蛤蚧冬眠期与非冬眠期消化道嗜银细胞的比较
2021-10-16赵成坚徐永莉姜建萍
赵成坚 黄 勇 刘 武 徐永莉 姜建萍 霍 娟*
1.广西壮族自治区药用植物园,南宁 530023;2.广西中医药大学,南宁 530200
消化道不仅是动物机体主要的消化器官,也是体内最大、最复杂的内分泌器官,其组织中含有种类繁多、功能各异的内分泌细胞,嗜银细胞是其中的一种,与神经系统和外分泌腺相互配合,共同调控消化吸收及体内其他生理过程。已有不少学者对哺乳动物如山羊[1]、猪[2]、金钱豹[3],鸟类如黄雀[4]、鹌鹑[5],鱼类如花[6],爬行动物如乌苏里蝮蛇[7]、草蜥[8]、胎生蜥蜴[9]等动物的消化道嗜银细胞进行研究,研究结果表明:不同种类动物消化道嗜银细胞的类型及分布存在较大差异,同种类的动物如爬行动物既有差异也有相似性,但不是绝对相同,与食性及栖息地环境有一定的关系[10]。
蛤蚧属于脊索动物门、爬行纲、有鳞目、壁虎科、壁虎属动物,为我国传统的珍稀濒危动物药材,具有调节免疫、抗肿瘤、平喘、抗炎及抗衰老等作用。主要分布于我国的广西、广东和云南南部及越南、缅甸北部。每年11月至第2年3月底,气温低于18 ℃时,蛤蚧进入冬眠期,此时的蛤蚧少进食或者不进食,活动量明显减少。已有学者对两栖纲蛙类如林蛙[11]、中华蟾蜍、黑斑蛙[12]在冬眠期与非冬眠期的消化道嗜银细胞进行研究,而爬行纲变温动物蛤蚧在冬眠期与非冬眠期消化道嗜银细胞的研究至今未见报道。本文拟对这2个时期的蛤蚧消化道嗜银细胞进行比较研究,以期为探讨嗜银细胞是否参与冬眠期的消化调节活动提供参考依据。
1 材料与方法
1.1 材 料
1)试验动物。本试验所用的蛤蚧来自广西壮族自治区药用植物园养殖基地,体格健康,雌雄不限,平均体重约60 g。冬眠期取样时间为1月,非冬眠期取样时间为6月,每个时期样本数为3条。
2)试验试剂。Bouin 固定液:40%甲醛溶液25 mL,冰醋酸5 mL,苦味酸饱和水溶液70 mL;硝酸银染液:1%硝酸银水溶液5 mL,0.2 mol/L 醋酸钠缓冲液(pH 5.6)10 mL,双蒸水85 mL;还原液:无水亚硫酸钠5 g,对苯二酚1 g,双蒸水100 mL;5%硫代硫酸钠水溶液。
1.2 方 法
1)取样。取冬眠期与非冬眠期的蛤蚧各3 条,采用毁髓法(解剖针从头颈部插入捣毁脑组织和脊髓)处死,解剖将胃部及肠道取出,用0.9%生理盐水冲洗。分别取胃贲门、胃体、胃幽门、十二指肠、空肠、回肠,每段约0.5 cm,浸泡于Bouin 固定液中48 h,然后流水冲洗几次,放入70%乙醇中保存备用。
2)石蜡切片。将材料取出,流水冲洗后进行常规石蜡切片步骤,即脱水、浸蜡、包埋、修块、切片。
3)脱蜡至水。依次将切片放入二甲苯Ⅰ20 min→二甲苯Ⅱ20 min→无水乙醇Ⅰ5 min→无水乙醇Ⅱ5 min→75%乙醇5 min,自来水洗,再用蒸馏水洗3次。
4)染色及还原。在60 ℃硝酸银染液恒温浸染3 h 后,用纸吸干组织周围的染液。放入45 ℃还原液中还原1 min,蒸馏水洗2 次,5%硫代硫酸钠水溶液2 min,蒸馏水洗数次。
5)脱水封片。切片依次放入无水乙醇Ⅰ5 min→无水乙醇Ⅱ5 min→无水乙醇Ⅲ5 min→二甲苯Ⅰ5 min →二甲苯Ⅱ5 min 透明,中性树胶封片。
6)显微镜检及细胞计数。在200倍光镜下对冬眠期与非冬眠期的蛤蚧的食道、胃体、胃幽门、十二指肠、空肠、回肠等6个部位嗜银细胞的分布情况进行观察,每个时期每个部位选取5个视野,用SPSS 22.0统计分析软件中的Duncan's 多重比较的方法对不同时期消化道各部位嗜银细胞密度进行统计学分析,并运用t检验对同一部位不同时期的嗜银细胞密度进行统计学分析,取“平均值±标准差”表示样本嗜银细胞的分布数量。
2 结果与分析
由于嗜银细胞有亲银的特性,经过银染液染色后,嗜银细胞呈黑色或棕黑色,背景为黄色或浅黄色,在光镜下对比较为明显,容易辨认。
2.1 嗜银细胞的形态特征
从切片镜检的结果来看,冬眠期蛤蚧消化道嗜银细胞的形态主要以不规则形、椎体形为主,而非冬眠期蛤蚧消化道嗜银细胞形态主要以椭圆形、长梭形、圆形为主,主要分布于黏膜上皮细胞之间、上皮细胞基部及腺部上皮细胞之间,2 个时期的形态特征及分布密度见图1~图8。

图1 冬眠期蛤蚧食道嗜银细胞

图2 非冬眠期蛤蚧食道嗜银细胞

图3 冬眠期蛤蚧胃体嗜银细胞

图4 非冬眠期蛤蚧胃体嗜银细胞

图5 冬眠期蛤蚧幽门嗜银细胞
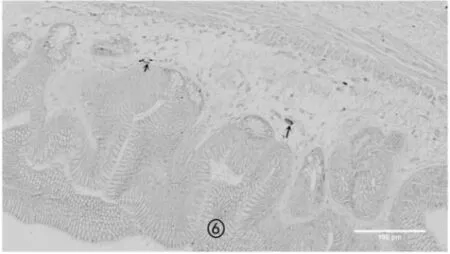
图6 非冬眠期蛤蚧幽门嗜银细胞

图7 冬眠期蛤蚧十二指肠嗜银细胞
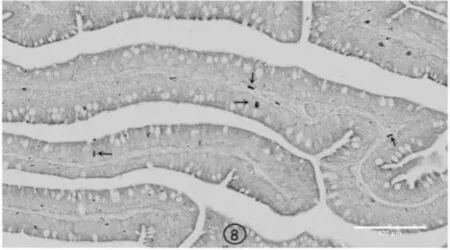
图8 非冬眠期蛤蚧十二指肠嗜银细胞
2.2 冬眠期与非冬眠期蛤蚧嗜银细胞的分布情况及密度
冬眠期蛤蚧胃部的嗜银细胞分布密度高峰位于幽门,低谷位于胃体;肠道部的嗜银细胞分布高峰位于十二指肠,低谷位于回肠。非冬眠期的分布高峰在食道。同一部位处于不同时期时的嗜银细胞分布密度差异极显著,冬眠期各个部位的嗜银细胞数量明显高于非冬眠期的数量(表1)。

表1 蛤蚧冬眠期与非冬眠期消化道各段嗜银细胞的密度
3 讨 论
蛤蚧作为没有口腔咀嚼能力的变温动物,其胃肠道消化能力特别强,可将成型的昆虫如蝗虫、土鳖虫、蟑螂的角质消化吸收,并且具有极强的耐饥饿能力,在长达3~4 个月的冬眠期间可不进食,可见其胃肠道内分泌细胞在营养吸收和能量转化过程中扮演十分重要的角色。作为主要的消化道内分泌细胞——嗜银细胞以分泌5-HT 为主,具有调节黏液分泌、胃肠蠕动、平滑肌收缩和血管扩张的功能。从组织切片结果来看,在各个时期从胃部到肠道均可看到有嗜银细胞分布,食道与胃幽门嗜银细胞的分布密度明显高于胃体,呈“U”形分布,这与王晓平等[13]报道的结果相一致。食物经过食道及胃贲门的挤压进入胃体,食道的嗜银细胞在感受吞食时机械压力刺激分泌激素,对食物起润滑和反馈调节作用,同时防止食物及胃液反流入食管。胃幽门控制胃的排空速度,因此需要更多的嗜银细胞参与调节[14]。华子暄等[15]将哺乳动物嗜银细胞在小肠的分布规律归纳为3 种类型,即第1 种类型是在十二指肠处嗜银细胞分布的数量最多,然后分布数量逐渐降低,在回肠处最低。目前,依据所研究的爬行动物消化道内嗜银细胞的分布情况,约有2/5 的物种分布密度高峰位于胃幽门[16]。蛤蚧便是其中之一。第2 种类型是嗜银细胞在小肠形成一个“U”形的密度分布曲线,即空肠处数量极少,而十二指肠与回肠处数量多。楚德昌等[12]对中华蟾蜍的研究结果属于这种类型。第3种类型是嗜银细胞在小肠中的分布呈高低变动的波浪形,王淼等[14]对东亚腹链蛇的研究结果属于这一类型。
从研究结果看,蛤蚧在不同时期消化道内的嗜银细胞形态各异,冬眠期嗜银细胞的形态特征主要以不规则形、椎体形为主,非冬眠期嗜银细胞的形态特征主要以圆形、椭圆形为主;蛤蚧冬眠期消化道各部位的嗜银细胞分布密度显著高于非冬眠期的各部位器官,这结果与同属于爬行纲有鳞目动物胎生蜥蜴[16]在饥饿状态下嗜银细胞分布情况相似,也与楚德昌等[12]对冬眠期与非冬眠期的中华蟾蜍和黑斑蛙消化管嗜银细胞的研究结果相似,表明冬眠期动物消化道活动虽然明显减弱,但其分泌机能储备反而有明显增加,但与王鸽等[17]对饥饿小鼠嗜银细胞的研究结果存在差异。由此可见,禁食处理及冬眠这一生物学行为特性对嗜银细胞的分泌会因物种的不同而有不同的影响,与食性、环境温度等因素相关。
冬眠是指一些动物在冬季随体温和代谢降低从而呈现出一种昏睡的状态,是对季节性能量短缺的一种生理适应。在人工饲养条件下,虽然可以人为提供充足的食物,但由于环境温度的降低及自身的生物学特性,蛤蚧停止了进食及活动,机体体温被调节到接近于环境温度的水平,心率、代谢率及其他生理活动也相应降低。长时间的低温和禁食,严重影响肠道的正常功能和肠道菌群结构。目前,对于引发蛤蚧冬眠的机制尚未明确,有待进一步研究。
