悬铃木幼苗的初生维管系统演化结构研究
2021-03-27王桂文张立秋王世东由继红刘丹丹陆静梅
王桂文,高 伟,张立秋,王世东,李 岩,由继红,刘丹丹,朱 军,陆静梅
(1.东北师范大学生命科学学院,吉林 长春 130024;2.公安部物证鉴定中心,北京 100038)
悬铃木为悬铃木科(Platanaceae)悬铃木属(PlatanusLinn.)植物.悬铃木科是一个只有1属约11种的小科[1].我国南北各地栽培悬铃木科植物约3种,多作为行道树和庭荫树[2],即一球悬铃木(PlatanusoccidentalisL.)、二球悬铃木(PlatanusacerifoliaWilld.)和三球悬铃木(PlatanusorientalisL.).其中二球悬铃木是三球悬铃木和一球悬铃木的杂交种,1640年在英国伦敦育成,后引种到世界各大城市,广泛栽培[3].每年5月份左右,悬铃木老果脱落导致大量的果毛飘散,而伴随着新生雄花的开放花粉大量散落,造成环境污染的同时,对人的皮肤和呼吸系统具有一定刺激作用[4-5].近年来,国内外研究者们对悬铃木的研究多集中在如何解决其花粉散落、落果飞毛的问题上,并通过筛选自然突变体植株[6-7]、化学诱导球果不正常发育促使果实脱落[8]、利用嫁接或组织培养的方式培育少球株系[9-11]、诱变育种[12]、多倍体育种[13]、基因工程育种[14-16]等多种手段进行了系统研究.成俊卿等[17]对悬铃木木材进行过解剖学研究,但是,有关悬铃木幼苗的初生维管系统演化结构的研究,鲜见报道.为此,本文对悬铃木幼苗初生维管组织系统进行了研究,以探索其维管演化进程,为证明子叶节区的原始性及其系统演化提供一定的依据.
1 材料与方法
实验材料为采自大连地区的普通二球悬铃木(PlatanusacerifoliaWilld.)种子.室温萌发,至具有1~3片真叶的幼苗.将幼苗切成6 mm小段,置于于FAA固定液中,采用石蜡切片法制成连续切片,厚度10 μm,番红、固绿双重染色,中性树胶封片.在数码生物显微镜下观察、照相.
2 实验结果
二球悬铃木幼苗为子叶出土幼苗,子叶长条线形.根据子叶节区理论[18],可将幼苗分为子叶节区、上胚轴苗区和下胚轴根区3部分.
2.1 子叶节区
二球悬铃木子叶节区较长,一般为8.9~13.8 mm,几乎占据整个幼苗下胚轴的全长.
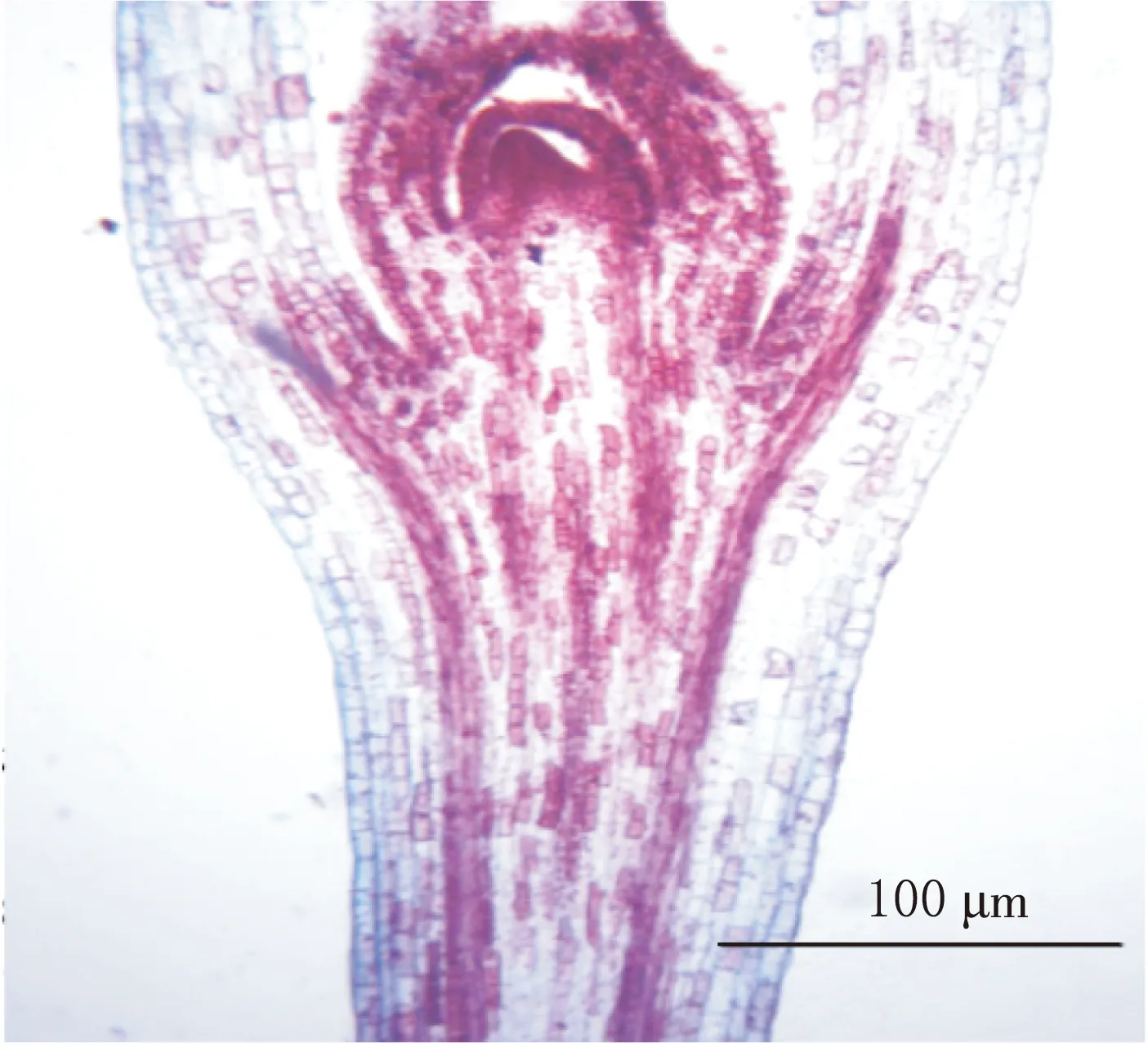
二球悬铃木子叶节区上部长2.9~3.2 mm、直径545~830 μm,至此,子叶节区上部向茎的真中柱过渡的雏形已经基本形成.越靠近苗端时,横切面上韧皮部彼此距离越明显,呈间断性条带状分离排列.在横切面上可见到子叶迹一分为二,形成二叶迹.子叶迹直接与子叶节区上部的管状中柱相连接,既有子叶迹又有子叶隙.被子叶迹隔开的两条形的维管束又分成1~3束,且中央部分被薄壁的髓细胞填充,同时子叶迹向外生长(见图1).

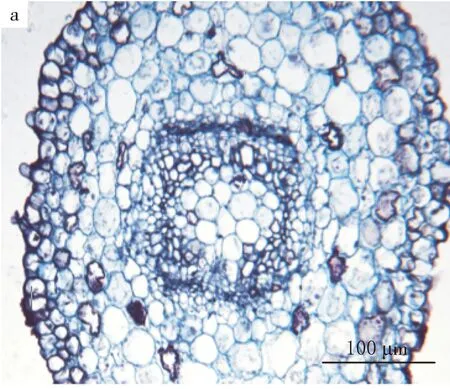
a.二叶迹形成
二球悬铃木子叶节区中部长4.0~4.3 mm,直径531~730 μm.位于子叶节区下部的上方.在这个区域,初生木质部出现了中始式管状中柱阶段.中始式二原型管状中柱的原生木质部位于两端,从原生木质部处一分为二,同时弦向发育,且中央部分被髓细胞占据,其后生木质部各向外侧转约45°,形成椭圆至方形的中柱类型(见图2).
a.子叶节的二叶迹
二球悬铃木子叶节区长1.6~2.0 mm,其横切面直径为430~662μm.中柱类型为中始式二原型单中柱.原生木质部导管直径很小,由环纹和螺纹加厚的导管构成;后生木质部导管直径较大,由梯纹、网纹和孔纹导管构成(见图3—4).
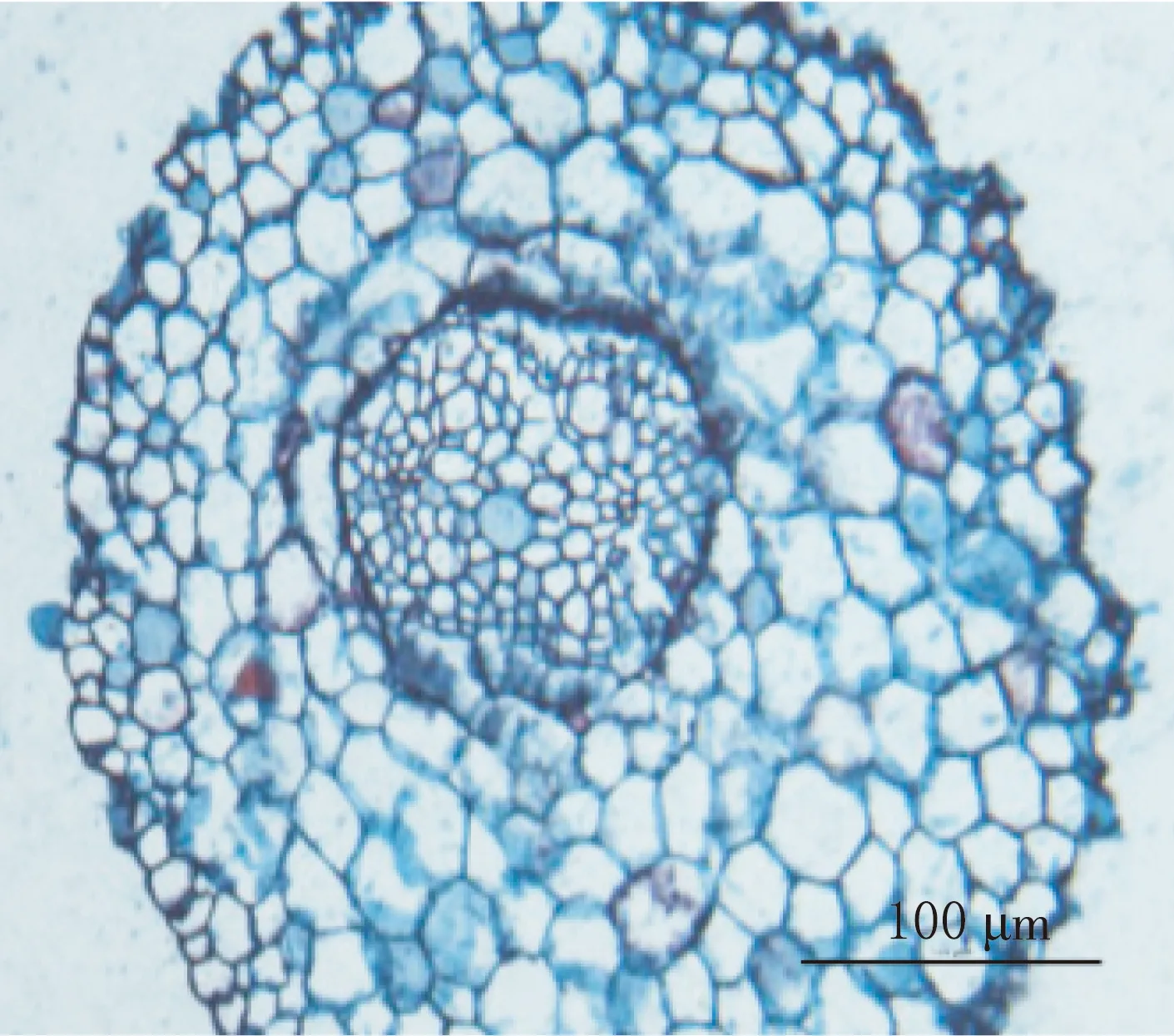
中始式二原型中柱
2.2 上胚轴——苗区
上胚轴苗区属于植物体的轴向后生体,位于子叶节区的上部.上胚轴下连子叶节区上部,上连顶芽,由子叶节区中、上部分化出来的外韧维管束雏形(见图1b)逐渐与茎的内始式真中柱连接(见图5).
茎的内始式中柱由大于6束的外韧维管束组成.子叶双迹清晰可见(见图6a).胚芽位于上胚轴上方.由顶芽发生的叶为三叶迹(见图6b).
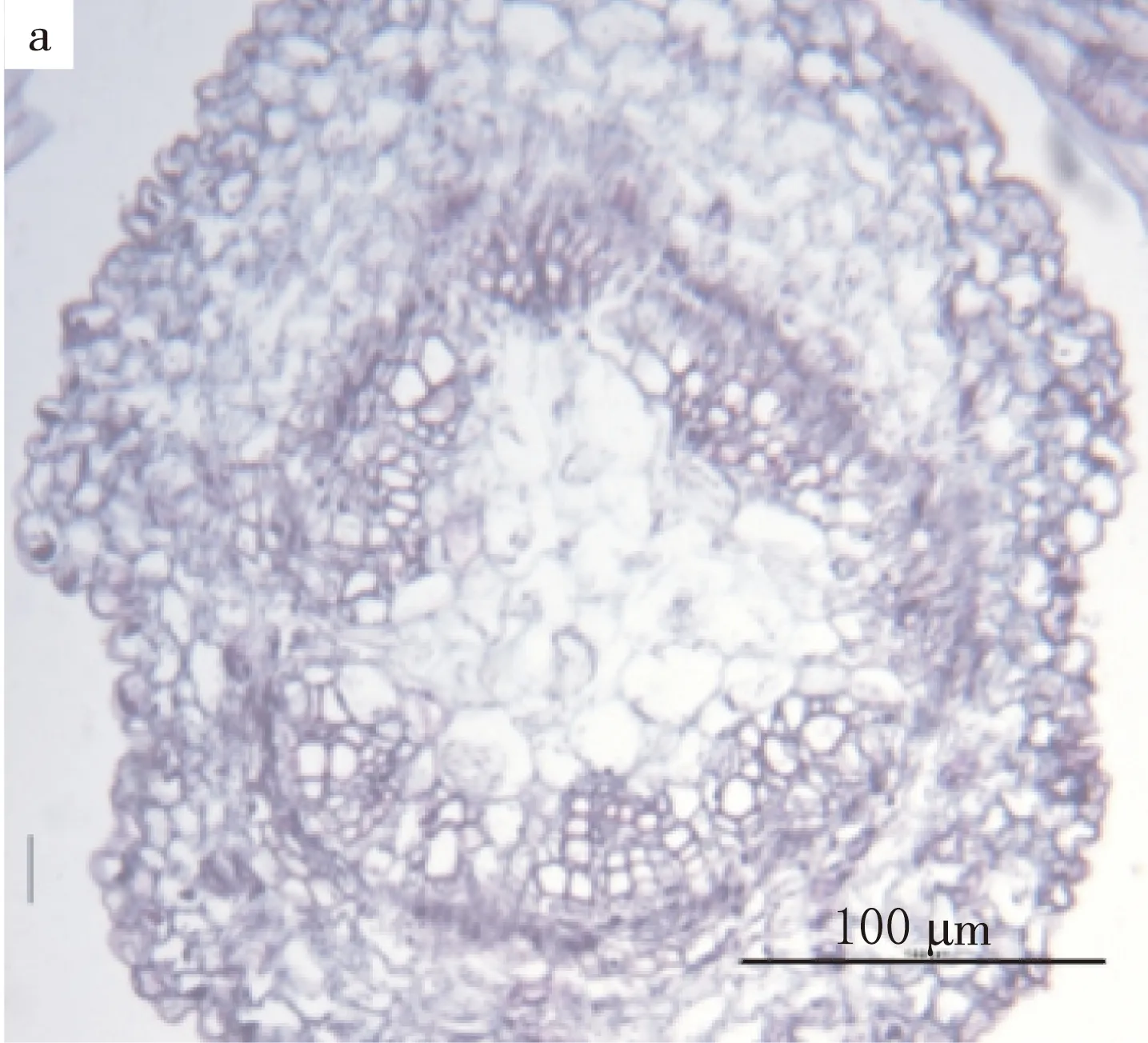
a.子叶分化出外韧维管束
a.子叶的双叶迹
由生长点和数枚幼叶及腋芽原基等结构,包括:茎的生长点、幼叶原基、幼叶和幼茎的初生结构,表皮、皮层和维管柱见图7.
2.3 下胚轴—根区
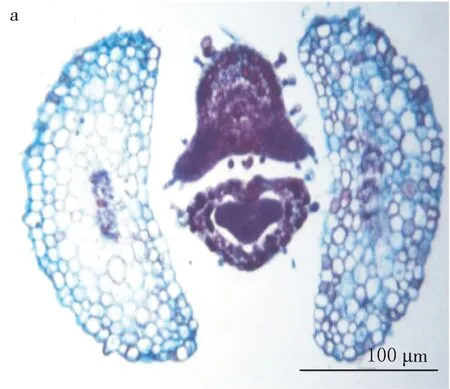
下胚轴根区过渡区位于子叶节区下部的下面,这一阶段极短,为0.88~1.08 mm.后生木质部向轴的中央部位旋转45°,完成了中始式的二原型单中柱向外始式过渡.二球悬铃木的主根为二原型(或真二原假四原)辐射中柱(见图8)
茎的生长点、叶原基和幼叶
3 分析与讨论
3.1 二球悬铃木子叶节区初生维管系统演化过程
根据子叶节区理论[18],子叶节区是种子蕨的孑遗部分,故可称作幼苗的轴向原生体,而分别称为上胚轴苗区与下胚轴根区的均为其幼苗的轴向后生体部分.从系统发育来看,后两者均系由前者所衍生.子叶节区相当于一个等二叉状或复等二叉状分枝的顶枝束,可分成上、中、下三部分.谷安根等[18]已经对毛茛科、葫芦科等双子叶草本植物进行了子叶节区研究,但是对木本植物子叶节区的研究目前尚未报道.
本论文中对二球悬铃木幼苗的研究结果表明,二球悬铃木幼苗的初生维管系统具有较长的子叶节区,几乎占据幼苗下胚轴的全部.通过其子叶横、纵切面观察,根端为外始式二原型(或真二原假四原)辐射中柱.子叶节区下部较为原始,具有中始式二原型单中柱,且这一阶段特别短;中央出现了具髓的中始式二原型管状中柱阶段,且这一阶段较长,髓细胞较大且均由薄壁细胞组成;上部维管束的数目逐渐增多,并且与茎的内始式真中柱极为相似,所以只存在子叶节区茎连接区.上胚轴苗区具有内始式真中柱.由于子叶节区部分中始式管状中柱的存在,为种子植物与真蕨类可能有过具有管状中柱阶段的共同祖先提供了证据,同时又为幼苗初生维管组织从子叶节区分别向根端和苗端演化提供了依据.由于二球悬铃木幼苗子叶节区下部出现中始式二原型单中柱及子叶与营养叶不同迹,充分证明了幼苗子叶节区的原始性.二球悬铃木幼苗的两子叶迹汇合形成长“V”字形.
3.2 子叶迹与营养叶叶迹
子叶节区的子叶为二叶迹,而胚芽起源的营养叶为三叶迹,由此可判断营养叶非子叶节区生成.子叶迹直接与子叶节区上部的管状中柱相连接,既有子叶迹又有子叶隙.
3.3 连接区与过渡区
鉴于子叶节区上部的维管束类型已经与茎的内始式真中柱极为相似并且较短,即子叶节区上部至上胚轴苗区维管束的变化并不是特别激烈,所以特称为子叶节区-茎连接区.子叶节区下部与根之间虽然距离较短,但是变化明显,所以特称为子叶节区-根过渡区.
3.4 拟分生组织
根据子叶节区理论[18],幼苗子叶节区的中、上部分保留有一团与顶端分生组织无关的分生组织性组织(meristematic tissue),即拟分生组织.但是从二球悬铃木幼苗横切面上来看,从中部开始形成具有髓的中始式二原型管状中柱;纵切面上看,中上部的拟分生组织不明显,这可能与选苗期有关,尚需进一步探究.
3.5 子叶节区类型
子叶节区类型可分为5种类型[18],其中顶枝伸长型的特点为:子叶节区颇长,几乎占据整个下胚轴的全长.从系统发育来看,它相当于由数次等二叉分枝的复合顶枝(syntelome)所衍生.其子叶柄稍下部的横切面上,可以观察到1~3或4至多枚子叶迹(偶数多迹).子叶节区下部和子叶节区-根过渡区甚短,其维管组织的向基过渡亦不甚激烈.子叶节区中部特长,其维管组织向顶(子叶)方向的转变十分激烈.在具顶枝生长型子叶节区的幼苗,只存在短的子叶节区-根过渡区和子叶节区-茎连接区.
根据二球悬铃木的初生维管组织特点,其子叶节区类型属于顶枝伸长型.
国内外,有关被子植物幼苗初生维管系统发育的研究,已经积累了大量的资料,研究重点主要集中在初生维管系统的过渡区上.目前,关于植物初生维管系统根、茎过渡方式主要包括3种主要理论:
伊姆斯和麦克唐纳(Eames and MacDaniels,1947)理论,这一学派认为:根和茎是互相连续的结构,共同组成植物体的体轴.在植物幼苗时期的茎和根相接的部分,出现双方各自特征性结构,即根的初生维管组织为间隔排列,木质部为外始式;茎的初生维管组织为内外排列.木质部为内始式的过渡,即:根-茎过渡区.过渡区通常很短,从小于1 mm到2~3 mm,很少达到几厘米.过渡区发生在胚根以上的下胚轴的最基部、中部或上部,终止于子叶节上.在过渡区内,表皮、皮层等是直接连续的,但维管组织要有一个改组和连接的过程.
Katherine Esau理论(1950),主张幼苗可分成根-下胚轴-子叶与上胚轴苗等两个单位, 上胚轴常似一种分离的结构,生在根-下胚轴-子叶的单位上,根只与子叶之间存在维管组织的转换,而与上胚轴(茎)间不存在过渡.就是说下胚轴是根与子叶之间的过渡区,并非是“根-茎过渡区”.过渡区代表一种连接,这并不能认为是介乎组织上有不同排列的两种器官之间的连接,而应看作是维管系统与叶子发育有关系的连接.因此,过渡区的研究必须说明根的维管系统和最早叶迹之间的关系.并以甜菜幼苗为例,说明双子叶植物根和子叶连接的过渡区.
子叶节区(Cotyledon node zone,1990)理论,谷安根等对毛茛科某些属植物幼苗初生维管系统的发育方面做了大量研究,并探讨了毛茛科不同属间的中柱演化关系,在此基础上,创建了子叶节区理论,并就其在被子植物演化形态学上的应用进行了阐述.根据子叶节区理论,将双子叶植物幼苗形态分为上胚轴苗区、子叶节区和下胚轴根区.其中的植物子叶节区是种子蕨的孑遗部分,故可称作幼苗的轴向原生体,而分别称其上胚轴苗区与下胚轴根区均为其幼苗的轴向后生体部分.从系统发育来看,后两者均由前者所衍生.本文验证了谷安根的子叶节区理论.
