基于UPLC-Q-TOF-MS技术对高盐稀态酱油 发酵过程中代谢产物变化的分析
2021-11-05冯云子黄梓堃赵谋明
冯云子,黄梓堃,赵谋明
(华南理工大学食品科学与工程学院,广东 广州 510640)
酱油是东南亚国家常见的大宗发酵调味品,因其特殊的风味逐渐受到世界人民的喜爱。其以大豆或豆粕等植物蛋白、面粉或小麦粉等淀粉质为原料,经微生物接种发酵而制得的一种具有特殊色、香、味的液态调 味品[1]。酱油中含有丰富的初级代谢产物和次级代谢产物,与酱油的风味形成和生理功能密切相关。初级代谢产物包括还原糖、氨基酸、脂肪酸、碳水化合物、有机酸、蛋白质等[1],而次级代谢产物则有生物碱、黄酮类、皂苷、酚酸、苯丙素、硫代葡萄糖苷、多胺等[2]。
代谢组学是研究样品中所有的低分子质量代谢产物的一种技术,该研究方法比单一指标的分析方法更加系统,目前广泛应用于药物学、环境监测、微生物等领域[3]。 近年来,代谢组学技术在传统发酵食品的研究领域应用越来越多,有助于解析发酵过程复杂的生物化学变化,揭示发酵机理[4],涉及的检测手段主要包括核磁共振(nuclear magnetic resonance,NMR)技术、离子迁移光谱(ion mobility spectroscopy,IMS)、气相色谱-质谱(gas chromatography-mass spectrometry,GC-MS)联用技术、液相色谱-质谱(liquid chromatography-mass spectrometry,LC-MS)联用等。Wu Xiaohe等[5]采用1H-NMR技术对清香型、浓香型和酱香型的白酒大曲进行非靶标代谢组学分析,分别鉴定出苹果酸、甘露醇、异亮氨酸等代表各自类型大曲的生物标志物;袁志鹰等[6]采用GC-IMS技术对发酵前后百合发酵乳的风味物质进行系统分析,找出15 种具有明显差异的挥发性成分;Feng Yunzi等[7]采用顶空固相微萃取和气相色谱-嗅觉测定技术检测出酱油中2-甲基丁醛、3-甲硫基-丙醛等25 种 挥发性化合物;Lee等[8]采用气相色谱-质谱联用和LC-MS联用技术对豆酱制作工艺进行分析,发现曲霉菌、芽孢杆菌、四联球菌和结合酵母分别与糖、脂肪酸和氨基酸代谢有关。
超高效液相色谱(ultra-high performance liquid chromatography,UPLC)与高分辨度四极杆飞行时间质谱(quadrupole-time-of-flight-mass spectrometry,Q-TOFMS)技术联用,近年来在代谢组学研究中的使用频率越来越高,相对于普通LC-MS技术具有更高的分辨度和灵敏度,对样品分析效率和准确性大大提高[9]。如Cheng Lizeng等[10]采用UPLC-Q-TOF-MS对青砖茶发酵前后的代谢图谱进行分析,共检测出102 种关键代谢产物,证明了微生物发酵是改变生茶叶滋味的关键过程。Lee等[11]采用GC-TOF-MS和UPLC-Q-TOF-MS系统分析了豆酱整体发酵过程的代谢途径及其产生的代谢产物,结合多元数据分析,将豆酱的发酵过程分为5 个阶段。然而,目前鲜有采用UPLC-Q-TOF-MS技术对酱油发酵过程中代谢物质变化进行研究的相关报道。
因此,本实验对高盐稀态酱油发酵过程进行对比分析,基于UPLC-Q-TOF-MS技术,结合主成分分析(principal component analysis,PCA)、正交偏最小二乘法回归分析(orthogonal partial least-squares discrimination analysis,OPLS-DA)等多元数据分析方法研究酱油发酵过程中代谢产物,明确酱油发酵阶段间具有显著性差异的物质,并确定其在酱油发酵过程中的变化趋势,旨在为改进酱油发酵工艺提供理论基础及支撑。
1 材料与方法
1.1 材料与试剂
酱油采用高盐稀态发酵工艺制作,分别于酱醪发酵第1、5、15、30、60、90天取样,样品贮存于密封的酱油瓶中,并贮藏于-20 ℃冰柜待用。
乙醚(分析纯,纯度≥99.5%) 国药集团化学试剂有限公司;甲醇、甲酸、乙腈(均为色谱纯) 西格玛奥德里奇(上海)贸易有限公司。
1.2 仪器与设备
EL204/EL3002电子天平 梅特勒-托利多仪器有限公司;Sorvall ST 16R台式离心机 赛默飞世尔科技(中国)有限公司;RE-52AA旋转蒸发仪 上海 亚荣生化仪器厂;SHZ-D循环水式真空泵 巩义予华仪器有限公司;MTN-2800W氮吹浓缩装置 天津奥特赛恩斯仪器有限公司;ACQUITY UPLC高效液相色谱仪、CORTECS UPLC C18色谱柱(2.1 mm×100 mm,1.6 μm) 沃特世科技(上海)有限公司;MicrOTOF-Q II高分辨串联质谱仪 布鲁克(北京)科技有限公司;Milli-Q超纯水机 美国密理博公司;WH-3微型旋涡混合仪 上海泸西分析仪器厂有限公司。
1.3 方法
1.3.1 酱油样品前处理
参考李会品[12]研究,并基于前期预实验结论优化得到如下方法:取20 mL酱油于50 mL离心管中,加入15 mL乙醚后涡旋振荡3 min,4 ℃、10 000 r/min离心15 min,用胶头滴管移取有机相于另一离心管中。再加入15 mL乙醚于原离心管,重复上述操作,合并有机相。40 ℃水浴下,氮吹至干,用1 mL 80%甲醇溶液复溶,过0.22 μm滤膜备用。
1.3.2 色谱条件
参考Seo[13]和Chang Tesheng[14]等的方法并进行优化改进。色谱柱采用CORTECS UPLC C18色谱柱(2.1 mm×100 mm,1.6 μm),采用0.1%甲酸(A)和乙腈(B)作为流动相进行梯度洗脱,流速为 0.2 mL/min。梯度洗脱程序为:0~1 min,85% A、15% B;1~12 min,85%~20% A、15%~80% B;12~22 min,20%~0% A、80%~100% B;22~26 min,0% A、100% B;26~28 min,0%~85% A、100%~15% B;28~30 min,85% A、15% B。柱温35 ℃,进样量2 μL。
1.3.3 质谱条件
质谱扫描模式为正离子模式,扫描范围m/z50~2 000,电子电离源能量10.0 eV,毛细管电压3.5 kV,离子源加热温度200 ℃,干燥气体流量 4.0 L/min,雾化器压力0.5 bar。
1.4 数据分析
流动相条件设定及控制采用Hyphenation Star(version 3.2,Bruker Daltoik GmbH,德国)软件控制,质谱数据信息用高分辨度飞行时间质谱软件Otofcontrol(version 4.0,Bruker Impact II,德国)采集,基峰图、总离子流图和紫外信息采用Bruker Compass Data Analysis软件(version 4.4,Bruker Daltoik GmbH,德国)进行分析。质谱信息定性参考数据库包括人类代谢组数据库HMDB 4.0(http://www.hmdb.ca)、METLIN数据库(http://metlin.scripps.edu)和MassBank数据库(http://www.massbank.jp)。数据分析采用Microsoft Excel 2019(微软公司,美国)。柱状图绘制采用OriginPro(version 2018C,OriginLab公司,美国),多元数据分析采用SIMCA(version 14.1,MKS Umetrics,瑞典)完成。
2 结果与分析
2.1 酱油发酵过程样品提取液的UPLC-Q-TOF-MS进样情况
采用UPLC-Q-TOF-MS技术,在高盐稀态酱油发酵过程中检出的所有代谢物质的数量及总峰面积如表1所示。酱油1 d中检出156 种物质,随着发酵的进行,酱油中的物质种类及总峰面积不断增加,第1个月增长较快,后期变化减缓;到发酵终点(90 d)检出物质共228 种,总峰面积达到2.8×108,物质种类及总峰面积分别为第1天样品的1.47 倍和6.57 倍。
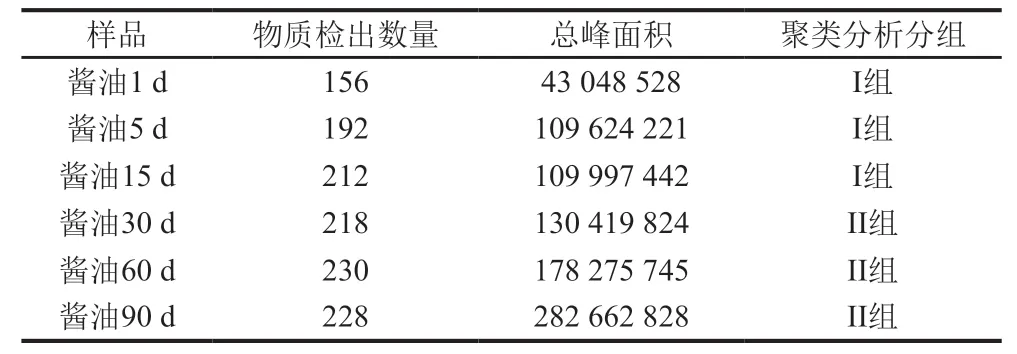
表1 酱油发酵过程中各样品提取液代谢物检出数量和总峰面积Table 1 Number and total peak area of metabolites detected in soy sauce samples during the fermentation process
图1统计了酱油发酵过程中所有样品提取液里被检出的物质种类及数量,共鉴定化合物155 种,种类最丰富的是氨基酸衍生物、脂肪酸和有机酸,分别为31、28 种和21 种。此外,还共检出13 种酯类、9 种酰胺类、8 种醇类、7 种氨基酸、7 种多肽、5 种异黄酮、3 种大豆甾醇、2 种生物碱等。
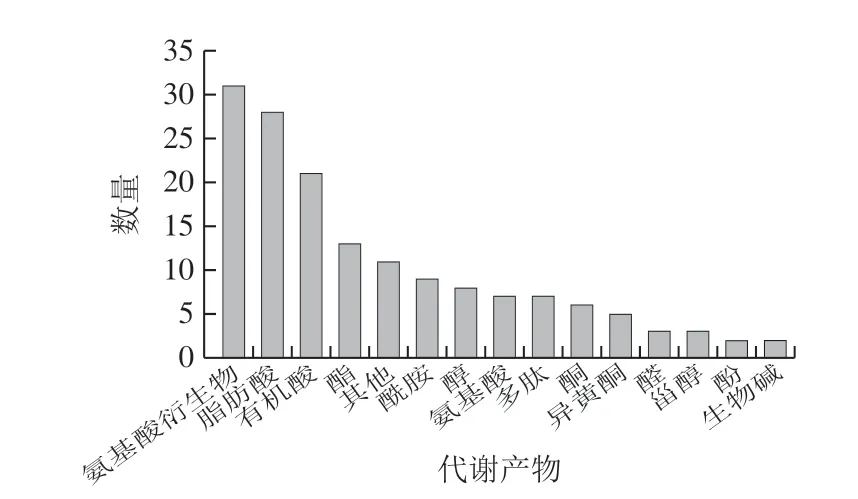
图1 酱油发酵过程样品提取液被检出的代谢产物种类Fig. 1 Change in composition of metabolites detected in soy sauce samples during the fermentation process
2.2 酱油发酵过程中样品提取液的差异代谢物质
基于各样品提取液中所有代谢物的聚类分析结果(表1),发酵过程中的样品可分为两组,第1、5、15天的酱油样品为组I(发酵初期),发酵30、60、90 d的酱油样品为组II(发酵后期),采用OPLS-DA对两组样品进行差异分析,认为变量投影重要性(variable importance projection,VIP)值大于1即为显著差异代谢物。由表2可知,发酵阶段间呈现显著性差异的代谢物质共39 种,其中已知化合物34 种。可根据结构分为7 大类,即氨基酸衍生物类(10 种)、脂肪酸类(9 种)、异黄酮类(5 种)、酯类(3 种)、有机酸类(2 种)、生物碱(2 种)和其他类(3 种)化合物,大部分代谢物含量随着发酵的进行逐渐增加。
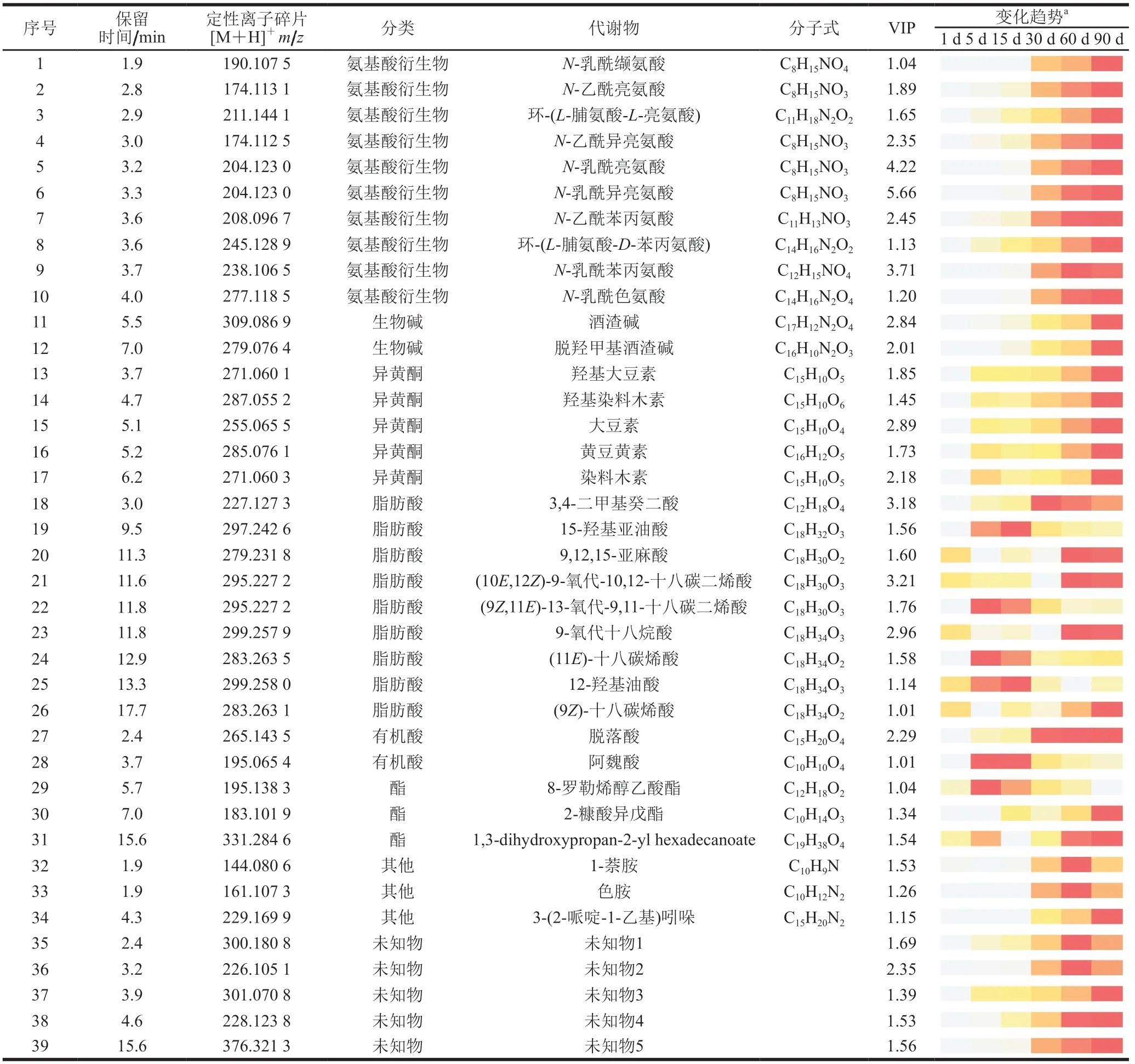
表2 酱油发酵过程样品提取液间的差异代谢物质(VIP>1)Table 2 Differential metabolites (VIP > 1) among soy sauce samples during the fermentation process
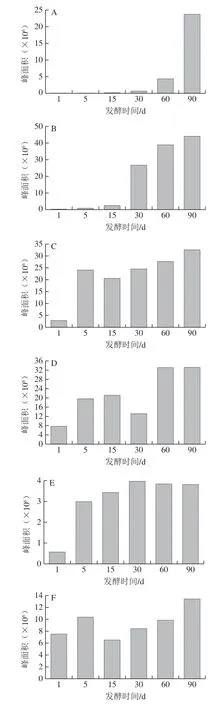
差异代谢物质的总峰面积和大类总峰面积变化趋势如图2所示。除有机酸外,各类物质的峰面积在发酵过程中上升。生物碱、氨基酸衍生物的峰面积在30 d后上升明显,异黄酮、脂肪酸和酯类的峰面积呈波动上升。有机酸类的峰面积在1~5 d变化较大(5.26 倍),并于第30天达到峰值,之后稍有下降。李彦[15]对发酵过程中的总酸含量进行测定,发现其在1~20 d间含量迅速增加,之后趋于平稳,其变化趋势与本模型相似。整体而言,差异代谢物的总峰面积变化与所有化合物总峰面积变化趋势基本一致,变化较大的阶段为1~5 d(3.10 倍)和30~60 d(1.56 倍)。除此之外,发酵第1天的样品中氨基酸衍生物、异黄酮和生物碱的峰面积占总峰面积的0.44%、6.82%和0.02%,到第90天,其分别升至20.11%、14.88%和10.81%,是酱油中的关键差异代谢产物。
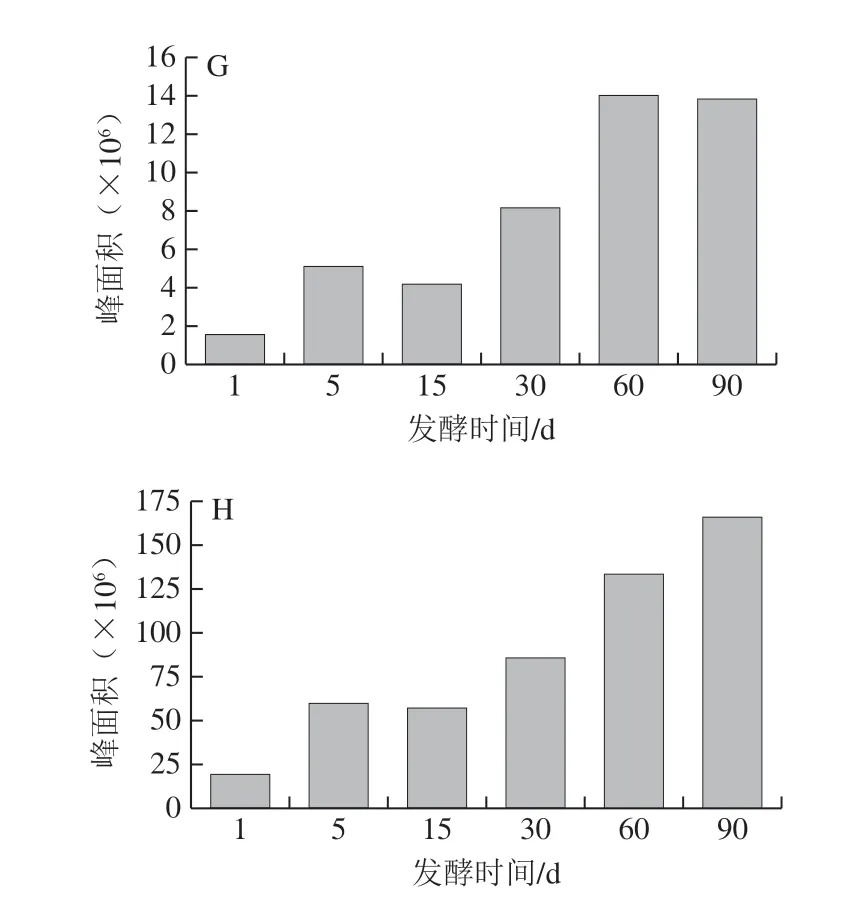
图2 各类差异代谢物在酱油发酵过程中的总峰面积变化Fig. 2 Changes in total peak area of each class of metabolites during soy sauce fermentation
由图3A所示,PC1和PC2的贡献率分别为70.2%和16.2%,表明模型具有较好的代表性。得分图(图3A)中,各样品点分散情况较好,说明不同发酵时间的酱油物质种类和含量均有差异。而载荷图(图3B)反映了酱油中的差异代谢物对PCA样本分类的贡献程度。整体上看,可以将酱油阶段分成发酵前期和发酵后期,随着发酵时间的延长,发酵过程具有从PC1负半轴往PC1正半轴演变的趋势。针对各样品,酱油1 d在第3象限,在对应的载荷图中没有点落于此,说明酱油1 d各类差异物质含量较低。酱油5 d、15 d在第2象限,载荷图中第2象限有6 个物质,主要包括4 种脂肪酸(9-氧代十八烷酸、(11E)-十八碳烯酸、15-羟基亚油酸、12-羟基油酸)、 8-罗勒烯醇乙酸酯和阿魏酸。这些物质在发酵过程中的变化趋势是先增后减,其峰面积于第5天或第15天达到峰值(表2)。酱油30 d、60 d和90 d均在1、4象限的分界线两侧,载荷图中则显示共有33 种物质都落于PC1正半轴上。说明随着发酵的进行,酱油发酵过程样品提取液间差异代谢物质主要集中于发酵后期(30 d之后)。
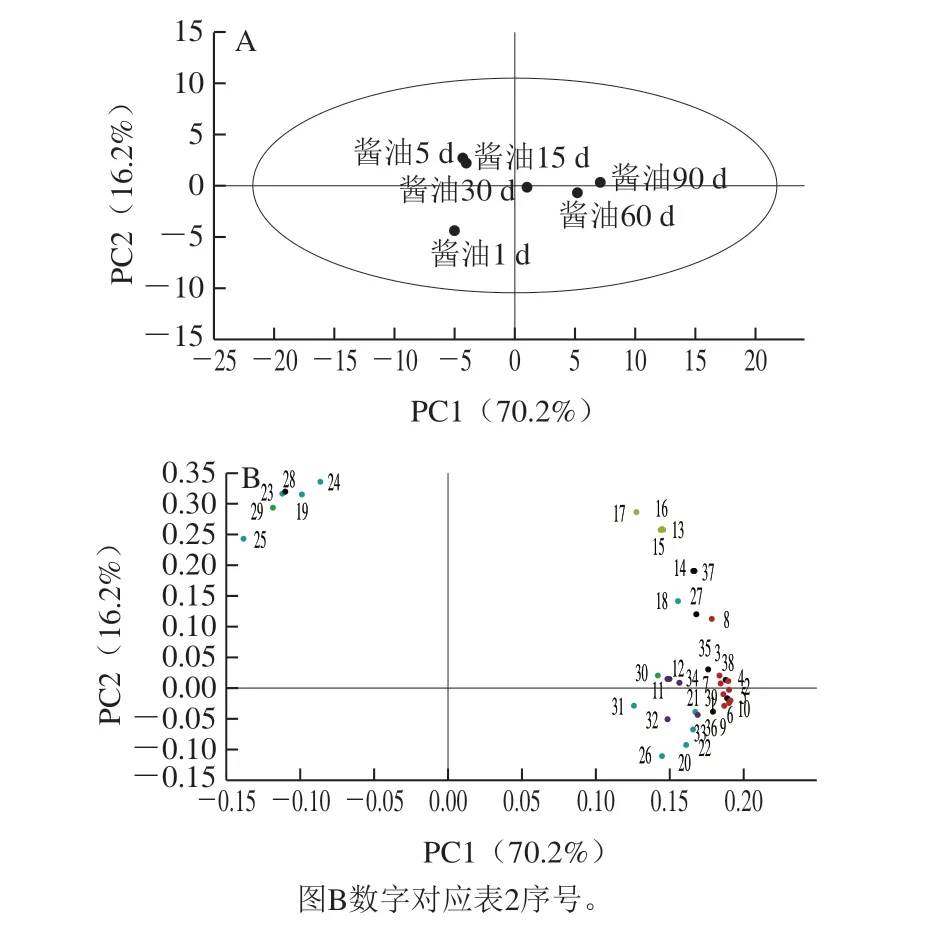
图3 酱油发酵过程样品提取液的差异代谢物质的PCA 得分图(A)和载荷图(B)Fig. 3 PCA score plot (A) and loading plot (B) of differential metabolites in soy sauce samples during the fermentation process
对于位于PC1正半轴的物质,5 种异黄酮落在 第1象限,即大豆素、黄豆黄素、染料木素、羟基大豆素、羟基染料木素。随着发酵的进行,异黄酮峰面积不断增加,在第90天达到峰值(图2)。异黄酮是具有高生物活性的一类物质,其具有抗氧化、抗肿瘤、增强免疫力等功能[16-17],是酱油中常见的次级代谢产物。Chang Tesheng等[14]曾在酱油中鉴定出上述异黄酮,并证明了大豆素和染料木素分别是羟基大豆素和羟基染料木素的前体物质。另外,脱落酸和3,4-二甲基癸二酸也落在第1象限,其峰面积在前30 d的增长较快,之后趋于稳定,在第90天达到峰值(表2)。
第1、4象限分界线中的物质点十分密集,包括9 种氨基酸衍生物。它们的峰面积在发酵过程中不断增加,在1~5 d和15~30 d变化较大(5.13 倍和10.62 倍),并第90天达到峰值(图2)。其中N-乳酰异亮氨酸、N-乳酰亮氨酸、N-乳酰苯丙氨酸的VIP值为差异代谢物前3,分别为5.66、4.22和3.71,表明其增长最为明显。含有乳酰基的氨基酸衍生物在帕尔马干酪(用脱脂乳制成的意大利干酪)和日式酱油中均有被发现[18-19],其中乳酰谷氨酸已经被证实对奶酪的鲜味具有贡献[19-20]。乳酰氨基酸的生成与乳酰转换酶有关,推测其含量与乳酸菌的作用可能相关[20],且乳酰氨基酸的生成可能对酱油发酵后期浓厚味的不断增强具有一定贡献。此外,还包括系列乙酰基氨基酸和环状二肽,如N-乙酰异亮氨酸、N-乙酰亮氨酸、环-(L-脯氨酸-D-苯丙氨酸)和环-(L-脯氨酸-L-亮氨酸)。有研究表明在大豆种子和日式酱油中曾检测到乙酰基谷氨酸[19,21]。对于环状二肽,有文献曾在烟曲霉培养液和链球菌培养液中鉴定出环-(脯氨酸-亮氨酸)和环-(脯氨酸-苯丙氨酸)[22],李会品[12]以XOD抑制活性为导向,在乙酸乙酯提取得到的酱油有机相中分离得到了另一种二肽,即环-(L-脯氨酸-L-丙氨酸),是酱油中的活性物质。但有研究表明,含L-脯氨酸的环肽是N端含有L-脯氨酸-L-X序列的肽在酶的作用下形成的,这些含L-脯氨酸的环肽可能主要呈苦味[23]。
酒渣碱及脱羟甲基酒渣碱也落于PC1正半轴周围。其峰面积在发酵前中期较低,从15 d开始变化差异显著(5.41~7.04 倍),第90天达到峰值(图2)。酒渣碱已被证实是酱油中的活性物质,其对于酱油的抗氧化活性具有卓越的贡献[12],并能抑制黄嘌呤氧化酶活性,能降低高尿酸血症小鼠的血清尿酸水平[24]。脱羟甲基酒渣碱也属于骆驼蓬生物碱的一种,在水果和黑醋栗中曾被检出[25]。此外,4 种脂肪酸在第4象限((10E,12Z)-9-氧代-10,12-十八碳二烯酸、(9Z,11E)-13-氧代-9,11-十八碳二烯酸,9,12,15-亚麻酸、(9Z)-十八碳烯酸)。其中(9Z)-十八碳烯酸与(11E)-十八碳烯酸互为顺反异构体。
其他类物质中,如色胺、萘胺等也落于第4象限。于金芝等[26]研究有发现在高盐稀态酱油生产过程中,色胺含量随发酵的进行先升后降,在第60天达到最大,其变化趋势与本研究结果一致。
3 结 论
酱油发酵过程中的样品共被鉴定出155 种代谢物质,其总峰面积在发酵1~5 d和30~60 d变化幅度较大,并于发酵末期(60~90 d)达到峰值。通过多元数据分析鉴定出34 种已知差异代谢产物(VIP>1)。氨基酸衍生物、异黄酮及生物碱等物质在酱油发酵过程中峰面积变化较大,均随发酵过程的进行而升高,并于发酵第90天达到峰值,分别占总峰面积的20.11%、14.88%和10.81%。而脂肪酸和酯类分别从18.16%和17.72%下降至15.17%和6.13%。另外,VIP值前3的物质分别为N-乳酰异亮氨酸、N-乳酰亮氨酸、N-乳酰苯丙氨酸,这些代谢物均为含乳酰基的氨基酸衍生物。差异代谢物质的明确为酱油产品的调控及开发提供了重要理论基础和方向,具有显著差异的代谢物质对风味及活性贡献值得进一步的研究探讨。
