自然贮藏对‘莫拉特Ⅱ’杂交臂形草种子萌发及抗氧化酶活性的影响*
2021-11-05蒋金娟朱德荣王鸿发罗富成
蒋金娟,王 琰,朱德荣,徐 翠,王鸿发,罗富成
(1.云南农业大学 动物科学技术学院,云南 昆明 650201;2.普洱市思茅区畜牧工作站,云南 普洱 665000)
种子休眠是指在预定的时间范围内,有生命活力体征的种子在利于其萌发的环境因子的组合条件下,本该萌发但没有发芽的现象[1-4]。许多植物的种子都具有这种现象,这是植物在长期进化过程中抵抗不良环境条件所形成的一种生态适应性,是植物适应环境的一种生存策略[5-8]。休眠的种子播种后不能及时出苗,导致田间出苗断垄现象时有发生,数年后又沦为田间杂草,这已成为草业生产严重的限制因素,是亟须解决的问题。
杂交臂形草 (Brachiaria hybrid) 为禾本科臂形草属多年生密丛型草本植物,是中国热带、亚热带地区优良的放牧及刈割兼用型牧草[9-10]。该牧草喜温暖潮湿气候,耐高温、耐干旱、耐贫瘠、耐酸,亦较耐寒[10-12],且生长速度快、产量高。但其种子落粒性强,发芽率低,休眠程度深,严重制约了该种子的推广应用[13-14]。王琰等[9]采用机械处理破除杂交臂形草种子休眠,发现去除稃片可显著提高种子的发芽率和活力指数,发芽率达到76.00%,活力指数为0.20,分别是对照的2.05 和1 034.78 倍。PEREIRA 等[15]研究发现:在42 ℃和98%的相对湿度下,长达48 h的加速老化有助于‘莫拉特Ⅱ’杂交臂形草种子的发芽和α-淀粉酶活性的提高。目前,关于杂交臂形草的相关研究主要集中在引种栽培和种子生产技术上,关于种子休眠方面的研究报道很少,其休眠的自然释放及休眠期的长短对生产影响很大,研究又尚未涉及。本试验以自然贮藏0~24 个月的‘莫拉特Ⅱ’杂交臂形草的净种子为试验材料,每2 个月取样1 次,测定其发芽率、活力水平及抗氧化酶活性,研究自然贮藏过程中种子活力及抗氧化酶活性的变化规律,旨在探明其休眠的自然释放过程,确定其休眠期的长短,为该优良牧草的推广应用提供理论依据。
1 材料与方法
1.1 试验材料
试验以云南省普洱市生产的‘莫拉特Ⅱ’杂交臂形草大田种子为原始材料。种子采收后经风选获得净种子,然后将其充分混匀,并装入尼龙袋,存放于干燥、通风的储藏室内自然贮藏0~24 个月。
1.2 研究方法
1.2.1种子生活力检测
试验前即贮藏当日,随机选取100 粒净种子,采用TTC 图像法测定种子生活力[16],3 次重复。
1.2.2种子预处理
从贮藏之日开始,每2 个月取样1 次。取样前将袋内种子搅拌均匀,然后随机称取足量种子置于500 mL的烧杯中,并用1.5%硫酸铜溶液浸种10 min 进行消毒,取出后用蒸馏水冲洗3 次,并将表面水分吸干备用。
1.2.3种子发芽试验
采用纸上发芽法[17],随机将100 粒种子置于标准发芽床,在人工气候培养箱内进行发芽试验,3 次重复。培养箱内全天光照,温度保持在24~26 ℃。发芽期间,每天定时浇水并观察记录种子的发芽情况。第10 天统计种子的发芽势,第21 天统计种子的发芽率。试验结束后,将幼苗置于105 ℃烘箱中杀青15 min,再降至80 ℃干燥24 h,称其干质量,并计算种子的发芽指数和活力指数[16]。


式中,Dt为发芽时间,d;Gt为与Dt相对应的每天发芽种子数;S为发芽试验结束时正常幼苗单株干质量,g。
1.2.4种子生理指标测定
过氧化物酶(POD)活性采用愈创木酚法[18]、超氧化物歧化酶(SOD)活性采用氮蓝四唑法[18]、过氧化氢酶(CAT)活性采用紫外分光光度法[19]测定。
1.3 数据分析
用Excel 2010 软件统计数据和作图,用SPSS 22.0 软件进行单因素方差(ANOVA)分析,采用Duncan’s 检验法进行多重比较。
2 结果与分析
2.1 种子的生活力和种子萌发随贮藏时间的变化
经TTC (2,3,5-氯化三苯基四氮唑)染色处理后,在立体显微镜下观测到‘莫拉特Ⅱ’杂交臂形草种胚被染成红色的种子占供试种子的64.33%。可见,云南省普洱市生产的‘莫拉特Ⅱ’杂交臂形草净种子的生活力为64.33%。
图1 显示:随贮藏时间的延长,‘莫拉特Ⅱ’杂交臂形草种子的发芽率和发芽势均呈现出逐渐升高的趋势,最后趋于稳定。在种子贮藏12 个月内,种子的发芽率变化不大,与对照(CK)均没有显著差异(P>0.05);贮藏第14 个月,种子的发芽率上升到40.00%,是对照的1.25 倍(P<0.05),说明休眠已经开始释放;贮藏22 个月,种子的发芽率为53.00%,是对照的1.66 倍(P<0.05);贮藏24 个月,发芽率略有上升,达到54.33%,是对照的1.70 倍(P<0.05),但与贮藏22 个月没有差异(P>0.05)。种子的发芽势在贮藏6 个月内变化不大,与对照(CK)均没有显著差异(P>0.05);贮藏8~20 个月,发芽势缓慢上升,较对照均存在显著差异(P<0.05);贮藏第22 个月后,发芽势趋于稳定,至第24 个月,发芽势达到53.00%,是对照的2.27 倍(P<0.05),但与贮藏第22 个月无差异(P>0.05),说明种子早已通过休眠。
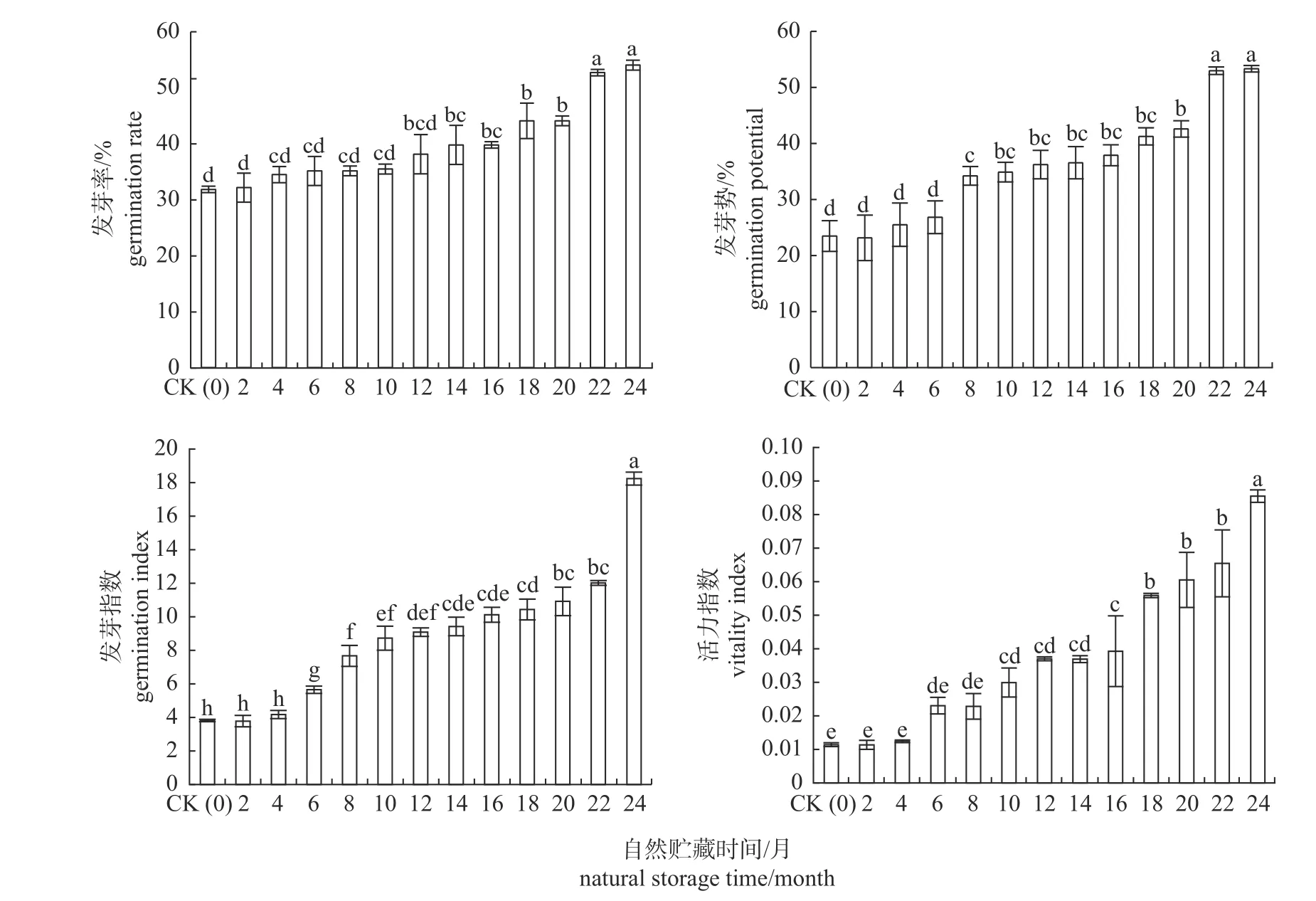
图1 贮藏时间对莫拉特Ⅱ杂交臂形草种子的影响Fig.1 Effect of storage time on ‘Morat Ⅱ’ Brachiaria hybrid seeds germination
从图1 可以看出:‘莫拉特Ⅱ’杂交臂形草种子的活力指数与发芽指数的变化趋势相似,随着贮藏时间的延长,均呈现逐渐升高的趋势。在种子贮藏4 个月内,种子的发芽指数变化不大,较对照(CK)没有差异(P>0.05);贮藏6~22 个月,种子的发芽指数呈现缓慢上升的趋势,较对照均有显著差异(P<0.05);贮藏至第24 个月,其发芽指数达到最高值为18.20,是对照的4.78 倍(P<0.05)。自然贮藏8 个月内,种子的活力指数变化不大,较对照(CK)均没有显著差异(P>0.05);贮藏10~22 个月,种子的活力指数呈现缓慢上升,与对照相比均有显著提高(P<0.05);至贮藏24 个月,种子的活力指数达到最大值,为0.085 3,是对照的7.48 倍(P<0.05)。
2.2 自然贮藏过程中杂交臂形草种子抗氧化酶活性的变化
图2 显示:‘莫拉特Ⅱ’杂交臂形草种子的SOD 活性与POD的变化情况相似,随着贮藏时间的延长均呈现逐渐下降的趋势。种子在收获后贮藏当日即对照(CK)的POD 活性最高,为28.33 U/(g·min) (FW),贮藏2 个月,POD 活性为26.67 U/(g·min) (FW),与对照无显著差异(P>0.05);贮藏4~24 个月,POD 活性缓慢下降,但较对照均有显著降低(P<0.05),至贮藏24 个月,种子酶活性仅为1.67 U/(g·min) (FW),较对照降低94.12% (P<0.05)。种子在收获后贮藏当日即对照(CK)的SOD 活性最高,为4 515.15 U/(g·min)(FW),与贮藏2 个月种子的SOD 活性差异不大(P>0.05);贮藏4~24 个月,SOD的活性缓慢下降,较对照均有显著差异(P<0.05);至贮藏24 个月,其活性降到最低,为107.85 U/(g·min) (FW),较对照降低97.61% (P<0.05)。
图2 显示:与POD 和SOD 活性的变化趋势相反,在自然贮藏条件下,随着贮藏时间的延长,‘莫拉特Ⅱ’杂交臂形草种子的CAT 活性呈现出逐渐升高的趋势。种子贮藏2~12 个月,酶活性增幅不大,较对照(CK)均无显著差异(P>0.05);贮藏14~24 个月,酶活性显著提高,较对照及贮藏前期(2~12 个月)均有显著差异(P<0.05)。贮藏24 个月,种子的CAT 活性最高,为813.66 U/(g·min),较对照显著提高91.29% (P<0.05)。

图2 自然贮藏过程中种子抗氧化酶活性的变化Fig.2 Changes of antioxidant enzymes activity during natural storage
3 讨论
3.1 自然贮藏时间对杂交臂形草种子萌发的影响
发芽率是衡量种子质量好坏的重要指标,反映种子发芽数量的多少;发芽势是反映种子质量优劣的主要指标之一,体现种子发芽的快慢和整齐度;发芽指数反映种子在整个发芽期的综合活力;活力指数既能反映种子发芽率和发芽速度的高低,又能够反映生长势及生长活力的强弱[20]。本试验表明:在自然贮藏过程中,‘莫拉特Ⅱ’杂交臂形草种子的发芽率和各项活力指标均随贮藏时间的延长而逐渐升高,与同为热带牧草和休眠程度同样较深的纳罗克非洲狗尾草种子的情况[21]类似。
‘莫拉特Ⅱ’杂交臂形草净种子的生活力为64.33%,贮藏22 个月时,其种子的发芽率达到53.00%,较贮藏24 个月没有差异(P>0.05),说明此时种子的休眠已经基本释放完毕,发芽率随贮藏时间的延长继续提高的空间越来越小。从另外一个角度来看,贮藏22 个月时种子的发芽率占净种子生活力的82.39%,按照80.00%具有生活力的种子能正常发芽作为通过休眠的标准[16],也说明贮藏22 个月后种子已经通过休眠。因此,可以确定‘莫拉特Ⅱ’杂交臂形草种子的休眠期为22 个月。
3.2 自然贮藏过程中杂交臂形草种子抗氧化酶活性的变化
SOD、POD 和CAT 是细胞内清除活性氧的主要保护酶系统。POD 是植物体内重要的一种抗氧化酶,CAT 是植物体内重要的清除酶,两者协同作用清除植物体内产生的过氧化氢等活性氧[22],增强植物的抗逆性。SOD 可以使有害的超氧阴离子自由基发生歧化作用,形成分子氧和过氧化氢,从而消除自由基对植物造成的伤害[23]。杨艳等[24]在研究珙桐(Davidia involucrateBaill)种子休眠解除与萌发的过程中发现:种子通过各抗氧化酶活性间的相互作用,促进种子萌发。本研究发现:随着贮藏时间的延长,‘莫拉特Ⅱ’杂交臂形草种子POD 和SOD的活性均逐渐降低,这与王秋爽等[25]研究NaCl 胁迫对鸡冠花(Celosia cristataL.)种子的酶活变化相似。韦小丽等[26]研究指出:随着贮藏时间的延长,麻风树种子SOD 活性逐渐下降;钱秀珍等[27]研究贮藏时间对油菜种子生理生化的影响时发现:随着贮藏时间的延长,种子SOD 活性也逐渐下降。说明在贮藏初期杂交臂形草种子的POD 和SOD 酶活性较高,有助于种子休眠解除,为种子休眠释放创造了有利条件;而随着自然贮藏时间的延长杂交臂形草种子的CAT 活性则呈现逐渐升高的趋势,这与罗富成等[28]研究种子休眠完全解除之前CAT 活性的变化趋势一致,金小雯等[29]也证实白燕7 号在4 年的贮藏期内,CAT 活性呈上升趋势。说明种子在自然贮藏过程中,代谢强度逐渐增加,种子休眠逐渐释放。
4 结论
(1)在自然贮藏条件下,随着贮藏时间的延长,‘莫拉特Ⅱ’杂交臂形草种子的发芽率和活力水平逐渐升高,贮藏第14 个月发芽率为40.00%,种子休眠已经开始释放,至贮藏第22 个月,种子休眠已经释放完毕,其休眠期为22 个月。说明‘莫拉特Ⅱ’杂交臂形草种子的休眠释放是一个渐进的过程。
(2)在自然贮藏条件下,随着贮藏时间的延长,‘莫拉特Ⅱ’杂交臂形草种子的POD 和SOD活性逐渐降低,贮藏初期的高活性为种子休眠的自然释放创造了有利条件;CAT 活性则逐渐上升,使种子代谢强度逐渐增加,推动了种子的休眠释放过程。
