塔罗科血橙室温贮藏期间花色苷和糖酸积累变化及相关代谢基因表达特征
2021-11-02洪敏贺明阳王日葵周炼王晶冯雨
洪敏,贺明阳,3*,王日葵,周炼,王晶,冯雨
(1.西南大学柑桔研究所,重庆 400712;2.中国农业科学院柑桔研究所,重庆 400712;3.中国科学院华南植物园植物资源保护与可持续利用重点实验室,广州 510650)
血橙[Citrus sinensis(L.)Osbeck]属甜橙类,常见的血橙品种有塔罗科(Tarocco)、桑吉耐洛(Sanguinello)和摩洛(Moro)[1-2]。血橙是唯一含花色苷的柑橘,其果皮和果肉呈现独特的红色是由于花青素与糖结合形成了苷类化合物[3-4]。花色苷属黄酮类化合物[5],是血橙中主要的抗氧化活性成分,能够清除自由基[6],在治疗和预防心脏病、癌症、动脉硬化和抗病毒活性方面也具有重要意义[7],其含量是评价血橙商业价值的重要指标[5]。
花色苷生物合成受温度、糖、酸等的影响。PANNITTERI 等[2]、CRIFÒ 等[8]和RAPISARDA 等[9]研究发现,4~10 ℃低温贮藏可促进血橙花色苷积累,延长血橙货架期,且CRIFÒ等[8]证实了花色苷生物合成与冷调控有关。可溶性糖和有机酸是评价柑橘果实风味和整体品质的重要指标[10],也是调控花色苷合成的重要因素。前人研究发现,经可溶性糖处理后,拟南芥[11-12]、西兰花[13]花色苷含量积累增加,尤其以蔗糖处理的效果最好[11],它可以特异调节糖依赖的花色苷合成上调[10]。LO PIERO等[14]发现,经低温贮藏后血橙柠檬酸含量降低,柠檬酸裂解酶活性增强,而柠檬酸裂解后的反应产物可促进黄酮的生物合成,该过程因冷藏而被激活。喻最新等[15]发现,草酸处理可提高血橙采后花色苷和可溶性糖含量,改善果实贮藏品质。但有机酸调控花色苷合成的研究较少,而对花色苷的辅色和稳态性研究较多[16-17]。目前,对血橙花色苷调控研究主要集中在低温,而在室温条件下血橙花色苷和糖酸含量变化及其相关代谢基因表达水平尚不清楚。
柑橘花色苷[18-19]、糖[20-22]、酸[23-24]代谢途径已明确。莽草酸途径通过连接糖代谢与次生代谢生成苯丙氨酸,开启花色苷代谢途径[25]。苯丙氨酸经苯丙氨酸解氨酶(phenylalanine ammonialyase, PAL)、肉桂酸4-羟化酶(cinnamate 4-hydroxylase, C4H)、4-香豆酸∶辅酶A连接酶(4-coumarate∶coenzyme A ligase, 4CL)、查耳酮合成酶(chalcone synthase,CHS)、查耳酮异构酶(chalcone isomerase,CHI)、黄酮醇3-羟化酶(flavonone 3-hydroxylase, F3H)、类黄酮3´-羟化酶(flavonoid 3´-hydroxylase, F3´H)、类黄酮3´,5´-羟化酶(flavonoid 3´, 5´-hydroxylase,F3´5´H)、二氢黄酮醇4-还原酶(dihydroflavonol 4-reductase, DFR)、花青素合成酶(anthocyanin synthase, ANS)、糖基转移酶(UDP-glucoseflavonoid 3-O-glucosyltransferase, UFGT)、谷胱甘肽硫转移酶(glutathione-S-transferases,GST)等催化合成花色苷[18-19]。黄酮醇合成酶(flavonol synthase,FLS)负调节花色苷合成[18]。柑橘是蔗糖积累型果实,蔗糖代谢是糖积累的重要过程,蔗糖合成酶(sucrose synthase, SS)可逆调控其合成与分解[21],并受蔗糖磷酸合成酶(sucrose phosphate synthase,SPS)的合成调控及酸性转化酶(acid invertase,AI)的分解调控,且催化反应不可逆[26]。柠檬酸是柑橘特有的有机酸,乙酰辅酶A与草酰乙酸在柠檬酸合成酶(citrate synthase,CS)的催化下合成柠檬酸[23],柠檬酸的分解主要由乌头酸酶(aconitase,AC)、异柠檬酸脱氢酶(isocitrate dehydrogenase, IDH)等催化[24]。
塔罗科血橙属晚熟柑橘[2],采收时温度偏高,果实采后呼吸作用比较强烈,且随着采收期推迟,其耐贮性降低,严重影响物流过程中血橙的商品性[27-28]。由于我国冷链物流体系尚不完善,所以本研究选用高效液相色谱(high performance liquid chromatography,HPLC)和实时荧光定量聚合酶链反应(polymerase chain reaction,PCR)技术分析室温条件下血橙采后花色苷和糖酸含量变化及相关代谢基因表达特征,以期为血橙物流和货架期品质保持研究提供理论依据。
1 材料与方法
1.1 材料与处理
本研究选用种植于重庆市璧山区绿跃血橙种植园的塔罗科血橙为试验材料。采收大小和色泽均匀、无机械损伤和病虫害、九成熟的果实,用配置的500 mg/L 咪鲜胺浸泡30 s,捞出后自然风干。将处理后的果实置于(20±5)℃室温条件下贮藏20 d。每次试验重复3 次,每次重复取10 个果实,每隔5 d测定一次相关指标。每次试验采集血橙果肉样品,经液氮速冻后于-80 ℃冰箱中保存,备用。
1.2 仪器与设备
TU-1901 紫外分光光度计,北京普析通用仪器有限公司;RE52CS-2旋转蒸发器,上海亚荣生化仪器厂;高效液相色谱(HPLC)仪,美国安捷伦公司;H1850R 台式高速冷冻离心机,长沙湘仪离心机仪器有限公司;CFX96TM实时荧光定量PCR仪,美国伯乐公司。
1.3 方法
1.3.1 总花色苷含量的测定
血橙果肉总花色苷含量的测定参考曹少谦等[3]的pH示差法。每个样品重复3次。
1.3.2 有机酸和可溶性糖的提取与测定
有机酸和可溶性糖的提取参考ZHANG 等[29]的方法。称取3 g 研磨后的血橙果肉,加入5 mL 80%冰乙醇,35 ℃水浴浸提20 min 后,在室温、5 000 r/min 条件下离心5 min,然后取上清液,重复抽提3次,合并上清液并定容至20 mL。取5 mL提取液,在室温、5 000 r/min条件下离心5 min,取3 mL上清液在35 ℃条件下旋转蒸干,然后用1.5 mL 双蒸水溶解,最后用直径13 mm、孔径0.22 μm 的水膜过滤纯化。提取的滤液用于测定可溶性糖和有机酸含量。HPLC 色谱条件参考喻最新等[5]的方法。每个样品重复3次。
总糖为蔗糖、葡萄糖、果糖含量之和,总酸为柠檬酸、苹果酸、抗坏血酸含量之和。
1.3.3 血橙果肉总RNA 提取及cDNA 合成
血橙果肉总RNA 提取参照天根DP441 型试剂盒的操作说明,然后参照宝生物RR047A 型试剂盒操作说明反转录成cDNA,-20 ℃保存,备用。
1.3.4 表达分析
血橙内参基因选用EF-1α[8](GenBank登录号为XM_015533332),花色苷及糖酸代谢相关基因引物序列分别参考CARMONA 等[18]和LIU 等[30]的研究。参照宝生物SYBR®Premix ExTaqTMⅡ试剂盒的操作说明,在实时荧光定量PCR 仪上进行扩增反应。反应体系如下:10 μL SYBR Green Ⅰ混合物,正向和反向引物各0.8 μL,1.5 μL cDNA 模板,6.9 μL ddH2O。扩增程序如下:95 ℃预变性30 s;95 ℃变性5 s,58 ℃退火30 s,40 个循环。熔解曲线绘制:95 ℃变性10 s,降温到65 ℃后开始以0.5 ℃/s 逐步升温,每5 s 采集一次荧光信号,反应至95 ℃时结束。每个样品重复3 次。采用2-ΔΔCT法进行相对定量分析。
1.3.5 数据统计与分析
采用Excel 2007进行数据统计,采用SPSS 20.0进行差异显著性分析和皮尔逊相关性分析。
2 结果与分析
2.1 室温贮藏期间血橙总花色苷含量变化
如图1所示,室温贮藏期间果肉总花色苷含量呈上升趋势。贮藏至第5 天,果实花色苷含量从2.86 mg/L 显著增加至8.26 mg/L,之后果实花色苷含量缓慢增加,到第20 天时其含量达到最大,为9.21 mg/L。整个贮藏期间花色苷含量提高了222%。

图1 室温贮藏期间血橙总花色苷含量的变化Fig.1 Changes of total anthocyanin content in blood oranges during room temperature storage
2.2 室温贮藏期间血橙花色苷代谢相关基因表达水平变化
如图2所示:PAL、4CL、CHS、CHI、F3H、F3´5´H、DFR、ANS、UFGT、GST表达水平基本呈上升趋势;FLS表达水平呈下调趋势;C4H表达水平呈波动变化。前人在血橙低温贮藏过程中也发现类似结果[15]。相关性分析发现:血橙花色苷含量与PAL、4CL、CHS、CHI、F3H、F3´5´H、DFR、ANS、UFGT、GST表达水平呈正相关,且与4CL、CHI、ANS的相关性显著,相关系数分别为0.691、0.683、0.662;与C4H、FLS表达水平呈负相关,但相关性不显著。

图2 室温贮藏期间血橙花色苷代谢相关基因表达水平的变化Fig.2 Changes of expression levels of anthocyanin metabolism related genes in blood oranges during room temperature storage
2.3 室温贮藏期间血橙可溶性糖和有机酸含量变化
如图3 所示:贮藏0~15 d,果实总糖、蔗糖、果糖和葡萄糖含量均显著增加至最大值,分别为76.28、36.43、20.55、19.29 mg/g,分别提高了16.4%、11.9%、23.8%、17.7%。贮藏15~20 d,果实总糖、蔗糖、果糖和葡萄糖含量均显著降低,分别下降了3.8%、3.8%、3.3%、4.3%。整个贮藏期间,果实总糖、蔗糖、果糖和葡萄糖含量分别提高了12.0%、7.7%、19.7%、12.7%。柑橘中柠檬酸是主要的有机酸,含量明显高于微量积累的苹果酸和抗坏血酸。总酸、柠檬酸和苹果酸含量呈显著波动变化。抗坏血酸含量从0.61 mg/g 显著降低到0.41 mg/g,随后显著增加至0.50 mg/g,这与前人在‘爱媛38号’中的研究结果[31]类似。
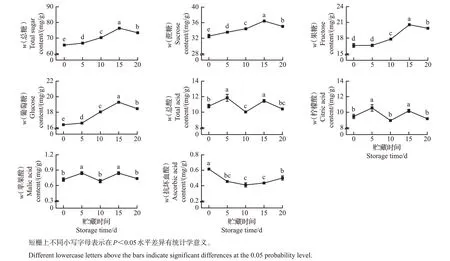
图3 室温贮藏期间血橙可溶性糖和有机酸含量的变化Fig.3 Changes of soluble sugar and organic acid contents in blood oranges during room temperature storage
2.4 室温贮藏期间血橙糖、酸代谢相关基因表达水平变化
如图4所示:SS1、SS2表达水平呈先下调后上调趋势;SPS1、SPS2、SPS3、AI表达水平呈下调趋势。相关性分析发现:蔗糖含量与SS1表达水平呈显著正相关,相关系数为0.547,与SS2无显著相关;与SPS1、SPS2表达水平呈显著负相关,相关系数分别为-0.591、-0.573;与SPS3、AI表达水平呈极显著负相关,相关系数分别为-0.893、-0.913。
由图4可见:CS表达水平先显著下降后显著上升;AC表达水平先显著下调后趋于稳定;IDH表达水平呈波动变化。相关性分析发现:柠檬酸含量与IDH表达水平呈极显著负相关,相关系数为-0.661;与CS、AC表达水平呈负相关,但相关性不显著。

图4 室温贮藏期间血橙糖、酸代谢相关基因表达水平的变化Fig.4 Changes of expression levels of sugar and acid metabolism related genes in blood oranges during room temperature storage
2.5 室温贮藏期间血橙花色苷积累与糖酸含量的相关性分析
由表1 可见:室温贮藏期间血橙花色苷含量与总糖、蔗糖、果糖和葡萄糖含量呈显著正相关,相关系数分别为0.688、0.748、0.606、0.687,其中与蔗糖的相关性最高;与抗坏血酸含量呈极显著负相关,相关系数为-0.852。

表1 皮尔逊相关系数Table 1 Pearson correlation coefficients
3 讨论
3.1 室温贮藏期间血橙可溶性糖积累及表达分析
本研究结果表明,血橙果实中主要积累蔗糖、果糖和葡萄糖等可溶性糖,在整个贮藏期间果实中蔗糖、果糖和葡萄糖含量比例约为2∶1∶1;龚荣高等[21]在甜橙果实膨大后期也发现类似结果。室温贮藏期间果实果糖和葡萄糖含量变化高于蔗糖,表明蔗糖在合成的同时也在分解[26]。此外,果实中可溶性糖含量呈先显著上升后显著下降的趋势。产生该趋势的原因可能是柑橘中酸类物质向糖类转换[32]或是果实衰老进程中细胞壁成分分解使糖积累暂时增加[15]。而贮藏后期糖积累下调可能是蔗糖降解为葡萄糖和果糖后进入糖酵解途径被消耗或是果肉将能量物质运输给果皮所致[32-33]。相关性分析发现,蔗糖含量与SS1表达水平呈显著正相关,与SS2无显著相关性,与SPS1、SPS2、SPS3呈显著负相关,认为蔗糖合成主要由SS1调控,而SPS在调控蔗糖合成方面不起主导作用。蔗糖含量与AI表达水平呈极显著负相关,表明蔗糖的分解主要由AI调控;赵智中等[20]在温州蜜柑中也发现类似结果。综合分析认为,血橙在室温贮藏期间主要由SS1、AI来调控蔗糖的合成与分解。刘永忠等[34]也发现,蔗糖合成酶(SS)和转化酶在柑橘汁胞糖代谢中起重要作用。
3.2 室温贮藏期间血橙有机酸积累及表达分析
血橙在短期室温贮藏期间总酸、柠檬酸和苹果酸含量呈显著波动变化,表明果实中主要有机酸一直在合成且不断被分解。相关性分析发现,柠檬酸含量与IDH表达水平呈极显著负相关,与AC相关性不显著,表明柠檬酸的分解主要由IDH发挥作用。前人研究发现,椪柑经热处理后柠檬酸含量降低,该过程中IDH表达量也显著升高[23]。本研究中柠檬酸含量与CS表达水平无显著相关,这与CHEN等的研究结果[24]一致。HUSSAIN等[35]也发现,柠檬酸合成酶(CS)活性提高也不会使柠檬酸积累增加。但也有抑制CS 活性后柠檬酸含量降低的报道[36]。目前,有机酸的降解产物主要是作为呼吸作用的底物参与三羧酸循环,从而为细胞供能,此外,它还是初生代谢物(如氨基酸和脂肪酸)和次生代谢物(与风味和香气有关)的重要前体物质[22,35],而对其合成机制还需进一步研究。
3.3 室温贮藏期间血橙花色苷积累及与糖酸含量的相关性分析
血橙在短期室温贮藏期间花色苷含量与4CL、CHI、ANS表达水平呈显著正相关,认为4CL、CHI、ANS是花色苷合成的关键结构基因。相关性分析发现,花色苷与总糖、蔗糖、果糖和葡萄糖含量呈显著正相关。前人在血橙[5]、黑穗醋栗[37]、杨梅[38]果实生长发育过程中也发现,果实花色苷积累与可溶性糖含量显著相关,糖为花色苷积累提供物质基础。糖作为花色苷合成的原料,通过糖代谢途径和信号机制影响花色苷合成[19]。本研究发现,花色苷与抗坏血酸含量呈极显著负相关,但抗坏血酸积累微量,其调控花色苷合成的关系还需进一步验证。本研究中,贮藏0~5 d血橙花色苷含量快速上升,之后缓慢上升。前人研究表明,糖酵解产生的磷酸烯醇式丙酮酸可通过莽草酸途径生成花色苷的直接前体物质苯丙氨酸[25]。贮藏初期花色苷含量急剧增加可能是可溶性糖含量显著增加为花色苷合成提供了原料所致。而在贮藏过程中由于果实间空间小、氧气稀薄,易进行无氧呼吸,糖酵解产生的丙酮酸作为底物开启乙醇代谢[28,39],相对抑制了花色苷合成,致使花色苷积累缓慢。王晶等[31]发现,低温贮藏期间‘爱媛38号’柑橘果实乙醇、乙醛含量整体呈上升趋势。后续可从血橙采后室温贮藏过程中乙醇、乙醛等异味物质积累及乙醇代谢途径方面进一步研究。
4 结论
短期内室温贮藏塔罗科血橙促进了花色苷和可溶性糖的积累,而柠檬酸含量呈波动变化,处于不断合成与分解状态。4CL、CHI、ANS、SS1、AI、IDH在花色苷和糖酸代谢过程中发挥重要作用。室温贮藏期间花色苷含量与总糖、蔗糖、果糖和葡萄糖含量呈显著正相关,认为可溶性糖含量增加是促进血橙花色苷积累的重要因素。本研究结果阐释了血橙在室温贮藏期间花色苷和糖酸积累变化及相关代谢基因表达特征,可为血橙物流和货架期品质保持研究提供理论依据。
