外源硒处理的花生芽蛋白中五种硒形态分析
2021-09-26邵志颖
薛 梅 邵志颖 李 彭
(南京财经大学食品科学与工程学院;江苏省粮油品质控制及加工技术重点实验室;江苏省现代粮食流通与安全协同创新中心,南京 210023)
花生是我国主要的经济作物和油料作物,种植面积广泛。花生营养价值较高,含有易被人体消化吸收的高质量蛋白和丰富的不饱和脂肪酸,其中含有8种人体必需氨基酸,亚油酸和油酸占总脂肪含量的80%左右[1]。此外,花生中的天然多酚物质白藜芦醇,具有抑菌、防癌、降血脂、抗氧化等保健功能,被科学界广泛关注[2]。经常食用花生可以改善脑血管功能,延缓衰老,增强记忆,被人们称为“长生果”[3,4]。同时,花生具有很强的富硒能力,它可以将70%以上从培养基中吸收的无机硒转化为有机硒[5],有关花生施用硒元素达到富硒效果以及富硒花生中硒形态的研究已有广泛报导[6,7]。花生种子发芽过程中,贮藏物质在水解酶的催化作用下被分解成小分子化合物,其营养物质更易于吸收。詹玉婷等[8]发现,与未发芽的花生相比,花生芽主要营养成分中的游离氨基酸和维生素C含量显著升高,粗脂肪和可溶性蛋白含量显著减少,活性成分白藜芦醇的含量也显著增加。花生芽作为一种新型芽菜品种,具有营养易吸收、风味独特、经济价值高、生产周期短、无污染等特点,备受中国消费者的青睐。花生芽也被称为“长生菜”,除了含有易于被人体吸收的脂肪、植物蛋白等成分,还含有多种维生素和微量元素[9]。此外,花生芽菜中也含有丰富的白藜芦醇,同样具有抗氧化、抗肿瘤、抗癌等保健功效[10]。
硒是人体必需的微量元素之一,是人体部分抗氧化酶和硒蛋白的重要组成部分[11]。该元素可以保护机体不受由自由基引起的氧化性损伤的损害,具有防癌、提高人体免疫力和抗衰老等功效[12]。硒元素无法在体内合成,人体所需的硒元素只能从食物中摄取。在我国,除了湖北恩施等少数地区外,72%的地区都属于贫硒地带,全国约三分之二的人口存在不同程度的硒摄入不足[13],因此,补硒对于提高我国人民身体素质显得特别重要。由于无机硒不易被吸收,并且易对人体造成毒害[14],因此经植物转化后的硒是人体摄入硒的重要来源,目前常采用土壤施硒和喷硒的方式,使无机硒转化为有机硒,通过农作物进入食物链,以供人体利用[15]。然而,土壤施硒很容易造成土壤和环境污染,富硒花生芽的生产可以通过硒溶液浸泡,在无土条件下完成,更加环保。花生中含有丰富的优质蛋白,花生芽生长周期短,加工简单,营养成分更易吸收,通过花生芽来补充膳食中的硒元素是一种一举两得的方法。近年来,有学者研究了多种蔬菜芽菜富硒后的形态种类及变化[16],Wang等[17]研究了花生芽的富硒机理,发现通过花生幼苗生物转化作用富集将无机硒转化为有机硒是培育富硒花生芽的有效手段,其富硒的机理跟花生幼苗的抗氧化系统和次生代谢均存在密切联系。但对花生芽富硒过程中的硒形态及其分布特征未见报道。本研究通过用Na2SeO3浸泡花生种子,使得无机硒在花生发芽过程中转化为有机硒,观察不同外源硒水平对花生芽生长状况的影响,分析花生芽不同部位无机硒和有机硒的赋存形态,以及不同形态的硒水平。旨在为富硒花生芽的培养和功能开发理论依据,也为花生这一优质营养资源的深度开发和利用提供参考。
1 材料与方法
1.1 材料与试剂
硒标准液(1 000 μg/mL)、硒代甲硫氨酸(SeMet,99%)、硒代胱氨酸(SeCys2,98%)、甲基硒代半胱氨酸(MeSeCys,95%)、亚硒酸钠(Na2SeO3,98%)、硒酸钠(Na2SeO4,98%)、α-淀粉酶、蛋白酶XIV(≥2 500 U/mg)、甲醇(色谱纯)、硝酸、磷酸、草酸、氢氧化钠、次氯酸钠、柠檬酸、磷酸二氢钠(均为优级纯)。
1.2 仪器与设备
MARS6微波消解仪,7700x电感耦合等离子体质谱仪,1260系列高效液相色谱仪,人工气候箱,64R冷冻离心机。
1.3 富硒花生芽发芽工艺
花生种子经筛选、除杂,取100 g成熟饱满均匀的颗粒,用1% NaClO溶液浸泡5 min,蒸馏水冲洗后,转移至已消毒烧杯中。平均分成5份,加入100 mL不同浓度的Na2SeO3溶液(浓度分别为 0、10、20、30和40 mg/L),置于培养箱中25 ℃恒温浸泡6 h后取出、超纯水冲洗、沥干,均匀铺在铺有2层纱布的培养皿(Ф=30 cm)中,均匀喷洒去离子水30 mL,置于人工气候箱中培养,温度为32 ℃,湿度为70%,每12 h喷淋20 mL蒸馏水,120 h后停止发芽。花生芽经超纯水冲洗后,将子叶、胚轴、胚根三部分分开,分别冷冻干燥,干燥后的样品用粉碎机粉碎,过40目筛,冷藏备用。
1.4 芽长、芽粗和发芽率
芽长和芽粗:随机选取20粒发芽花生种子,用最小刻度为0.1 cm 的软尺对花生芽的长度进行测量;用游标卡尺对芽粗进行测量。
发芽率:按GB/T 3543.4—1995《农作物种子检验规程:发芽试验》,随机取100粒种子进行花生富硒发芽试验,统计发芽率。
1.5 标准曲线的制备
Se元素标曲的制备:将Se元素标准工作溶液以1%硝酸-超纯水稀释,配制成0.0、1.0、5.0、10.0、20.0、50.0、100.0 μg/L,ICP-MS调谐合格后,将标准溶液引入仪器,测定Se元素和内标元素的信号响应值,以Se元素的浓度为横坐标,待测元素与内标元素响应信号值的比值为纵坐标,绘制标准曲线。
硒形态标曲的制备:将5种硒形态标准品混合并逐级稀释,配置成浓度为0.0、1.0、5.0、10.0、20.0、50.0 μg/L的混合标准工作液,以五种硒形态峰面积-浓度做图,得到标准曲线。
1.6 样品中硒蛋白的提取
准确称取10 g冻干样品,加入30 mL丙酮搅拌2 h,4 000 r/min离心10 min,弃上清,重复操作3次,将沉淀干燥后得到脱脂样品。取脱脂后的样品0.2 g,加入5 mL α-淀粉酶溶液(26.48 mg/mL),混合溶液在37 ℃水浴中超声波(40 kHz)30 min,37 ℃下水浴振荡2 h。再加入2 mL蛋白酶XIV溶液(7.85 mg/mL)于提取液中,在45 ℃水浴中超声波30 min,然后45 ℃下水浴振荡2 h提取样品中硒的化合物。提取液在5 000 r/min下离心20 min,取上清液过孔径为5 ku的超速离心膜,除去提取液中可能会干扰色谱分析的大分子杂质,经0.45 μm过滤膜后上机分析。
1.7 ICP-MS测总硒含量
称取0.3 g样品于消解罐中,加入5 mL HNO3和1 mL H2O2进行微波消解,微波消解程序见表1。消解结束后,冷却,打开消解罐,样品溶液转移至10 mL容量瓶中,用超纯水洗涤3次,合并洗涤液,再用超纯水定容,摇匀,同样方法做试剂空白试验。分别测定花生芽冻干样品和硒蛋白样品中的硒含量。

表1 微波消解程序
用ICP-MS测定样品中总硒含量,ICP-MS仪器参数见表2。

表2 HPLC-ICP-MS分析条件
1.8 花生发芽过程中不同部位消化后硒形态含量测定
硒形态的测定采用HPLC-ICP-MS。ICP-MS和HPLC的工作参数见表2。
硒形态含量计算公式:
式中:X为试样中每种硒形态的含量/mg/kg;C为样品溶液中每种硒形态的浓度/μg/L;C0为空白溶液中每种硒形态的浓度/μg/L;V为样品溶液最终定容体积/mL;M为试样质量/g;f为稀释倍数,计算结果保留两位有效数字。
1.9 数据处理
实验进行5次重复,每次重复6个平行。采用Origin 8.5对数据进行处理作图,用SPSS 22.0软件对结果进行分析,多重比较采用邓肯氏新复极差法(P<0.05)。
2 结果与讨论
2.1 外源硒对花生芽生长情况的影响
由图1可见,随着Na2SeO3浓度的升高,花生芽长、芽粗和发芽率均呈下降趋势。当Na2SeO3浓度为30 mg/L时,芽长开始受到抑制,降低为62.5 mm,显著低于对照和10 mg/L组。当Na2SeO3浓度升高至40 mg/L时,花生芽生长进一步受到抑制,芽长低至58.8 mm。此外,外源硒浓度低于30 mg/L时,各处理组芽粗没有显著差异,当浓度为40 mg/L时,芽粗受到抑制,为5.29 mm,显著低于对照、10 mg/L和20 mg/L组。发芽率是确定作物播种量的重要依据,也是鉴定种子质量好坏的主要标志[18]。随着Na2SeO3浓度的升高,花生发芽率呈现下降趋势。未经硒处理的花生发芽率最高,为96.7%,Na2SeO3浓度低于20 mg/L时,花生发芽率没有显著变化。随着外源硒浓度继续升高,发芽率显著降低。综上指标可见,高浓度外源硒对花生芽的生长具有明显的抑制作用。胡婷等[19]研究了外源硒对绿豆发芽情况的影响,发现绿豆在Na2SeO3浓度为0.5~5.0 mg/L时发芽率达到最高100.0%,Na2SeO3浓高于10.0 mg/L时,种子发芽率低于对照组。彭诚等[20]研究发现,白菜种子发芽率在较高质量浓度Na2SeO3溶液下表现出抑制作用。王玉凤等[21]通过研究番茄种子发现,在外源硒质量浓度为4.0 mg/L时,其发芽率最大,种子萌发第7天时的芽鲜质量、根鲜质量、芽长、根长、芽干质量和根干质量也达到最大值;而高质量浓度的硒处理(>4.0 mg/L)则表现为一定的抑制作用。综合以上研究,高浓度外源硒会不同程度抑制粮食和蔬菜种子的发芽,这与本实验中花生种子发芽率研究结果一致。
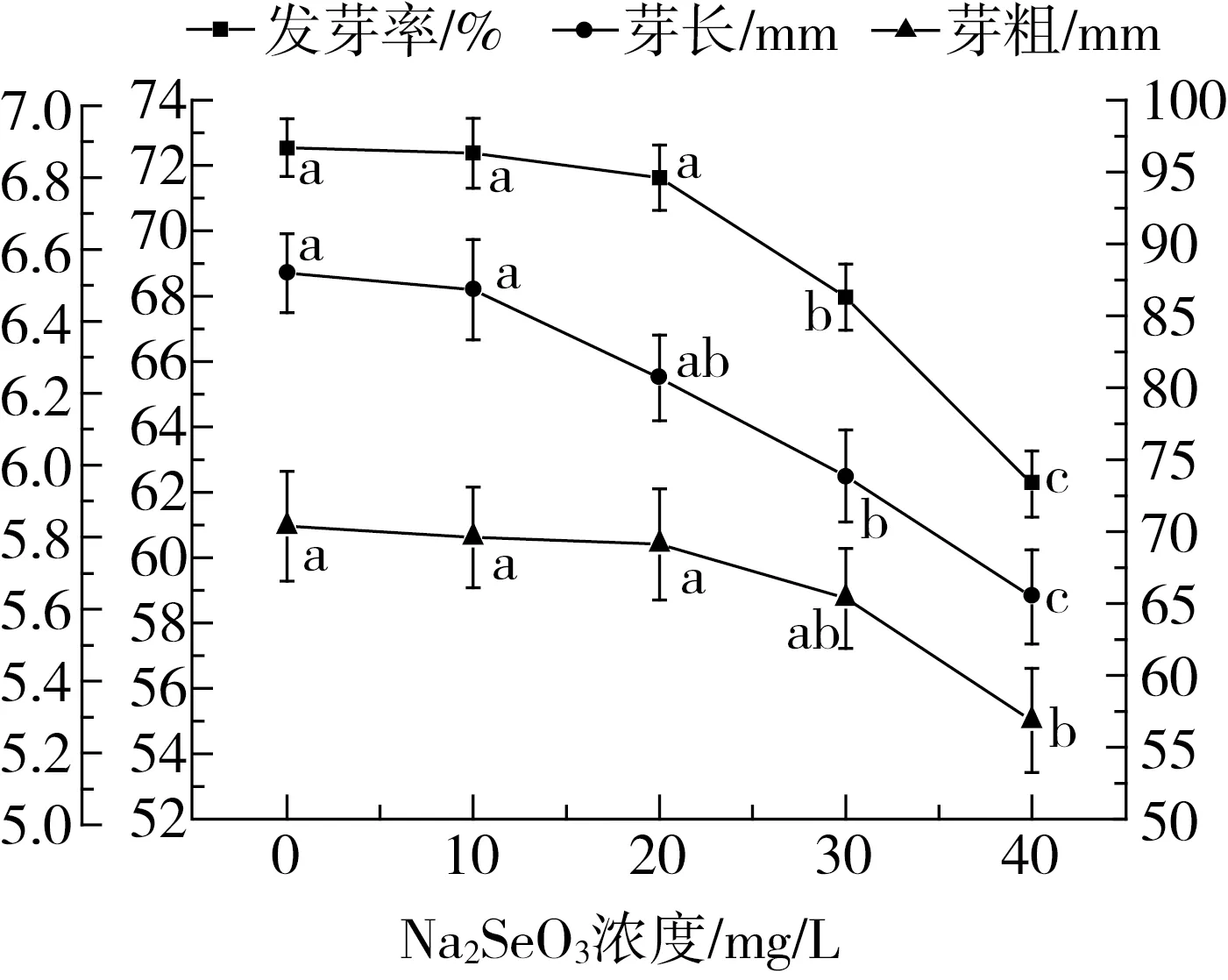
注:同一系列不同字母表示差异显著(P<0.05)。图1 不同浓度硒浸泡的花生发芽5 d后的芽长、芽粗和发芽率
2.2 色谱和质谱条件优化
2.2.1 五种硒形态色谱分离条件优化
本实验选用Hamilton PRP X-100(250 mm×4.1 mm,10 μm)阴离子交换柱进行5种硒形态化合物的分离,比较了磷酸二氢铵和柠檬酸2种流动相,结果表明,用40 mmol/L磷酸二氢铵作为流动相时,所测5种硒形态化合物并不能良好的分开,MeSeCys和SeIV形态峰出现重叠。选用6 mmol/L柠檬酸所为流动相时,能够实现5种不同的硒形态化合物的完全分离,此外,柠檬酸作为流动相,无毒、成分单一易于配置,因此选用柠檬酸作为流动相。有学者在进行硒形态分离时发现,在不同的酸度条件下,硒的不同形态会以阳离子、阴离子或者两性离子的形式存在,因此,流动相pH值是影响5种硒形态的保留时间和分离效果的主要因素[22]。本实验比较了柠檬酸pH为4.5、5.0和5.5时的分离效果,结果表明,当pH为4.5时,SeMet和SeIV形态峰出现重叠,不能完全分开;当pH值达到5.0时,SeIV形态峰保留时间缩短,与SeMet峰完全分开,实现了5种硒形态化合物的完全分离,同时,SeVI保留时间明显缩短,提高了分离效率;当pH值增大至5.5时,MeSeCys和SeIV峰无法完全分开,而SeMet和SeVI峰几乎重叠。通过优化梯度洗脱发现,当柠檬酸浓度逐渐增大时,SeVI保留时间缩短,当浓度为15 mmol/L时,既可以缩短SeVI的保留时间,又可以实现5种硒形态化合物的完全分离,当浓度大于15 mmol/L时,SeMet和SeVI形态峰重叠。综合考虑保留时间和分离度,优化5种硒形态化合物混合标准溶液色谱图见图3。
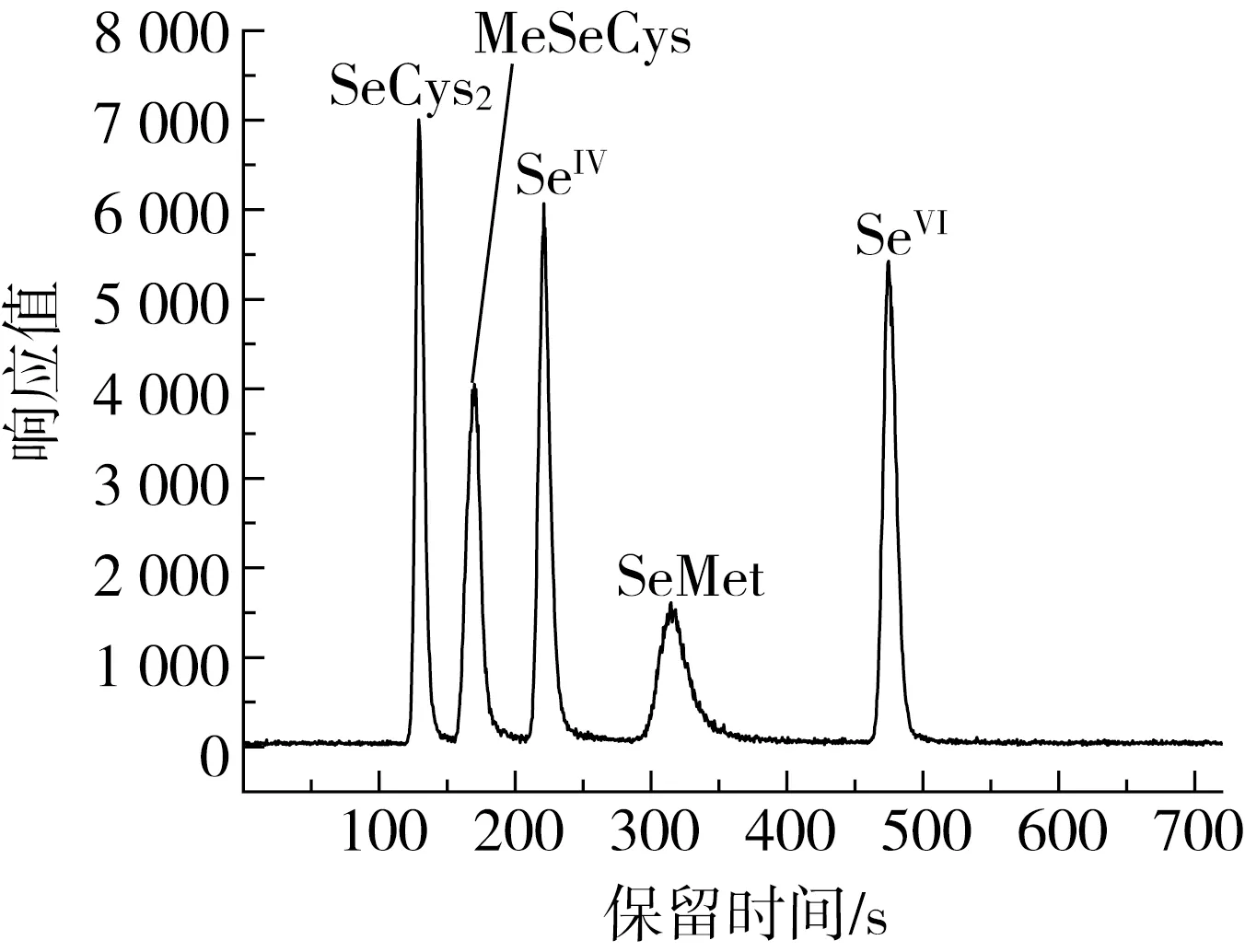
图2 五种硒形态的HPLC-ICP-MS色谱图
2.2.2 质谱条件优化
质谱干扰包括同量异位素重叠干扰、多原子离子干扰、难熔氧化物离子干扰和双电荷离子干扰[23]。Se元素的同量异位素中,78Se和80Se会受到40Ar38Ar+、40Ar40Ar+、HBr+等多原子离子干扰,背景值太高不能获得很好的灵敏度。本研究采用高氦分析模式(HEHe)消除多原子离子干扰,并对仪器条件进行优化,通过7Li、89Y和205Tl矫正仪器,使其分辨率在0.65~0.080 u,氧化物(CeO+/Ce+)≤3%,双电荷≤2%,确保消除双电荷、同位素、氧化物、氢化物及多原子离子等物质对结果的干扰。同时,为了消除非质谱干扰,降低基体效应的影响,减少分析信号的漂移,本研究通过内标加入进行定量校正。基于测定同位素丰度最大原则和谱线干扰最少原则,选择72Ge作为内标,对78Se进行定量测定。
2.3 外源硒对花生芽不同部位硒含量的影响
Se元素标准曲线为y=2.403x+0.999 1,校正曲线线性相关系数为0.999 6,元素的检出限为0.014 μg/kg。分别测得花生芽各部位总硒含量见表3。
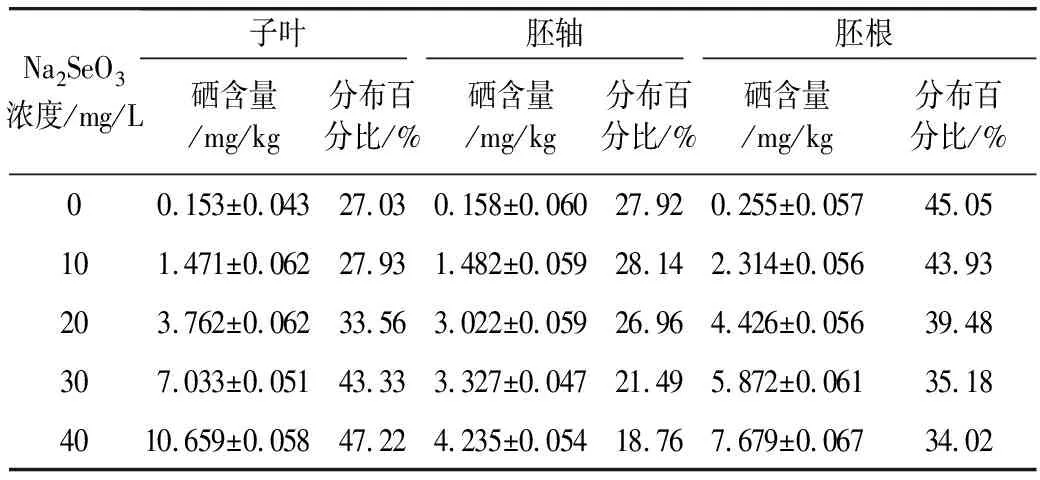
表3 花生芽各部位总硒含量
结果表明,随着Na2SeO3浓度的升高,各部位硒含量均显著增加。由各部位硒的分布百分比可见,当Na2SeO3浓度低于10 mg/L,花生芽各部位富硒能力为胚根>胚轴>子叶;当Na2SeO3浓度为20 mg/L,各部位富硒能力为胚根>子叶>胚轴,当Na2SeO3浓度升高至30和40 mg/L,花生芽各部位富硒能力为子叶>胚根>胚轴。Wang等[17]研究发现,Na2SeO3浓度低于10 mg/L时,花生幼苗子叶中的总硒含量显著低于胚轴和胚根,表明幼苗的根部和下胚轴对硒的富集能力强于子叶。该研究与本实验结果一致,这可能跟幼苗通过根部吸收和转化无机硒有密切关系。然而,随着外源硒浓度升高,子叶中的硒含量比例逐渐升高,胚根和胚轴中硒含量比例逐渐降低,说明高浓度的外源硒抑制了花生发芽过程中硒元素由子叶向胚轴和胚根的转移。
2.4 外源硒对花生芽各部位硒形态及其含量的影响
2.4.1 HPLC-ICP-MS测定硒形态的线性方程和检出限
按仪器工作条件对5种硒形态化合物标准溶液系列进行测定,将样品处理过程的空白溶液连续测定10次,以标准偏差的3倍计算仪器检出限。5种硒形态的保留时间、线性方程和检出限,见表4。在0~50 μg/L范围内,各硒形态校正曲线呈良好的线性关系,线性相关系数为0.999 1~0.999 7,检出限为0.4~2.2 μg/kg。结果表明,该方法具有较高的灵敏度。

表4 五种硒形态的保留时间、线性方程和检出限
2.4.2 花生芽蛋白中硒形态及含量
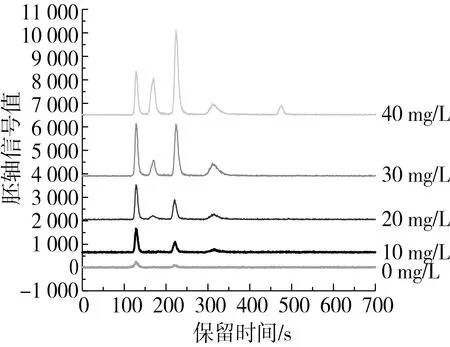
不同浓度外源硒浸泡的花生发芽5 d后,运用HPLC-ICP-MS法分别测定子叶、胚轴、胚根中的硒形态及分布特征,测定图谱及数据结果分别见图3和表5。

表5 花生芽蛋白中五种硒形态含量和百分比

图3 花生芽子叶、胚轴和胚根中的硒形态HPLC-ICP-MS色谱图
结果表明,当Na2SeO3浓度低于10 mg/L时,3个部位中的硒形态均为SeCys2、SeIV和SeMet 3种,其中SeCys2含量最高,子叶中有机硒百分比为63.72%和63.05%,胚轴和胚根中有机硒则达到75.42%~81.14%,说明在该浓度范围,花生经过5d发芽,大多数无机硒转化为有机硒,并且,胚轴和胚根部位有机硒转换能力高于子叶。当Na2SeO3浓度为20 mg/L时,胚轴和胚根中,除了SeCys2、SeIV和SeMet 3种硒形态外,出现了MeSeCys,含量较低;该浓度下子叶有机硒降低为57.46%,胚轴和胚根中有机硒占比为73.98%和75.21%,说明有机硒转化率胚根>胚轴>子叶。当Na2SeO3浓度为30 mg/L时,3个部位中的硒形态均为SeCys2、MeSeCys、SeIV和SeMet 4种,其中,子叶部位的SeIV含量最高,占49.01%,显著高于低浓度处理组,MeSeCys含量最少,占2.78%;胚根和胚轴中4种硒形态含量均为SeCys2>SeIV>SeMet>MeSeCys,且有机硒占比显著高于子叶,说明胚轴和胚根的有机硒转化能力高于子叶。当Na2SeO3浓度为40 mg/L时,3个部位均出现5种硒形态,SeVI只有在该浓度下出现,含量最低,占总硒形态的2.97%~4.08%,SeVI的出现可能是大量的SeIV长时间未被吸收而氧化后的结果;各部位有机硒占比为胚轴>胚根>子叶。此外,外源硒浓度为10和20 mg/L时,各部位有机硒所占比例较高,且花生芽生长情况不受影响,适宜作为食用和生产研究参考。

五种硒形态中,有机硒对人体健康均具有积极作用。SeMet被认为是硒的最易吸收的形式,能显著抑制前列腺癌的患病率;SeCys2最常见于含硒蛋白质,例如谷胱甘肽过氧化物酶、碘甲状腺原氨酸脱碘酶和硫氧还蛋白还原酶,具有抗氧化和调节甲状腺功能的作用[24];MeSeCys具有化学预防作用[25]。随着外源硒浓度升高,SeCys2在各部位中占比逐渐降低,MeSeCys、SeIV和SeVI逐渐增加,SeMet变化没有规律,各部位形态组成的变化说明无机硒向有机硒的转化方式发生了变化。Bodnar等[16]研究了SeVI在西蓝花、萝卜、红甘蓝、芥菜等6种芽菜中的转化,发现Na2SeO3浸泡的蔬菜种子发芽后,MeSeCys含量增加,该结果与本实验结果一致。此外,花生芽中总有机硒占比随着外源硒浓度升高,由75.18%降至50.48%,富硒能力受到显著抑制。无论是子叶还是胚轴与胚根,都能检测到无机硒和有机硒两种形态同时存在,这说明亚硒酸钠被花生种子吸收后,一部分以无机硒的形式输送到花生芽的不同部位后再转化为有机硒,也可能有一部分无机硒在子叶转化成有机硒后再运输到其他部位。该结果与先前学者报导结果一致[26,27]。但也有可能涉及植物吸收硒元素以后在植物体内存在无机硒与有机硒的互相转化过程[28]。
3 结论
无机硒可以通过花生幼苗的生物转化作用转化为有机硒富集在花生芽各个部位,是培育富硒花生芽的有效手段。随着Na2SeO3浸泡浓度的升高,花生芽长、芽粗和发芽率均呈降低趋势,高浓度外源硒抑制花生芽生长;高浓度的外源硒抑制了花生发芽过程中硒元素由子叶向胚轴和胚根的转移;未经硒浸泡的花生发芽后,三个部位均检测到三种硒形态,其中SeCys2占比最高,随着硒浓度升高,各部位硒形态种类增加,无机硒(SeIV和SeVI)占比逐渐升高,SeCys2和MeSeCys占比逐渐降低,SeMet占比没有规律性变化,各部位有机硒总量占比降低,无机硒向有机硒转化率收到抑制。
