草地贪夜蛾鱼尼丁受体与双酰胺类杀虫剂的结合模式及抗性风险分析
2021-10-27王凯博陶丽红李佳俊吴文伟
李 康, 王凯博, 陶丽红, 李 丽, 李佳俊,吴文伟, 肖 春*,, 叶 敏*,
(1. 云南农业大学 云南生物资源保护与利用国家重点实验室,昆明 650201;2. 云南省农业科学院 农业环境资源研究所,昆明 650201)
草地贪夜蛾Spodoptera frugiperda(以下简称FAW),属于鳞翅目、夜蛾科、灰翅夜蛾属,原产于美洲热带和亚热带地区,广泛分布于美洲大陆[1]。2019 年1 月11 日首次在中国云南省江城县发现FAW 幼虫[2],其主要为害玉米、水稻、高粱和甘蔗等粮食作物的芯叶及果穗,造成农作物产量减少和品质下降[3]。化学农药是防治FAW 的重要措施。除虫脲 (diflubenzuron)、茚虫威 (indoxacarb) 以及氯虫苯甲酰胺 (chlorantraniliprole)、氟苯虫酰胺 (flubendiamide) 等双酰胺类杀虫剂是中国防治FAW 的推荐用药,在FAW 的防治中发挥了重要作用[4]。
目前,杀虫剂抗性行动委员会 (Insecticide Resistance Action Committee, IRAC) 分类的双酰胺类杀虫剂共有5 个品种:氯虫苯甲酰胺、氟苯虫酰胺、溴氰虫酰胺 (cyantraniliprole)、环溴虫酰胺(cyclaniliprole) 和四唑虫酰胺 (tetraniliprole)[5]。氯虫苯甲酰胺、氟苯虫酰胺、溴氰虫酰胺等双酰胺类杀虫剂具有杀虫广谱、内吸性好、高选择性及对刺吸式口器害虫防效优异等特点[6-8],且主要作用鳞翅目和鞘翅目昆虫鱼尼丁受体 (ryanodine receptor,RyR),其激活RyR 导致钙离子释放,破坏昆虫体内钙离子平衡,从而影响昆虫的神经肌肉功能[9]。RyR 是一种分布于动物肌细胞内质网和肌浆网上的钙离子通道蛋白,由4 个相同的亚基构成的同源四聚体,每个亚基含有约5 000 个氨基酸残基,是目前已发现的最大的离子通道型蛋白[10]。以RyR 为靶标开发的氟苯虫酰胺和氯虫苯甲酰胺等双酰胺类杀虫剂对小菜蛾Plutella xylostella(以下简称DBM)、斜纹夜蛾Spodoptera litura等鳞翅目昆虫均具有较高的毒性,对哺乳动物则低毒、安全。但是,随着双酰胺类杀虫剂的大量使用,其抗药性问题也日益突出[11-14]。
目前,研究鳞翅目害虫对双酰胺类杀虫剂的抗性多以DBM 为模式害虫。DBM 对双酰胺类杀虫剂已产生了高水平抗性:Guo 等对从广东不同地区采集的4 个DBM 种群进行检测发现,其对氯虫苯甲酰胺的抗性水平达303~658 倍[11];据Troczka 报道,泰国DBM 田间种群对氯虫苯甲酰胺和氟苯虫酰胺的抗性分别高于200 倍和750 倍,菲律宾DBM 田间种群对氯虫苯甲酰胺和氟苯虫酰胺的抗性分别超过4 100 倍和1 300 倍[12]。另外,FAW 对双酰胺类杀虫剂也易产生抗性:2018 年在美国波多黎各田间发现,FAW 种群对氟苯虫酰胺产生了500 倍抗性,对氯虫苯甲酰胺产生了160 倍抗性[13];Bolzan 等发现,室内筛选所得抗氯虫苯甲酰胺FAW 种群与溴氰虫酰胺存在低水平交互抗性 (27 倍),而对氟苯虫酰胺则存在高达42 000 倍的交互抗性[15]。目前,DBM 对双酰胺类杀虫剂的抗药性机制较为明确:Troczka 在泰国和菲律宾的DBM 抗性种群中检测到RyR 的G4946E 突变,通过Sf9 体外表达研究证实了G4946E突变可影响双酰胺类杀虫剂与RyR的结合[12];Steinbach 等进一步强调了DBMRyR的G4946E 突变是导致其对双酰胺类杀虫剂产生抗性的主要机制[16];郭磊等在云南采集的抗性种群中筛查得到了新的突变位点E1338D、Q4594L和I4790M,并通过荧光配体结合实验验证了含多个突变的DBMRyR 与氯虫苯甲酰胺的亲和力显著减弱[11,17-18]。其他鳞翅目害虫中:甜菜夜蛾Spodoptera exigua的抗性种群对氯虫苯甲酰胺的抗性在150 倍以上,通过对其RyR 跨膜区域进行测序发现,抗性种群的主要突变为I4743M,但未检测到G4900E 等位基因突变[14];二化螟Chilo suppressalisRyR 的G4910E 和I4758M的突变也会导致其对双酰胺类杀虫剂产生不同程度的抗性[19-20]。
FAW 对双酰胺类杀虫剂的抗性机制相关研究较少。在现有报道中,Boaventura 对田间采集的FAW 种群进行抗药性诱导,与敏感种群相比,其对氯虫苯甲酰胺和氟苯虫酰胺的抗性分别为225 倍和5 400 倍。通过对FAWRyR 的C-末端结构域Ⅱ~Ⅵ进行测序,发现了一个单核苷酸多态性,导致I4734M 突变,因此被认为是对双酰胺类杀虫剂产生抗性的靶点[21]。在有关DBMRyR 已报道的突变位点中,G4946E 和I4790M 频率最高,其中G4946 对应FAWRyR 中的G4891 氨基酸,I4790 对应FAWRyR 中的I4734 氨基酸。FAWRyR中的I4734 与DBMRyR 的I4790 突变报道一致,但尚未见有关FAWRyR 中的G4891 氨基酸突变引起抗药性的报道。FAWRyR 的G4891 和I4734 氨基酸突变是否是造成其对双酰胺类杀虫剂产生抗性的原因仍需探索。
随着计算机模拟技术的发展,分子对接、同源建模等技术手段日趋成熟。通过计算机预测受体与配体之间的结合模式为研究靶标害虫对杀虫剂的抗性机制提供了新的方法[22]。蛋白质的三维结构由氨基酸序列确定,由于蛋白质的同源性,可以根据已知的蛋白质三维结构预测未知结构的同源蛋白质的初步结构[23]。目前,FAW 和DBM的RyR 结构尚未被解析,但兔子Oryctolagus cuniculus、野猪Sus scrofa等哺乳动物的RyR 的结构解析已较为成熟,且兔子RyR 与氯虫苯甲酰胺结合的结构已被解析 (PDB:7CF9):氯虫苯甲酰胺主要与兔子RyR 蛋白C 端跨膜区域中S0-S4构成的活性口袋结合[24]。FAWRyR (序列号:QCQ29110.1)[21]与DBMRyR (序列号:AET09964.1)[25]的氨基酸序列已知,可通过同源建模的方法获得其三维模型及分子对接的方法分析亲和构象。
因此,本研究拟通过同源建模的方法构建DBMRyR 和FAWRyR 跨膜区域的三维模型,通过在FAWRyR 中引入已报道的DBMRyR 的突变位点 (G4946E,对应FAWRyR 中G4891) 和FAWRyR的突变位点 (I4734M),构建FAWRyR 的突变模型;通过分子对接的方法研究FAWRyR 突变前后与双酰胺类杀虫剂的结合模式,分析FAWRyR 突变位点与抗药性的关系。本研究从结构生物学角度探究FAW 抗双酰胺类杀虫剂的机理,旨在为研究FAW 对双酰胺类杀虫剂的抗性奠定理论基础。
1 材料与方法
1.1 计算机软件与在线工具
1.1.1 软件 Open Babel-2.4.1[26]、UCSF Chimera-1.11.2[27]、Pymol-2.1.0[28]、Autodock Tool-1.5.6[29]和Autodock Vina-1.1.2[30]。
1.1.2 在线工具
同源建模在线工具SWISS-MODEL[31]:https://swissmodel.expasy.org/。
蛋白质质量评价工具SAVEs:https://servicesn.mbi.ucla.edu/SAVES/。
蛋白质晶体结构PDB 数据库:http://www1.rcsb.org/。
NCBI 数据库:https://www.ncbi.nlm.nih.gov/。
跨膜区域预测工具TMHMM[32]:http://www.cbs.dtu.dk/services/TMHMM/。
作用力分析工具PLIP[33]:https://projects.biotec.tu-dresden.de/plip-web/plip。
保守性分析在线工具MEME[34]:(https://memesuite.org/meme/)。
1.2 试验方法
1.2.1 同源建模及模型评价 从NCBI 数据库中下载DBMRyR 氨基酸序列 (序列号:AET09964.1)和FAWRyR 氨基酸序列 (序列号:QCQ29110.1),通过TMHMM 分别预测DBMRyR 和FAWRyR 的跨膜区域。通过同源建模方法构建FAWRyR (未突变,WT) 蛋白三维模型:选择FAWRyR 跨膜区域 (S1-S6) 的氨基酸序列,以Ma 等报道的与FAWRyR氨基酸序列同源度为61.8%的兔子RyR 冷冻电镜结构 (PDB:7CF9)[24]的跨膜区域为模板,用SWISS-MODEL 工具构建FAWRyR 跨膜区域 (S1-S6) 的模型;选择FAWRyR 跨膜区域 (S0) 的氨基酸序列,通过SWISS-MODEL 进行序列对比分析,以PDB:1p7b.1.A 为模板构建FAWRyR 跨膜区域S0 的模型。使用Pymol-2.1.0 和UCSF Chimera-1.11.2 软件对在线够建的模型进行拼合、优化,构建出WT FAWRyR 的四聚体结构。DBMRyR 跨膜区域模型的构建方法亦如此。
使用SAVEs 的Procheck[35]工具,验证所建模型在立体化学上的合理性及能量上的稳定性。各残基间的Pis 和Phi 两个角度的合理性评价结果以拉氏图 (Ramachandran Plot) 展示。根据拉氏图中最优区域、额外允许区域、一般允许区域等的百分比评价所建蛋白质模型是否合理[35]。
1.2.2 FAWRyR 突变体构建 将已报道的DBMRyR 的突变位点G4946E、I4790M,以及甜菜夜蛾S. exiguaRyR 的G4900E、I4743M[14],二化螟C. suppressalisRyR 的G4910E、I4758M[19-20]与FAWRyR 氨基酸序列进行对比,结果表明其分别对应FAWRyR的G4891E 和I4734M,其中I4734M 为已报道的FAWRyR 突变位点[21]。故本研究使用UCSF Chimera-1.11.2 软件分别将FAWRyR中G4891 的“Gly”突变为“Glu”、I4734 的“Ile”突变为“Met”,构建突变模型FAWRyR-E4891 和FAWRyR-M4734。将WT FAWRyR、FAWRyRE4891 及FAWRyR-M4734 的模型分别用Autodock Tool-1.5.6 进行能量最小化和去水加氢等处理,作为分子对接受体。
1.2.3 配体构建 从IRAC 查询双酰胺类杀虫剂:氯虫苯甲酰胺、氟苯虫酰胺、溴氰虫酰胺、环溴虫酰胺和四唑虫酰胺的化学结构,使用Open Babel-2.4.1 软件获得化学结构的3D 结构,利用Autodock Tool-1.5.6 进行能量最小化等处理,作为分子对接配体。
1.2.4 分子对接及结合模式分析
1.2.4.1 分子对接 根据已报道的兔子RyR 与氯虫苯甲酰胺复合晶体结构的结合位点,通过蛋白质模型叠合预测所建模型的活性口袋位置,并构建Docking-Box (图1)。FAWRyRs 对接所用Box 参数为:center-x = 252.259,center-y =207.152,center-z = 299.459;size-x = 22.0,size-y =18.0,size-z = 18.0。
将1.2.1 节和1.2.2 节中处理好的蛋白质受体与1.2.3 节中准备好的配体使用Autodock Vina-1.1.2 软件进行对接,计算平台HP-PC:Inter Corei5-1035G1CPU@3.6 GHz;对接参数为:exhaustive ness = 8,numb-modes = 9,energyrange = 3,对接结果按结合亲和力打分排序。
1.2.4.2 结合模式分析 分别选择5 种配体的最优构象,构建其与受体的复合物,利用PLIP 在线工具分析配体与受体之间的氢键、卤键、疏水作用、π-π 相互作用及π-阳离子作用等非共价相互作用力,使用Pymol-2.1.0 绘制结合模式图。
1.2.5 基因序列保守性分析 将FAWRyR 跨膜区域的蛋白质序列作为查询序列,在NCBI 中使用BLASTP 查询相关同源蛋白。最终得到100 条与FAWRyR 同源的蛋白氨基酸序列,且主要以鳞翅目昆虫为主 (表1)。将这100 条RyR 氨基酸序列用MEME (https://meme-suite.org/meme/) 工具计算分析,获得Motif 图,并进行保守性分析。

表1 与FAWRyR 跨膜区域氨基酸序列具有同源的昆虫基因数量Table 1 The number of insect genes with homologous amino acid sequences with FAWRyR of transmembrane region
2 结果与分析
2.1 同源建模结果及评价
根据DBMRyR 和FAWRyR 的氨基酸序列,分别以PDB:7CF9 的跨膜区域为模板,构建的跨膜区域S1-S6;以同源较高的PDB:1p7b.1.A 为模板,在线构建的跨膜区域S0,拼合后所得最终RyR 四聚体结构如图2a、2d 所示。由图2a、2d 可知:FAW 和DBM 的RyR 的四个亚基呈轴对称分布,每个亚基的氨基酸序列相同,中间有一由亲水性氨基酸残基围成的孔,钙离子从中通过[36];由图2b、2e 可知,跨膜区域主要由几组平行的α 螺旋构成,展示出明显的跨膜蛋白特性。
WT FAWRyR 和DBMRyR 的模型评价结果如图2c、2f 所示:WT FAWRyR 及DBMRyR 的Procheck 评估结果分别为:最优区域残基占比87.4%、89.8%,额外允许区残基占比11.7%、9.5%,两者残基位于允许区域的比例均大于90%,模型合理。
将WT FAWRyR 与DBMRyR 的模型进行叠合对比 (图3a、3b),并把DBMRyR 跨膜区域的氨基酸序列与WT FAWRyR 跨膜区域的蛋白模型进行对比 (图3c),三维结构对比结果显示:两者的RMSD 值为0.082,其三维模型差异较小。二维序列对比结果表明:两者的同源度为92.55%,覆盖度为99%,具有较高的同源性。
2.2 分子对接结果分析
2.2.1 双酰胺类杀虫剂与WT FAWRyR 的结合模式分析 对接方法验证:将WT FAWRyR 与5 种双酰胺类杀虫剂分别对接后 (保留9 个构象),根据对接亲和力打分进行排序,并选择最佳对接构象与已解析的兔子RyR 与氯虫苯甲酰胺的复合晶体进行叠合对比。结果表明:预测的配体构象与实际构象相似,结合模式合理,证明对接方法可行。
WT FAWRyR 与氯虫苯甲酰胺、氟苯虫酰胺、溴氰虫酰胺、环溴虫酰胺及四唑虫酰胺最优构象的结合亲和力分别为:-31.813 6、-35.581、-33.488、-30.139 2 和-38.092 6 kJ/mol。对结合模式进行分析可知:L4458、Y4462、Y4643、L4646 和F4892 与氯虫苯甲酰胺的苯环及吡啶环形成疏水作用;K4642 与吡啶环上的N 原子形成氢键;Y4639 与吡啶环形成π-π 相互作用;K4642的N 原子与吡唑环形成π-阳离子作用;G4891与吡啶环上的Cl 原子形成卤键 (图4a)。L4458、T4863、V4890 与氟苯虫酰胺的苯环形成疏水作用;K4642 与羰基上的O 原子形成氢键,R4896与磺酰基上的O 原子形成氢键,V4890、G4891与酰胺上的NH 形成氢键;R4896 的N 原子与苯环形成π-阳离子作用 (图4b)。L4458、Y4462、Y4643、L4646 和F4892 与溴氰虫酰胺的苯环、吡啶环形成疏水作用;K4642、G4891 分别与吡啶环上的N 原子和吡唑环侧酰胺结构NH 形成氢键;Y4639 与吡啶环形成π-π 相互作用;K4642与吡唑环形成π-阳离子作用;G4891 与吡啶环上的Cl 原子形成卤键 (图4c)。L4458、L4646、Y4863、V4890 和F4892 与环溴虫酰胺的吡啶环及环丙基形成疏水作用;K4642 与吡唑环的N 原子形成氢键,G4891 与环丙基侧酰胺结构的NH 形成氢键,R4896 与吡唑环侧羰基的O 原子形成氢键;K4642、R4896 的N 原子分别与吡啶环和吡唑环形成π-阳离子作用 (图4d)。Y4462、Y4643、L4646 和F4892 与四唑虫酰胺的苯环形成疏水作用;K4642、G4891 分别与吡啶环上的N 原子和吡唑环侧酰胺结构的NH 形成氢键;Y4639 和Y4863 分别与吡啶环和四唑环形成π-π 相互作用;K4642 的N 原子与吡唑环形成π-阳离子作用;S4860 与三氟甲基上的F 原子形成卤键 (图4e)。由于存在疏水作用、π-π 相互作用、π-阳离子作用及分子间形成的氢键和卤键,使分子能稳定存在。5 种杀虫剂与WT FAWRyR 对接后的位置如图4f 所示,可见其位置都位于预测的活性口袋内。
2.2.2 双酰胺类杀虫剂与FAWRyR-E4891 的结合模式分析 FAWRyR-E4891 与氯虫苯甲酰胺、氟苯虫酰胺、溴氰虫酰胺、环溴虫酰胺及四唑虫酰胺最优构象的结合亲和力分别为:-26.371 8、-29.720 6、-28.464 8、-27.209 和-32.650 8 kJ/mol。对结合模式进行分析可知:Y4639、K4642 和Y4738 与氯虫苯甲酰胺的苯环上的甲基和吡啶环形成疏水作用;Y4639 和K4642 分别与苯环侧酰胺结构的NH 和吡啶环上的N 原子形成氢键;Y4639 与苯环形成π-π 相互作用;K4642 的N 原子与吡啶环形成π-阳离子作用 (图5a)。F4638、Y4639、Y4738、K4642、K4742 和Y4863 与氟苯虫酰胺的苯环及甲基形成疏水作用;K4642 与苯环侧羰基上的O 原子形成氢键;K4642 的N 原子与苯环形成π-阳离子作用;Y4639 与苯环形成ππ 相互作用;Y4639 与三氟甲基上的F 原子形成卤键(图5b)。L4458、Y4639、K4642 和Y4738 与溴氰虫酰胺的苯环、苯环上的甲基和吡啶环形成疏水作用;Y4639、K4642 分别与酰胺结构NH 和吡唑环侧上羰基的O 原子形成氢键;Y4639 与苯环形成π-π 相互作用,K4642 的N 原子与吡啶环形成π-阳离子作用 (图5c)。Y4639、K4642、Y4863、V4890 和R4896 与环溴虫酰胺的吡啶环及环丙烷等形成疏水作用;R4896 与吡唑环侧羰基的O 原子形成氢键;K4642 和R4896 的N 原子分别与吡唑环和吡啶环形成π-阳离子作用 (图5d)。V4890与四唑虫酰胺的吡啶环形成疏水作用;K4642 与四唑环上的N 原子形成氢键;K4642 的N 原子与四唑环、吡唑环形成π-阳离子作用 (图5e)。FAWRyRE4891 与5 种杀虫剂对接的位置如图5f 所示,可见其5 种构象的位置都位于预测的活性口袋内。
2.2.3 双酰胺类杀虫剂与FAWRyR-M4734 的结合模式分析 FAWRyR-M4734 与氯虫苯甲酰胺、氟苯虫酰胺、溴氰虫酰胺、环溴虫酰胺及四唑虫酰胺最优构象的结合亲和力分别为:-32.232 2、-35.581、-33.906 6、-29.720 6 和-38.512 2 kJ/mol。对结合模式进行分析可知:L4458、Y4462、Y4643、L4646 和F4892 与氯虫苯甲酰胺的苯环和吡啶环形成疏水作用;K4642 与吡啶环上的N 原子形成氢键;Y4639 与吡啶环形成π-π 相互作用;K4642的N 原子与吡唑环形成π-阳离子作用;G4891 与吡啶环上的Cl 原子形成卤键 (图6a);L4458、Y4738、K4742、Y4863 和V4890 与氟苯虫酰胺的苯环及甲基形成疏水作用;K4642 和K4742 分别与磺酰基上的O 原子形成氢键,V4890 和G4891与苯环侧酰胺结构的NH 形成氢键;R4896 的N 原子与苯环形成π-阳离子作用 (图6b);L4458、Y4462、Y4643、L4646 和F4892 与溴氰虫酰胺的苯环和吡啶环形成疏水作用;K4642、G4891 分别与吡啶环的N 原子和酰胺结构的NH 形成氢键;Y4639 与吡啶环形成π-π 相互作用,K4642 与吡唑环形成π-阳离子作用;G4891 与吡啶环上的Cl 原子形成卤键 (图6c);L4458、L4646、Y4863、V4890 和F4892 与环溴虫酰胺的吡啶环及环丙烷形成疏水作用;K4642 与吡唑环上的N 原子形成氢键,G4891 与环丙基侧酰胺的NH 形成氢键,R4896 与丙三环侧羰基的O 原子形成氢键;R4896的N 原子与吡啶环形成π-阳离子作用 (图6d);Y4462、Y4643、L4646 和F4892 与四唑虫酰胺的苯环形成疏水作用;K4642 与吡啶环上的N 原子形成氢键,G4891 与苯环侧酰胺结构的NH 形成氢键;Y4639 和Y4863 分别与吡啶环和四唑环形成π-π 相互作用;K4642 的N 原子与吡唑环形成π-阳离子作用;S4860 与三氟甲基上的F 原子形成卤键 (图6e);FAWRyR-M4734 与5 种杀虫剂对接的位置如图6f 所示,可见其5 种构象的位置都位于预测的活性口袋内。
2.3 FAW 突变位点分析
由表2 可知:FAWRyR 的4734 位异亮氨酸(Ile) 突变为蛋氨酸 (Met) 后,与双酰胺类杀虫剂的结合亲和力未下降;FAWRyR 的4891 位甘氨酸(Gly) 突变为谷氨酸 (Glu) 后,与双酰胺类杀虫剂的结合亲和力明显降低。
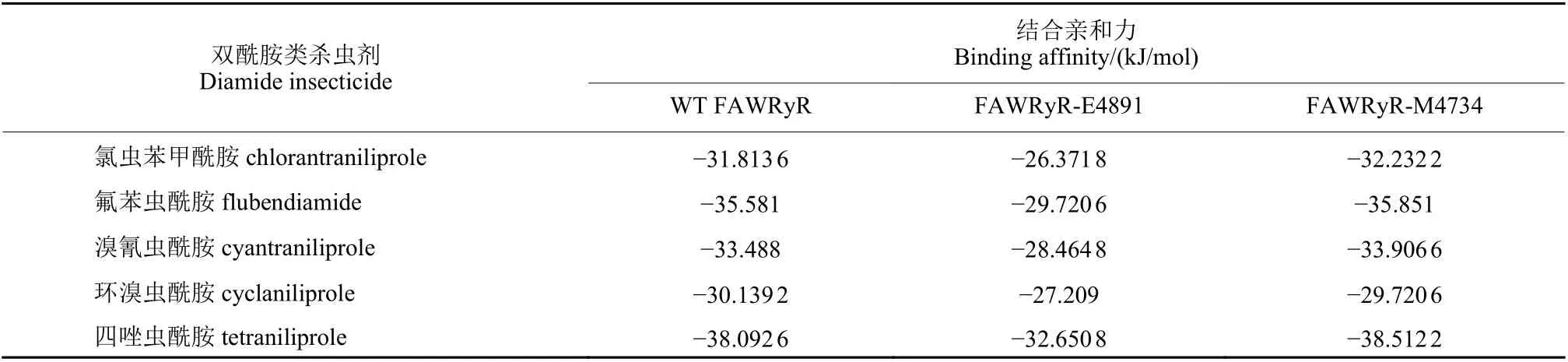
表2 FAWRyR 突变前后与双酰胺类杀虫剂的结合亲和力变化Table 2 The binding affinity changes of the wild-type (WT) & mutant FAWRyRs and diamides
五种双酰胺类杀虫剂与WT FAWRyR、FAWRyRE4891 和FAWRyR-M4734 的结合模式如图7 所示:将FAWRyR 的I4734 突变为M4734 后,与双酰胺类杀虫剂之间的作用力方式、参与形成作用力的残基和WT FAWRyR 基本一致,初步分析其主要原因是FAWRyR 的4734 号异亮氨酸(Ile) 不参与配体与受体之间作用力的形成。而FAWRyR 的4891 号甘氨酸 (Gly) 与双酰胺类杀虫剂之间存在氢键、卤键相互作用,突变为谷氨酸(Glu) 后,不能与双酰胺类杀虫剂形成氢键、卤键,导致受体与配体之间的作用力方式减少、亲和力下降。FAWRyR 中G4891 突变为E4891 可能是导致FAW 对双酰胺类杀虫剂产生抗药性的主要原因,而I4734M 突变可能与抗药性的产生无关。
2.4 氨基酸保守性分析
由MEME 工具分析得到Motif 图 (图8、图9),图中FAWRyR 跨膜区域的活性口袋由L4458、Y4461、L4641、Y4462、Y4639、K4642、Y4643、S4860、F4861、S4864、V4888、V4890、G4891、F4892 和K4893 构成 (如图8 所示),其中Y4461、L4641、Y4462、Y4639、K4642、Y4643、S4860、F4861、S4864、V4888、V4890、G4891、F4892、K4893 均在保守区域,而L4458 在不保守区域,存在L4458Y 突变的可能 (如图9 所示),但尚未见L4458 氨基酸突变为Y4458 氨基酸的相关报道,且L4458 氨基酸不与双酰胺类杀虫剂形成氢键等关键相互作用,L4458Y 突变引起双酰胺类抗药性的几率较低。而G4891 氨基酸虽在FAWRyR的保守区域,但前文研究表明,G4891E 突变会引起FAWRyR 与双酰胺类杀虫剂的结合亲和力下降,因此,引起抗药性的G4891E 突变可能为随机突变。
3 讨论
解析鳞翅目害虫RyR 与双酰胺类杀虫剂的结合模式一直是国内外研究的热点。Steinbach 等根据Yan 等[37]解析的兔子RyR (PDB:3J8H) 的结构构建了DBMRyR 的同源模型,在此模型上引入G4946E 和I4790M 突变,并分析了双酰胺类杀虫剂与DBMRyR 的结合模式,提出RyR 与双酰胺类杀虫剂的结合位点可能在这两个突变位点附近,但双酰胺类杀虫剂与DBMRyR 的结合模式、相互作用力以及形成作用力的残基等尚不明确[16]。2020 年,Ma 等使用冷冻电镜解析了兔子RyR 和氯虫苯甲酰胺复合体的结构 (PDB:7CF9),揭示了兔子RyR 中氯虫苯甲酰胺的结合口袋、作用方式以及参与结合的残基[24],但FAWRyR 与双酰胺类杀虫剂的结合模式尚不明确。另外,FAW 对双酰胺类杀虫剂的抗性机制的相关研究较少。因此,本研究在已有研究的基础上,选择PDB:7CF9为模板,同源构建FAWRyR 的三维模型,并使用分子对接方法分析FAWRyR 与双酰胺类杀虫剂的结合模式。结果表明:5 种双酰胺类杀虫剂与FAWRyR 结合的活性口袋以及两者之间形成作用力的氨基酸残基基本一致。该结果与Qi 等报道的“氯虫苯甲酰胺和氟苯虫酰胺虽结构上略有差异,但结合位点相同”[38-39]的观点一致。
目前,鳞翅目害虫对双酰胺类杀虫剂的抗性可能与DBMRyR 的G4946E、I4790M,S. exiguaRyR 的I4743M 以及C. suppressalisRyR 的G4910E和I4758M (均对应FAWRyR 的G4891、I4734) 突变有关。鳞翅目害虫RyR 跨膜区域氨基酸序列同源度一般在92%以上[40],本研究通过对比WT FAWRyR和DBMRyR 跨膜区域的氨基酸序列,发现两者的同源度高达92.55%;对比两者RyR 三维模型,其RMSD 值为0.082,两者同源度极高。因此,本研究根据DBMRyR 的G4946E 和I4790M 突变构建了FAWRyR-E4891 和FAWRyR-M4734 突变体,并分析了其与双酰胺类杀虫剂的结合模式。结果表明:FAWRyR 的G4891 突变为E4891 后,与双酰胺类杀虫剂的亲和力下降。目前已有研究结果表明,DBMRyR、C. suppressalisRyR 的G4946E 和G4910E (均对应FAWRyR 的G4891) 突变均可能导致其对双酰胺类杀虫剂产生抗性。因此,FAWRyR的G4891E 突变,可能是FAW 对双酰胺类杀虫剂产生抗性的原因之一。在今后的研究中,需重点关注FAWRyR 的G4891E 突变。现有研究认为,FAWRyR 的I4734M 突变也可能与FAW 对氯虫苯甲酰胺和氟苯虫酰胺的抗性有关[21]。但本研究发现,FAWRyR 的4734 号氨基酸不参与其与双酰胺类杀虫剂的结合,I4734 突变为M4734 后FAWRyR 与双酰胺类杀虫剂的亲和力并未下降。另外,Ma 等的研究结果表明:DBMRyR 的I4790(对应FAW 的I4734) 也不是其与双酰胺类杀虫剂形成亲和力的主要残基[24]。Boaventura 等在研究FAWRyR 的4734 号氨基酸突变与抗药性关系时所测田间幼虫样本数量有限,田间样本中I4734M突变频率较低[21],I4734M 突变是否能够导致FAW 对双酰胺类杀虫剂产生抗性仍需进一步研究探索。本研究也表明:G4891 在FAWRyR 的保守区域,引起抗药性的G4891E 突变可能为随机突变。除G4891 外,K4642、V4890 和R4896 与双酰胺类杀虫剂也可能存在氢键相互作用,同时也存在随机突变的可能性。因此,在后期研究中,应重点关注这3 个氨基酸的突变,并进一步研究该突变与其对双酰胺类杀虫剂抗性的关系。另外在未来研究中,除了使用通过分子对接、氨基酸序列保守性分析的方法外,还可以考虑使用突变预测工具,如:AIMMS[41]、KRDS[42]在线工具和生物学网络建模方法[43]等预测其他随机突变与抗药性之间的关系。
目前,本研究仅通过同源建模方法构建了FAWRyR 突变前后的三维模型,通过分子对接方法模拟FAWRyR 突变前后与双酰胺类杀虫剂的结合模式,能够为分析FAW 对双酰胺类杀虫剂的抗性机制提供理论依据。但是,分子对接的方法只能初步分析配体与受体之间的亲和力,而分子动力学模拟能更加准确地计算配体与蛋白受体的绝对结合自由能,其在药物结合靶标研究中被广泛应用[44-45]。LAMMPS、Gromacs、NAMD 等传统的分子动力学模拟软件操作繁琐,不适合进行大量的结合模式分析和结合自由能计算工作。目前也有一些图形化的分子动力学模拟在线工具 (如:LARMD[46]) 正在研发中,在未来研究中可以考虑运用这些工具进一步分析配体与受体之间的结合模式,计算亲和力。另外,本研究通过计算机模拟得出的理论计算结果并未经过实际的分子生物学实验验证,且同源建模和分子对接方法仅是计算机模拟方法,无法完全准确模拟受体和配体之间的结合模式。在今后研究中,可以通过分子生物学实验进一步验证该文的理论计算结果,以及可以使用冷冻电镜或X-ray 衍射法分析FAWRyR与双酰胺类杀虫剂复合体的结构,精确分析两者之间的结合模式。
4 结论
本研究通过蛋白质模建、分子对接等计算机分子模拟方法,分析了WT FAWRyR 与双酰胺类杀虫剂的结合模式;根据已报道的鳞翅目害虫RyR 的突变位点,构建FAWRyR 突变体,并对其分析与5 种双酰胺类杀虫剂的结合模式。主要结论有:
1) 通过同源建模的方法构建了FAWRyR 的C 端跨膜区域及其突变体的三维模型。
2) 通过对比分析FAWRyR 突变前后与5 种双酰胺类杀虫剂的结合模式,发现G4891E 突变会导致FAWRyR 与双酰胺类杀虫剂的结合亲和力下降、形成的作用力方式减少,从而导致FAW 对双酰胺类杀虫剂产生抗性。
本研究从结构生物学角度初步分析了FAW 抗双酰胺类杀虫剂的机理,能够为研究FAW 对双酰胺类杀虫剂的抗性提供线索,同时为今后杀虫剂分子设计和结构修饰提供参考。在今后研究中可结合分子生物学实验、冷冻电镜或X-ray 衍射法进一步验证FAW 抗双酰胺类杀虫剂的机理。
