梅干菜和腌制雪菜细菌菌群多样性分析
2021-09-29虞任莹彭思佳肖庆霞郭斯统罗海波
虞任莹,彭思佳,李 艺,肖庆霞,郭斯统,罗海波,
(1.南京师范大学食品与制药工程学院,江苏南京 210023;2.宁波市鄞州三丰可味食品有限公司,浙江宁波 315145)
雪菜(Brassica juncea(L.)Czern. et Coss. var.multicepsTsen et Lee)又名雪里蕻,属十字花科芸薹属,是芥菜的变种,也称叶用芥菜[1]。雪菜在我国南北各地均有种植,尤其在江浙一带栽培甚广。雪菜采收后除少部分直接用于鲜食外,通常采用干法腌制和盐水泡制的方法加工成梅干菜和泡酸菜产品进行保藏[2]。雪菜腌制成的梅干菜色泽油光黄黑、滋味鲜美、香气浓郁、风味独特,同时还具有解暑热,洁脏腑,消积食,治咳嗽,生津开胃等功效[3],而雪菜经盐水泡制发酵后富含有机酸、醇类、酯类及氨基酸等多种风味物质,呈现出特有的鲜香味,深受国内外消费者喜爱[4]。传统梅干菜和腌制雪菜多为家庭自制,生产周期过长且盐分较高,造成市场上销售的产品品质参差不齐,产品档次不高[5]。
微生物是影响梅干菜和腌制雪菜等发酵类蔬菜营养成分、挥发性物质、风味及品质差异的关键因素,同时也受蔬菜原料种类及不同地域和腌制环境条件的制约。近年来,关于梅干菜和腌制雪菜中发酵微生物方面的研究主要集中在乳酸菌的分离鉴定[6]、挥发性风味物质[7−8]、酚酸及其抗氧化活性[9−10]、低盐腌制[11]等,对梅干菜和腌制雪菜微生物菌群结构及多样性的研究报道较少。尚雪娇等[12]采用Illumina MiSeq高通量测序技术对恩施地区雪里蕻梅干菜菌群多样性进行解析,乳杆菌属(Lactobacillus)、假单胞菌属(Pseudomonas)、葡萄球菌属(Staphylococcus)等8种相对丰度大于1.0%的菌属;陈希等[13]采用16S rDNA克隆文库法对雪菜低盐腌制过程中微生物群落多样性进行分析,发现腌制初期主要优势菌群为肠杆菌属(Enterobacter)和欧文菌属(Erwinia),发酵中后期主要优势菌群为植物乳杆菌(Lactobacillus plantarum)。谢国芳等[6]对贵州地区雪里蕻腌菜中的微生物进行分离筛选,结果表明弯曲乳杆菌(Lactobacillus curvatus)、植物乳杆菌(Lactobacillus plantarum)和希腊魏斯氏菌(Weissella hellenica)为主要优势菌。因此,开展梅干菜和腌制雪菜微生物多样性研究,对推动传统梅干菜和腌制雪菜的工业化生产具有重要意义。
本文以宁波地区雪里蕻梅干菜和腌制雪菜为研究对象,采用Illumina Miseq高通量测序技术分析梅干菜半成品、成品及腌制雪菜成品中菌群结构及多样性,比较干法腌制和盐水泡制发酵蔬菜产品微生物菌群多样性差异,为传统腌制蔬菜微生物资源的挖掘及其工业化应用提供理论依据。
1 材料与方法
1.1 材料与仪器
雪里蕻梅干菜半成品、梅干菜成品、腌制雪菜成品(图1) 宁波市鄞州三丰可味食品有限公司提供;MagicPure Size Selection DNA Beads 北京Transgen(全式金)生物技术有限公司;Qubit3.0 DNA检测试剂盒 美国Life公司;2×Taq Master Mix 南京Vazyme(诺唯赞)生物科技公司;E.Z.N.A.Soil DNA试剂盒

图 1 梅干菜、腌制雪菜产品图Fig.1 Image of the pickled and dried mustard and the fermented potherb mustard
美国OMEGA公司。
AUY220电子分析天平 日本SHIMADZU(岛津)公司;Pico-21型台式离心机 美国Thermo Fisher公司;32866型Qubit 3.0荧光计、Q32866型Qubit T100TM Thermal 美国Invitrogen公司;pHS-3C型精密pH计 上海三信仪表厂;GL-88B型漩涡混合器 海门市其林贝尔仪器制造有限公司;Cyeler型PCR仪 美国BIO-RAD公司;DYY-6 C型电泳仪电源、DYCZ-21型电泳槽 北京市六一仪器厂;凝胶成像系统 美国UVP公司。
1.2 实验方法
1.2.1 样品制作及取样 梅干菜半成品:添加雪菜重量6%的食盐,常温下腌制2个月,经清洗、挑选、切段后,烘干至水分含量15%~20%,2020年5月下旬取样;梅干菜成品:梅干菜半成品装坛密封,常温发酵2个月,2020年7月下旬取样;腌制雪菜成品:雪菜经6%盐水常温泡制发酵2个月至色泽金黄,2020年5月下旬取样。三组样品分别采取随机抽样方法取样1 kg备用,共采集三个平行样品。
1.2.2 样品预处理 在无菌超净操作台上,将梅干菜半成品,梅干菜成品和腌制雪菜成品剪成碎渣,备用。
1.2.3 DNA的提取 采用基因组DNA提取试剂盒提取梅干菜半成品、梅干菜成品和腌制雪菜成品样本微生物基因组DNA,并用琼脂糖凝胶电泳检测。
1.2.4 PCR扩增及高通量测序 以提取的基因组DNA为模板,扩增16S rDNA序列的V3~V4区域,并在梅干菜半成品、梅干菜成品和腌制雪菜成品样本的正向引物上连接含不同碱基的标签条码序列以达到区分目的。采用PCR仪扩增后用2%的琼脂糖凝胶电泳检测。随后进行Illumina Miseq高通量测序。
1.2.5 高通量测序结果分析 梅干菜半成品、梅干菜成品和腌制雪菜成品样本通过Illumina Miseq高通量测序处理后得到双端序列,本实验根据连接不同碱基的标签,找到对应引物之间的接口,取出接头序列。根据PE reads(读取)之间的overlap(覆盖)关系进行拼接,再依据barcode(条码)序列和引物序列进行区分和比较得到有效序列。最后,根据序列质量(reads质量和merge效果)进行质量控制和筛选,过滤复杂且难处理的短序列,去除末端质量值20以下的序列,得到各个样本的有效序列。
在16S rDNA高通量测序过程中,97%的相似性进行OTU(操作分类单元)聚类分析和物种分类学分析。根据OTU聚类分析的结果,研究各个样品微生物的菌群多样性。通过Alpha多样性来分析不同样品的菌群多样性和丰富度,并运用Ace、Chao1、Shannon、Simpson以及Coverage指数来进行评估。其中ACE、Chao1指数用于菌群丰度的评估,而Shannon、Simpson指数则是对菌群多样性进行评估[14]。Coverage指数是测序深度指标,表示每个样本文库的覆盖率。根据分类学信息,在各个水平上统计群落结构并讨论梅干菜半成品、梅干菜成品和腌制雪菜成品微生物群落特征。
1.3 数据处理
利用软件Usearch 5.2.236进行OTU分析,Mothur 1.30.1进行Alpha多样性指数和稀疏性曲线分析,R 3.2作OTU分布韦恩图和稀疏性曲线图,采用Excel 2016作细菌群落结构分布图。
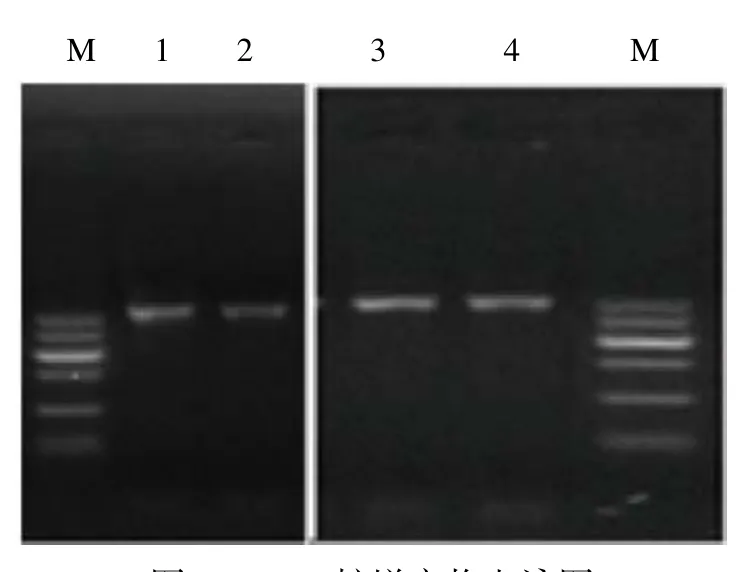
图 2 PCR扩增产物电泳图Fig.2 Electrophoretogram of polymerase chain reaction products
2 结果与分析
2.1 PCR扩增结果
细菌的16S rDNA具有分子大小适中、突变率低等特点,另外因其具有物种特异性,常被用作分子标签来鉴定细菌种类[15]。将提取的微生物基因组DNA作为模板,16S rDNA V3~V4区序列扩增出电泳图见图2,条带清晰,满足后续测序实验的要求。
2.2 测序序列与OTU数量分析
采用Illumina Miseq高通量测序方法对梅干菜半成品、梅干菜成品和腌制雪菜成品等不同样本进行测序,获得总数为236906条有效序列的样本原始序列,经过质控筛选及软件优化后,三组样本的有效序列为192412条,有效序列数目可满足后续实验的需求。进而对样本序列进行聚类分析,在97%相似水平下的OTU生物信息统计发现,各处理组得到的OTU数量范围为1315~3021(表1)。
OTU样本分布韦恩图可以直观地看出梅干菜半成品、梅干菜成品和腌制雪菜成品样本的OTU数目组成相似性及重叠(图3)。梅干菜半成品、梅干菜成品和腌制雪菜含有的OTU分别为2415、3021和1315个,由此可见在同一扩增区域下,梅干菜成品样本OTU数量多于梅干菜半成品和腌制雪菜成品样本,说明梅干菜成品样本中含有丰富的微生物,而腌制雪菜成品样本中的微生物多样性较少。
2.3 Alpha多样性分析
通过Alpha多样性分析可以反映微生物群落的丰度和多样性。Coverage指数代表每个样品的文库覆盖率,覆盖率达到95%以上就表明文库覆盖率高,未测出概率低,样本序列科学合理,表明测序结果代表样本中微生物的真实情况[16]。由表2可知,梅干菜半成品、梅干菜成品和腌制雪菜成品的Coverage指数均在0.97及以上,说明测序对样本中微生物的覆盖率高,测序结果合理,可满足样本微生物菌群结构和多样性分析的需要[17]。同时,Shannon-Wiener曲线变化趋势也逐渐平缓(图4),说明梅干菜半成品、梅干菜成品和腌制雪菜成品测序数据量合理,能够反映样本中绝大多数的菌群信息。上述现象表明,随着测序量增加,Shannon指数变化趋于平缓,三组样本中菌群多样性不会发生明显变化,微生物分析结果充分。总体来看梅干菜半成品、梅干菜成品和腌制雪菜成品的曲线纵坐标宽度,发现梅干菜成品中微生物物种组成最丰富,其次是梅干菜半成品,三组样本中腌制雪菜成品微生物物种组成数量最低。

图 3 OTU样本分布韦恩图Fig.3 Venn diagram for the operational taxonomic unit distribution of different samples
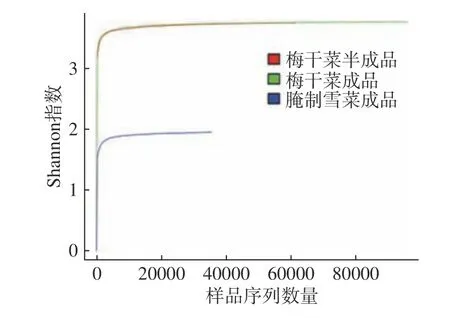
图 4 不同样本Shannon-Wiener曲线分析Fig.4 Shannon-Wiener curves of different sample
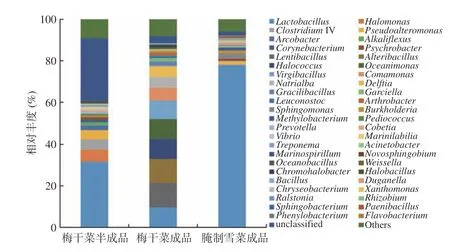
图 5 Genus水平所有样本群落结构分布图Fig.5 Distribution map of the community structure of all samples at genus level
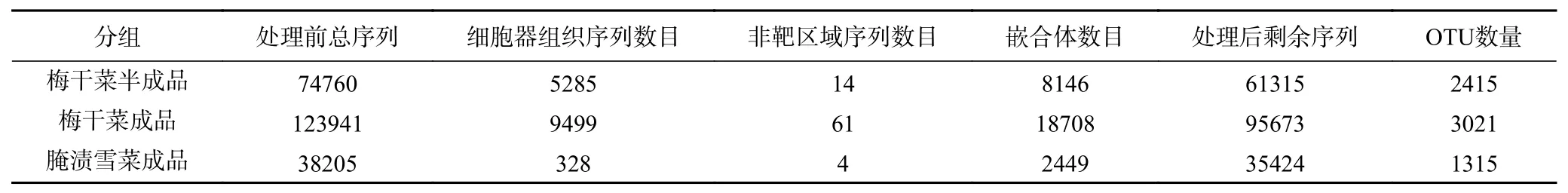
表 1 各样本有效序列与OTU数量Table 1 Valid sequence and OTU numbers of different samples

表 2 不同样本Alpha多样性指数Table 2 Alpha diversity index of different sample
ACE和Chao1指数是样本菌群丰度的表征,数值越高表明菌群丰富度越高[14]。由表2可知,梅干菜成品样本的ACE和Chao1指数最高,其次是梅干菜半成品样本,腌制雪菜成品样本的ACE和Chao1指数最低,表明三组样本中梅干菜成品的菌群丰度最高,其次是梅干菜半成品,而腌制雪菜成品的菌群丰度最低。
Shannon和Simpson指数与样本中菌群多样性相关,其中Shannon指数与菌群多样性呈正相关,而Simpson指数与菌群多样性呈负相关[14]。由表2可知,梅干菜成品与梅干菜半成品样本的Shannon指数高于腌制雪菜成品,而Simpson指数则是相反的结果,表明梅干菜成品和半成品样本中物种多样性较丰富,其中梅干菜成品菌群多样性稍大于梅干菜半成品,而腌制雪菜成品样本中物种多样性较低。

表 3 各样品菌群在属水平上的相对丰度(%)Table 3 Relative abundance of bacterial flora at the genus level in different sample (%)
2.4 菌群多样性分析
对OTU序列进行分类学分析,获得梅干菜半成品、梅干菜成品和腌制雪菜成品样本在genus分类水平上的菌群组成统计(表3和图5)。由表3可知,梅干菜半成品、梅干菜成品和腌制雪菜成品菌群中相对丰度大于1%的菌属分别有8、13、5属。梅干菜半成品的主要优势菌属是乳杆菌属(Lactobacillus)、嗜盐单胞菌属(Halomonas)、梭状芽孢杆菌属(ClostridiumIV)、假交替单胞菌(Pseudoalteromonas)、弓形杆菌属(Arcobacter)、产碱杆菌属(Alkaliflexus)、棒状杆菌属(Corynebacterium)和嗜冷杆菌属(Psychrobacter);梅干菜成品的主要优势菌属是慢生芽孢杆菌属(Lentibacillus)、交替芽胞杆菌属(Alteribacillus)、嗜盐球菌属(Halococcus)、乳杆菌属(Lactobacillus)、海单胞菌属(Oceanimonas)、枝芽孢菌属(Virgibacillus)、丛毛单胞菌属(Comamonas)、钠白菌属(Natrialba)、代尔夫特菌属(Delftia)、薄壁芽孢杆菌属(Gracilibacillus)、硫酸盐还原菌(Garciella)、明串珠菌属(Leuconostoc)和节杆菌属(Arthrobacter);腌制雪菜成品的主要优势菌属是乳杆菌属(Lactobacillus)、鞘氨醇单胞菌属(Sphingomonas)、假交替单胞菌(Pseudoalteromonas)、伯克霍尔德菌(Burkholderia)和甲基杆菌属(Methylobacterium)。
梅干菜半成品中未鉴定菌属相对丰度较高,这可能归因于梅干菜原料较高的菌群多样性。与梅干菜半成品相比,梅干菜成品中表现出部分菌群增长的趋势,菌群的丰度和多样性均提高,表明成品中的各菌属可能是梅干菜风味形成的主要原因。腌制雪菜样本菌群多样性较低,主要优势菌属为乳杆菌属(Lactobacillus),占菌群总数的78.08%,表明乳杆菌属(Lactobacillus)对腌制雪菜的风味和品质有重要影响。腌制雪菜的制作工艺为泡制,乳酸菌发酵产生乳酸,可能抑制不耐酸微生物的生长繁殖;且发酵环境适合兼性厌氧菌生长,而抑制了好氧菌的生长。
3 讨论
已有研究表明,乳杆菌属(Lactobacillus)在大多数传统发酵蔬菜产品中为绝对优势菌属[18−19]。康建依等[20]研究发现乳杆菌属(Lactobacillus)为发酵蔬菜中后期的主要优势菌属,发酵7 d时所占比例达93.5%。尚雪娇等[12]研究表明,乳杆菌属(Lactobacillus)在梅干菜中占据主要作用,相对丰度为60.5%。陈希等[13]对雪菜低盐腌制过程中微生物群落多样性进行分析,结果发现乳杆菌属(Lactobacillus)相对丰度随腌制时间延长逐渐上升,腌制18 d时达到最大值77.5%。鲍伟等[21]研究发现乳杆菌属(Lactobacillus)在臭冬瓜、臭苋菜梗和酸笋等3种浙江传统发酵蔬菜中相对丰度分别为17.88%、8.17%、6.58%,但在酸茭白中相对丰度较低。此外,郭倩倩等[22]分析了晴隆酸菜不同发酵时期的微生物群落结构和多样性,结果表明魏斯氏菌属(Weissella)、乳杆菌属(Lactobacillus)和明串珠菌属(Leuconostoc)等为主要优势菌属。本实验中,乳杆菌属(Lactobacillus)在梅干菜半成品、梅干菜成品及腌制雪菜成品中相对丰度均较高,分别为31.70%、9.51%、78.08%,同时梅干菜半成品中检出片球菌属(Pediococcus),梅干菜成品中检出明串珠菌属(Leuconostoc)和魏斯氏菌属(Weissella),与前人研究结果相似。
梅干菜半成品鉴定到的其他菌属中,嗜盐单胞菌属(Halomonas)可发酵葡萄糖产酸,并具备还原硝酸盐为亚硝酸盐的能力[23],这可能与梅干菜腌制早期亚硝酸盐含量快速上升有关;梭状芽孢杆菌属(ClostridiumIV)、弓形杆菌属(Arcobacter)、棒状杆菌属(Corynebacterium)、伯克霍尔德菌属(Burkholderia)和弧菌属(Vibrio)的部分种具有致病性[24],产碱杆菌属(Alkaliflexus)和不动杆菌属(Acinetobacter)为条件致病菌[25],嗜冷杆菌属(Psychrobacter)等为常见的腐败菌[26],这些微生物很可能来源于原料种植基地和腌制环境,但在梅干菜成品中相对丰度极低甚至未检出,表明后发酵对梅干菜的菌群结构有显著影响,有利于提高梅干菜的食用安全性。梅干菜成品鉴定到的其他菌属中,慢生芽孢杆菌属(Lentibacillus)、交替芽胞杆菌属(Alteribacillus)、嗜盐球菌属(Halococcus)、海单胞菌属(Oceanimonas)、枝芽孢菌属(Virgibacillus)、薄壁芽孢杆菌属(Gracilibacillus)、盐芽孢杆菌属(Halobacillus)和色盐杆菌属(Chromohalobacter)为嗜盐菌[27],总相对丰度达54.98%,表明嗜盐菌为梅干菜后发酵阶段的绝对优势微生物,其共生和协同作用可能对梅干菜风味的形成有重要贡献。硫酸盐还原菌属(Garciella)能通过异化作用进行硫酸盐还原,在二价铁盐的存在下可生成黑色物质[28],色盐杆菌属(Chromohalobacter)在固体培养基上可形成奶酪状的紫色菌落[29],盐芽孢杆菌属(Halobacillus)能产橙色非水溶性色素[30],这可能是梅干菜在发酵后期呈现黄黑色的重要原因。此外,丛毛单胞菌属(Comamonas)、钠白菌属(Natrialba)、代尔夫特菌属(Delftia)和节杆菌属(Arthrobacter)相对丰度均在1%以上,但目前关于上述微生物的生理生化特性研究报道较少,有待进一步研究。腌制雪菜成品鉴定到的其他菌属中,鞘氨醇单胞菌属(Sphingomonas)、产吲哚金黄杆菌属(Chryseobacterium)、黄单胞菌属(Xanthomonas)和黄杆菌属(Flavobacterium)均可产生黄色色素物质[31],甲基杆菌属(Methylobacterium)可在液体培养基中形成粉色表面菌环或菌膜[31],这可能与腌制雪菜呈现金黄色感官品质有关。
不同腌制方式、腌制阶段和不同地区的发酵蔬菜产品优势微生物菌属也有较大差异。尚雪娇等[12]对恩施地区梅干菜细菌多样性进行了研究,发现其主要优势菌属为乳杆菌属(Lactobacillus)、假单胞菌属(Pseudomonas)、葡萄球菌属(Staphylococcus)、魏斯氏菌属(Weissella)、鞘脂单胞菌属(Sphingomonas)、嗜冷杆菌属(Psychrobacter)、四联球菌属(Tetragenococcus)和棒状杆菌属(Corynebacterium)。陈希等[13]研究了宁波地区雪菜低盐腌制过程中微生物群落多样性,结果表明腌制初期主要优势菌群为肠杆菌属(Enterobacter)和欧文氏菌属(Erwinia),发酵中后期为乳杆菌属(Lactobacillus plantarum)。谷懿寰等[19]分析了海南7个地区的30份不同种类传统发酵竹笋、小西瓜、白菜和豆角样品中微生物多样性,发现乳杆菌属(Lactobacillus)、肠球菌属(Enterococcus)和片球菌属(Pediococcus claussen)为主要优势菌属。本实验中,梅干菜半成品、梅干菜成品和腌制雪菜成品中相对丰度大于1%的菌属分别为8、13、5属,其中梅干菜成品中菌群结构较复杂,菌群多样性和相对丰度均较高,这与前人在传统腌制发酵蔬菜中鉴定到的微生物菌属差异较大。以上结果表明,不同地区和环境条件可能对传统发酵蔬菜产品的微生物多样性有较大影响。干法腌制比盐水泡制发酵蔬菜产品的微生物多样性要高,这可能与盐水泡制不利于好氧菌的生长,同时乳杆菌属发酵产酸后对其他菌属有抑制作用有关。
4 结论
Illumina Miseq高通量测序分析结果显示,梅干菜半成品、梅干菜成品和腌制雪菜成品特有的OTU分别为2415、3021和1315个,表明干法腌制比盐水泡制蔬菜产品的菌群结构和多样性更复杂。
梅干菜半成品、梅干菜成品和腌制雪菜成品菌群中相对丰度大于1%的菌属分别有8、13、5属,各种菌属可能对传统腌制蔬菜风味的形成发挥协同作用。
乳杆菌属(Lactobacillus)在梅干菜半成品、梅干菜成品和腌制雪菜成品3种产品中相对丰度均较高,分别为31.70%、9.51%、78.08%,是传统发酵蔬菜的主要优势菌属。嗜盐菌在梅干菜成品中相对丰度达54.98%,为梅干菜后发酵阶段的绝对优势微生物。硫酸盐还原菌属(Garciella)、色盐杆菌属(Chromohalobacter)和盐芽孢杆菌属(Halobacillus)可能与梅干菜色泽形成有关。鞘氨醇单胞菌属(Sphingomonas)、甲基杆菌属(Methylobacterium)、产吲哚金黄杆菌属(Chryseobacterium)、黄单胞菌属(Xanthomonas)和黄杆菌属(Flavobacterium)可能在腌制雪菜色泽形成过程中发挥重要作用。
