

表1 土壤放线菌的菌落特征与菌丝形态
2.2 土壤放线菌的抑菌活性的测定
通过牛津杯法测定菌株发酵液的抑菌活性,结果见表2。5株菌中,YX1、YX2和YX5的抑菌效果较突出,YX1能够抑制大肠杆菌、铜绿假单胞菌、枯草芽孢杆菌的生长,其中对大肠杆菌的抑制效果较好;YX2能够抑制大肠杆菌、金黄色葡萄球菌,其中对大肠杆菌的抑制效果较好;YX3只对枯草芽孢杆菌有抑菌效果;YX4只对铜绿假单胞菌有抑菌效果;YX5能够抑制大肠杆菌、金黄色葡萄球菌、枯草芽孢杆菌的生长,其中对大肠杆菌的抑制效果较好,对枯草芽孢杆菌抑菌效果较低敏。

表2 土壤放线菌发酵液对4种病原菌抑制效果
2.3 土壤放线菌的生理生化鉴定
参照放线菌生理生化鉴定方法对5株放线菌进行生理生化鉴定,鉴定结果如表3。

表3 各菌株生理生化鉴定结果
由表3可以看出,所有的菌株均能使淀粉水解,都不能产生H2S和黑色素,均能使明胶液化和牛奶胨化,均能利用葡萄糖,均不能利用蔗糖,只有菌株YX3可以使纤维素分解,只有菌株YX5不能利用果糖,只有YX1和YX3能使硝酸盐还原。
2.4 土壤放线菌16S rRNA基因PCR扩增结果
用试剂盒法提取菌株的DNA,经过PCR扩增,得到菌株16S rRNA基因序列片段,用琼脂糖凝胶电泳仪对扩增产物进行检测,凝胶成像仪进行观察拍照,发现1 500 bp相应位置都有均一、明亮条带出现,扩增结果如图1,这表明PCR扩增成功,低温保存。

图1 PCR扩增产物琼脂糖凝胶电泳
2.5 土壤放线菌系统发育分析
将PCR产物纯化后直接送至上海生工生物股份有限公司进行测序。根据16S rRNA测序结果,登陆NCBI在线查询,对YX1~YX5的序列进行Blast分析(采用16S rRNA序列对 GenBank中的数据库进行搜索),找到其中序列相似性最高的序列(表4),在菌株的16S rRNA序列同源性高达95%时可以认为属同一属,当序列同源性高达97%以上可认为属于同一种[14]。通过5株分离菌株的16S rRNA序列测序结果及其同源菌株在GenBank 中公布的16S rRNA 序列,利用Mega 5软件采用Neighbor-joining法构建出系统发育树,结果见图2。由系统发育进化树分析可以看出,YX1、YX2、YX4与Streptomyces rishiriensis(NR_044141.1)和Streptomyces rishiriensis(NR_112392.1)亲缘关系较近;YX3与Streptomyces sindenensis(NR_115441.1)聚在一起,可能属于该菌株;YX5与Streptomyces alboviridis(NR_115374.1)亲缘关系较近。
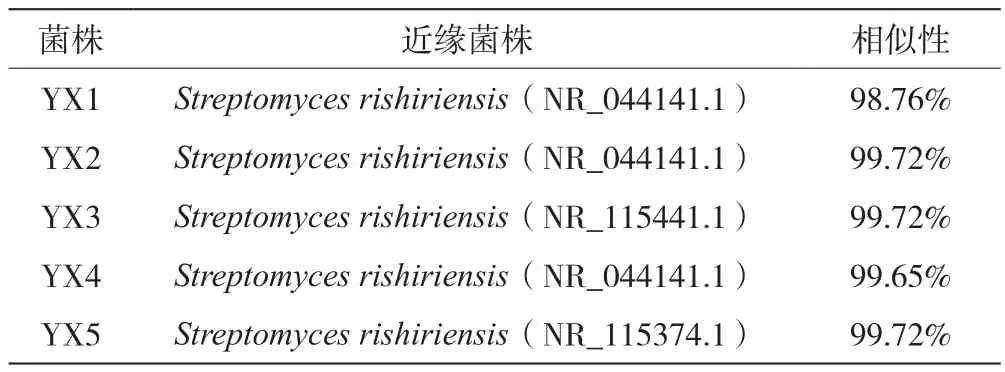
表4 5株菌株16S rRNA基因序列比对结果

图2 基于邻接法构建的系统发育树
根据16S rRNA序列同源性分析结果显示,结合生理生化特征和系统发育分析,将YX1、YX2、YX4鉴 定为Streptomyces rishiriensis;YX3鉴 定 为Streptomyces sindenensis;YX5鉴定为Streptomyces alboviridis;YX1~YX5均为链霉菌属(Streptomyces)。
3 讨论
本次实验从川西高原地区6份土壤中分离纯化得到了5株放线菌,通过对5株放线菌的形态特征观察、生理生化鉴定和16S rRNA基因扩增比对以及系统发育树的构建,最终鉴定出这5株菌的属,均为链霉菌属(Streptomyces)。链霉菌是产生抗生素活性物质最多的一类放线菌,由前人的研究可知,迄今已知的抗生素中,有51%是由链霉菌产生的[15]。不少抗生素已用于防治植物上的有害病菌,如井冈霉素、春雷霉素等用于防治植物真菌性病害[16]。在本研究中可以发现,在川西高原地区存在重要的具有进一步研究开发价值的放线菌资源,尤其是链霉菌资源,是农用和畜牧业抗生素产生菌的重要分离资源和种质资源库,对于发展畜牧业和生态环境建设有着重要的研究价值。
本研究在分离根际土壤放线菌的基础上对放线菌进行了生理生化鉴定,可以发现,5株菌株的淀粉水解能力、明胶液化能力、牛奶胨化能力均较强,而纤维素分解能力较弱,仅只有YX3能够使纤维素分解,5株菌株不能产H2S,说明均无分解含硫有机物的能力。这与来航线等[17]分离的放线菌生理生化试验结果相似。这些菌株对于淀粉酶、蛋白酶、纤维素酶的开发应用提供了研究材料,后期可进一步详细研究菌株的产酶条件等相关酶学性质,对其进行处理改造提高酶的活力和产量,为酶工业生产提供更多微生物资源。
本研究发现土壤中放线菌种类和数量较少,但根据叶景静等[18]在广西茅尾海红树林植物根际土壤中发现放线菌共88株,有6目8科9属;罗亚军等[19]在西藏沙棘根瘤及根际土壤中分离放线菌共112株,有7目12科23属,而本次实验只分离出5株链霉菌属的菌株,说明土壤中还有更多的放线菌还未分离出来,樊炳君等[20]通过分离桉树根际土壤放线菌研究发现,不同培养基上放线菌的生长情况和数量差异很大。其中,以采样地土壤制成的培养基放线菌的出菌率最高,推测可能是由于桉树大量吸收养分而形成的贫营养环境的选择作用使得适应寡营养的放线菌类群占据了绝对优势的原因。未来将通过增加筛选培养基的种类和改变抑制剂的种类和配比,来改善放线菌数量和种类较少的问题,争取从川西高原筛选到更多有价值的放线菌。罗亚军等[19]对西藏沙棘根瘤及根际土壤放线菌进行抗菌活性研究发现,有17株菌至少对一种检定菌有抗菌活性;潘文娟等[21]对西藏湖泊放线菌进行抗菌活性测定发现,有55株菌至少对一种检定菌有抗菌活性,这表明放线菌能产生抗菌活性物质,抑制病原菌生长。本次研究通过牛津杯法对放线菌进行4种病原菌的抑菌活性的测定,发现YX1、YX2和YX5的抑菌效果较好。这些具有抑菌抗性的放线菌还可以进一步研究其促生功能、抗逆性作用、功能基因以及抗菌活性产物与其药用价值间的关系,全面而系统地揭示川西高原垂穗披碱草根际土壤放线菌的药用价值与应用前景,达到保护垂穗披碱草资源的目的。因此,在后续的研究工作中,可进一步对这些菌株的抗菌活性进行测定,研究其产生抗菌活性的物质,揭示其次生代谢产物的化学组成,力求从中发现具有较好应用价值的新型活性物质,为新药的开发提供依据。另可针对禾本科常见的病原菌筛选出较好拮抗活性的微生物资源并制作相应产品进行应用,将有助于川西高原草场的改良,有利于促进我国的畜牧业发展和环境保护事业。
