军曹鱼(Rachycentron canadum)性腺分化及首周年发育的组织学观察
2021-09-01邝杰华陈刚马骞毛非凡周启苓黄建盛施钢张健东
邝杰华,陈刚,2*,马骞,2*,毛非凡,周启苓,黄建盛,2,施钢,张健东,2
( 1.广东海洋大学 水产学院,广东 湛江 524025;2.南方海洋科学与工程广东省实验室(湛江),广东 湛江 524025)
1 引言
军曹鱼(Rachycentron canadum)隶属鲈形目(Perciformes)、 军 曹 鱼 科 (Rachycentridae)、军 曹 鱼 属(Rachycentron),为广盐暖水性海水鱼类,俗称海鲡、海龙鱼,广泛分布于热带和亚热带海域(东太平洋除外)[1],在南海部分海域亦有少量分布。由于军曹鱼具有生长速率快、易于驯化、抗病力强、肉厚、肉质细腻等特点[2],近年来,其在我国南方沿海地区的养殖发展迅猛,随着人工繁殖和大规模育苗技术的突破,已成为我国极具前景的海水网箱养殖鱼类之一。
性腺发育是鱼类繁殖的前提条件,对鱼类种群的繁衍发展至关重要。硬骨鱼类性腺发育需经历发生、分化及成熟3个阶段,原始生殖细胞(PrimordialGerm Cells,PGCs)迁移至生殖嵴后与体细胞构成性腺原基,随着性腺原基内的PGCs和体细胞进一步分裂增殖,逐渐发育为原始性腺[3]。随后,原始性腺分化出两性特征,而性腺从分化结束到形成成熟精巢或卵巢的整个过程均会表现出明显的阶段性特征,绝大部分还会表现出周期性变化。目前,国内外学者[4-5]主要根据解剖学和细胞学上的特征将鱼类性腺的发育划分为不同分期。军曹鱼为多次产卵类型鱼类,在外观上缺乏性别二态性,难以通过体表特征准确判断其雌雄。近年来,国外学者[6-7]已对部分水域野生军曹鱼种群的繁殖习性和性腺发育特征开展了相关基础研究;许乐乐等[8]通过对性成熟军曹鱼精巢的组织学观察,分析精巢生殖上皮的动态变化,以评估军曹鱼精巢发育状况的周年性变化;Dhanasekar等[9]利用透射电镜技术对军曹鱼精巢中的生精细胞进行观察分析,首次记录了军曹鱼精子超微结构,并详细描述其发育过程;Dutney等[10]发现内分泌干扰化学物质的干扰可导致军曹鱼性激素分泌失衡,部分个体甚至出现雌雄同体的现象。然而,有关军曹鱼原始生殖细胞的起源、迁移和分化及配子发生等方面的研究尚未见报道。
本研究以1~360孵化日龄(days post hatching,dph)的不同发育阶段军曹鱼为研究对象,采用石蜡组织切片和苏木精-伊红(Haematoxylin-Eosin,H.E.)染色法,系统观察并描述了军曹鱼原始性腺的形成、分化特征及精巢和卵巢首周年发育的组织结构变化,相关结果将进一步丰富军曹鱼的繁殖生物学研究内容,有助于阐明其性腺发育规律,为海水鱼的繁育生产提供理论参考。
2 材料与方法
2.1 实验材料及处理
2019年4 月,在广东省湛江市东海岛养殖科技园通过人工繁殖获得军曹鱼初孵仔鱼,从海上网箱中挑选自养的3龄以上成鱼作为亲鱼,在室外水泥池(100 m3)中强化培育,注射 LRH-A3和 HCG 催产,雌鱼注射剂量分别为 LRH-A38 μg/kg,HCG 1 200 IU/kg,雄鱼的注射量为雌鱼的1/2,效应时间为10~12 h。孵化后的仔鱼培育于室内水泥池(5 m×3.5 m×1.5 m)中,于55 dph时将其转移至广东省茂名市电白县某室内养殖基地,饲养于直径为9 m,水深为2.5 m的金属圆形水池内,流水充气,养殖盐度为27.0~30.5,温度为25.0~30.0℃。
实验用鱼均经40 mg/L的丁香酚麻醉,测量形态学指标后进行取样。(1)1~15 dph(5~35 mm):整体取样,每 2 d取 1次样,每次取样 20尾;(2)16~52 dph(40~150 mm):对于性腺未形成的稚鱼直接剪取腹部性腺所在位置,对于性腺已成形的稚幼鱼则解剖取出性腺,每 3~4 d取 1 次样,每次取样 20尾;(3)60~360 dph(21.8~68.5 cm):分别对60 dph、90 dph、120 dph、150 dph、185 dph、210 dph和360 dph体质良好的军曹鱼进行取样,解剖后迅速取出性腺,其中,实验用雌鱼共 26尾,体重为 165.0~5 050.0 g,体长为 22.4~68.5 cm;雄鱼共21尾,体重为160.0~4 225.0 g,体长为21.8~64.5 cm。采集的所有样品均经4%多聚甲醛(Paraformaldehyde,PFA)固定24 h,随后转至70%乙醇中保存。
2.2 实验方法
将已固定好的组织样品经过50%、70%、80%、95%(第1次脱水)、95%(第2次脱水)、100%(第1次脱水)及100%(第2次脱水)乙醇进行梯度脱水,二甲苯透明,石蜡浸透和包埋。用Leica RM2125手动轮转式切片机进行连续切片,切片厚度为 5~6 μm,采用苏木精-伊红染色,最后用中性树胶封片保存,通过Nikon E80i显微镜进行观察,并采集显微图像。参照刘筠[11]对鱼类性腺分期的方法,对不同日龄军曹鱼精巢和卵巢进行发育分期的划分。
3 结果
3.1 原始生殖细胞的特征变化及迁移
3 dph(平均全长为4.5 mm),在鱼体腹部后方靠近生殖孔的位置,可观察到游离的单个原始生殖细胞分布于肠管上方与中肾管侧下方之间的空腔(图1a)。原始生殖细胞呈椭圆形,其直径(7~8 μm)明显大于周围的体细胞,细胞核(3~4 μm)占比大且边界明显,着色较浅,核仁清晰(图1b)。5 dph(平均全长为6.1 mm),原始生殖细胞沿着腹腔背侧的肠系膜迁移至中肾管的正下方,呈梨形,直径增大(图1c)。7 dph(平均全长为8.2 mm)和9 dph(平均全长为10.1 mm),原始生殖细胞均迁移到达贴近肾脏下方的位置,即生殖嵴处,细胞形态无明显变化,仍为单个游离状态(图1d,图1e)。11 dph(平均全长为 12.3 mm),腹膜上皮组织的体细胞开始聚集于生殖嵴处原始生殖细胞的周围(图1f)。15 dph(平均全长为 23.7 mm),原始生殖细胞已被数层体细胞包围,在生殖嵴处形成两个朝向体腔的芽状突起,即原始性腺的雏形,标志着原始生殖细胞胚后迁移的结束(图1g)。

图1 军曹鱼原始生殖细胞的胚后迁移Fig.1 Migration of primordial germ cells of Rachycentron canadum during early development
3.2 原始性腺的分化
18 dph(平均全长为31.5 mm),体细胞与生殖细胞通过有丝分裂不断增殖,体细胞的分裂速度相对更快,生殖细胞被多层体细胞完全包绕,原始性腺的横切面呈瘤状,并分布有血细胞(图2a)。21 dph(平均全长为 37.5 mm)和 24 dph(平均全长为 53.0 mm),原始性腺逐渐延长,体积不断增大,两侧性腺的一端与腹腔系膜相连,而另一端则游离于腹腔背侧,每侧性腺的横切面呈细长的棒状,面积约为 80 μm×15 μm(图2b,图2c)。27 dph(平均全长为 68.2 mm)和 30 dph(平均全长为79.3 mm),原始性腺的横切面进一步增大,中部逐渐变宽而两端较窄,生殖细胞分裂加快,数量显著增加,但并未出现明显的性腺分化特征(图2d,图2e)。
34 dph(平均全长为 90.6 mm),原始性腺开始出现分化,可观察到部分性腺组织中排列着被体细胞包绕呈簇状发育的生殖细胞群,部分生殖细胞的直径为5 μm左右,呈卵圆形,胞质较少,细胞核占比大,已显现出卵原细胞的形态学特征,预示将向卵巢方向分化(图2f);仍有部分个体的原始性腺尚未出现分化特征,其横切面相对较为狭长,生殖细胞数量较少,且在性腺靠近系膜的一端形成微血管,通过对比,推测为未发生分化的精巢(图2g)。
40 dph(平均全长为 107.9 mm),在推定的精巢组织中,生殖细胞经有丝分裂大量增殖并聚集形成精原细胞群(图2h)。44 dph(平均全长为 118.4 mm),卵巢中的生殖细胞已基本分化为卵原细胞,血管发达,血细胞数量较多且散布于整个性腺组织内,部分体细胞聚集于性腺组织的中央区域,形成一条着色较深的细胞带;随后这些聚集的性腺体细胞向两侧扩张远离,在原来细胞带的位置产生一条狭窄的裂隙,即为卵巢腔的雏形,标志着卵巢解剖学上分化的开始(图2j)。48 dph(平均全长为 131.4 mm)和 52 dph(平均全长为147.7 mm),精巢组织内的体细胞和精原细胞进一步分裂,横切面明显增大并散布有少量血细胞,精原细胞集中分布于精巢的腹侧边缘,被一层基底膜所包被形成囊状的精原细胞团,细胞团之间由小裂缝隔开,为精母细胞囊的形成做准备,表明精巢已开始细胞学上的分化(图2i);卵巢组织中的裂隙不断向两侧延伸和扩大,最终形成明显的卵巢腔,卵原细胞通过分裂和聚集形成生殖细胞囊,细胞直径也增大至15~20 μm,逐渐向初级卵母细胞过渡,嗜碱性较弱,被染成浅紫色(图2k,图2l)。
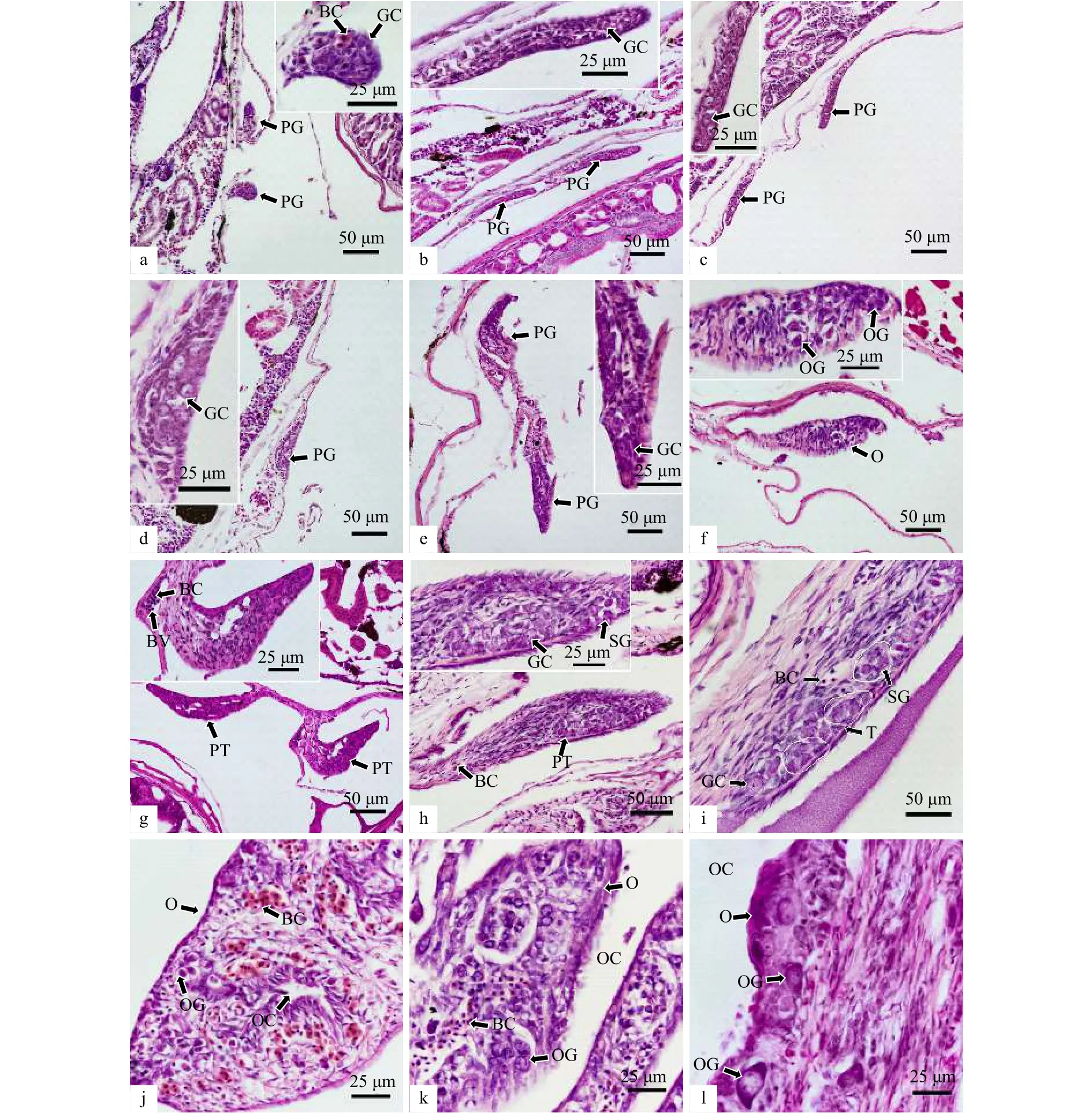
图2 军曹鱼原始性腺的分化Fig.2 Differentiation of primary gonad of Rachycentron canadum
3.3 精巢的首周年发育特征
根据精巢中生殖细胞的种类、成熟度、数量占比及排列方式,可将60~360 dph军曹鱼的精巢样品分为5个发育分期,分别为精原细胞增殖期(I期)、精母细胞生长期(II期)、精母细胞成熟期(III期)、精子细胞变态期(IV期)、精子成熟期(V期),尚未发现精子退化吸收期(VI期),各发育期精巢特征如下。
60 dph(体长为(22.6±1.4)cm),精巢发育至 I期,呈细线状,横切面积较小,精原细胞的数量较多,聚集于囊泡状的结构内,分布于精巢的中部及腹侧,同时也可观察到少量由精原细胞转化而成的初级精母细胞和次级精母细胞(图3A,图3a)。
90 dph(体长为(30.1±1.6)cm),精巢发育已进入II期,生殖细胞的数量较少,结缔组织及体细胞占整个精巢组织的绝大部分。精巢中已形成零星的精小叶,精小叶内可观察到精原细胞、初级精母细胞以及次级精母细胞(图3B,图3b)。
120 dph(体长为(36.8±1.4)cm),精巢发育由 II期向III期过渡,精小叶数量增多,集中分布于精巢的外缘。精小叶中形成精小囊,精小囊内以初级精母细胞和次级精母细胞为主,同时也出现了少量的精细胞(图3C,图3c)。
150 dph(体长为(40.3±2.0)cm),精巢发育进入III期,精小叶数量进一步增多,随着精母细胞的成熟分裂,精小囊逐渐向精小叶腔扩展。当精细胞成熟后,精小囊破裂,将其释放至精小叶腔中,因此在精小叶内可见大量的初级精母细胞、次级精母细胞、精细胞以及少量的精原细胞(图3D,图3d)。
185 dph(体 长 为 (46.5±0.4)cm),精 巢 仍 处 于III期,精小叶散布于整个精巢组织中,精小囊内初级精母细胞、次级精母细胞成堆出现,精小叶腔中精细胞的数量明显增多(图3E,图3e)。
210 dph(体长为(49.5±1.2)cm),精巢已发育至IV期,精小叶的体积增大,形状逐渐变得狭长,位于精巢边缘的精小囊体积较大,以次级精母细胞为主,而精巢内部的精小囊则基本破裂,精小叶腔中充斥精细胞,小部分精细胞已开始变态为精子(图3F,图3f)。
360 dph(体长为(63.4±1.3)cm),精巢处于 V 期,精小叶进一步扩大变得更为狭长,从精巢中央向边缘呈辐射状分布,精小囊分布于精小叶间的交界处,仍有少量的精母细胞,大部分精细胞已变态为精子,可观察到密集的精细胞与成熟精子充满整个精小叶腔(图3G,图3g)。

图3 军曹鱼精巢首周年发育的组织学变化Fig.3 The histologic changes of first annual testicular development of Rachycentron canadum
3.4 卵巢的首周年发育特征
根据卵母细胞的发育特征及成熟度,可将60~360 dph军曹鱼的卵巢样品划分为卵原细胞增殖期(I期)、初级卵母细胞小生长期(II期)和初级卵母细胞大生长期(III期),卵巢尚未发育至成熟期,各发育期卵巢特征如下。
60 dph(体长为(23.2±1.7)cm),卵巢发育至 I期早期,卵巢腔占据整个卵巢的绝大部分,卵巢的横切面呈环形,内缘呈褶皱状,部分凸起向卵巢腔中央延伸形成产卵板,产卵板内以卵原细胞为主,细胞质含量较少,具有明显的细胞核,个别卵原细胞已向初级卵母细胞过渡,分裂为第I时相卵母细胞(图4A,图4a)。
90 dph(体长为(31.5±1.7)cm),卵巢仍处于 I期,具有明显的卵巢腔,产卵板较短,以卵原细胞及其分裂分化而成的第I期时相卵母细胞为主,细胞核占比较大,核内染色质呈丝状并组成稀疏的网状结构(图4B,图4b)。
120 dph(体长为(37.1±2.0)cm),卵巢发育由 I期向II期过渡,产卵板逐渐延长,大部分为第Ⅱ时相卵母细胞,细胞直径明显增大,呈椭圆形或不规则形,细胞质呈强嗜碱性,被染成紫蓝色,卵母细胞间的间隔较宽,在产卵板的边缘仍可观察到少量卵原细胞和第I时相卵母细胞(图4C,图4c)。
150 dph(体长为(42.9±1.5)cm),卵巢发育进入II期,卵母细胞基本进入第II时相,在大部分卵母细胞的细胞质中可观察到一个被染成深紫色的团块状卵黄核,这是早期阶段第II时相卵母细胞的特征(图4D,图4d)。
185 dph(体长为(47.7±1.6)cm),卵巢处于 II期,产卵板内的卵母细胞排列更为紧密,细胞核与细胞质相应增大,部分直径较大的卵母细胞中核仁的数量增加至2~3个,细胞质内出现数颗大小不一的油滴(图4E,图4e)。
210 dph(体 长为 (52.6±0.9)cm),卵 巢 仍 处 于II期,卵母细胞的核仁直径明显增大,并继续分裂出多个较小的核仁,嗜碱性减弱,被染成浅紫色,而细胞质中的油滴数量进一步增多(图4F,图4f)。
360 dph(体长为(67.17±1.3)cm),卵巢发育进入III期,以早期第III时相卵母细胞为主,细胞直径增大至100~150 μm,核仁分布于核膜内缘,油滴数量较多呈环带状分布,并向细胞质外缘逐渐扩展,质膜外已开始形成两层滤泡膜,仍有少数卵母细胞处于第II时相(图4G,图4g)。
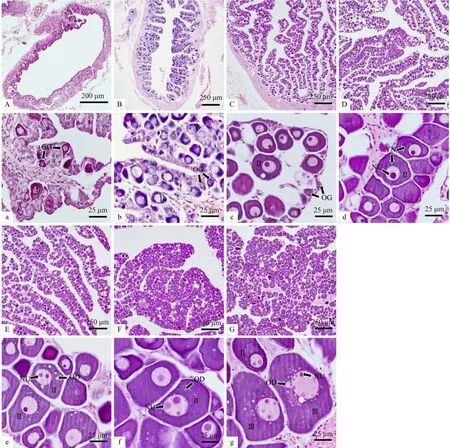
图4 军曹鱼卵巢首周年发育的组织学变化Fig.4 The histologic changes of first annual ovarian development of Rachycentron canadum
4 讨论
4.1 军曹鱼原始生殖细胞的迁移及性腺的分化特征
本研究通过连续的组织学观察,详细记录了军曹鱼原始生殖细胞的形态特征变化、迁移方式,原始性腺的形成及分化规律。军曹鱼原始生殖细胞在迁移过程中保持单个游离的状态且未发生有丝分裂,与大多数已报道硬骨鱼类中的研究结果相同,如革胡子鲶(Clarias gariepinus)[12]、 大 菱 鲆 (Scophthalmus maximus)[13]、牙鲆(Paralichthys olivaceus)[14]、泥鳅(Misgurnus anguillicaudatus)[15]等。军曹鱼原始生殖细胞迁移至生殖嵴后被体细胞完全包绕,于15 dph时形成芽状的原始性腺,相似地,大黄鱼(Larimichthys crocea)原始性腺的形成时间为 16 dph[16];大菱鲆[13]和牙鲆[14]分别在 20 dph 和 22 dph 时形成原始性腺,而稀有鲫(Gobiocypris rarus)[17]的原始性腺则形成于 9 dph,表明硬骨鱼类原始性腺在形成时间上存在明显的种间差异。原始性腺发育早期体细胞有丝分裂速度加快,而生殖细胞的有丝分裂进程在后期才明显加快。Yang等[14]在牙鲆中也发现了相似的现象,据此推测,在胚后迁移及原始性腺早期发育阶段,生殖细胞可能处于分裂受阻的状态。在34 dph军曹鱼稚鱼中可发现两类存在明显特征差异的性腺,第1类性腺横切面呈水滴形,生殖细胞数量较多,推测其将发育为卵巢,第2类性腺的横切面较狭长,生殖细胞数量也较少,则推定为分化前的精巢;同样地,在大菱鲆[13]、青鳉(Oryzias latipes)[18]、三刺鱼(Gasterosteus aculeatus)[19]和蓝鳃太阳鱼(Lepomis macrochirus)[20]性腺分化的研究中,推定的卵巢组织中生殖细胞数量均显著高于精巢。随后,军曹鱼的第1类性腺在44 dph时形成卵巢腔,标志着卵巢分化开始。由此可见,原始性腺中生殖细胞数量的差异可作为判定军曹鱼性腺分化方向的标准之一。50 dph时,推定的精巢中精原细胞聚集于性腺腹侧,并由基底膜包被形成精原细胞囊,预示着精巢分化的开始[21],但此时尚未见输精管和精小叶等精巢分化的主要标志出现。上述结果表明,军曹鱼的性腺分化属于分化型雌雄异体型,且卵巢的分化时间要早于精巢。
4.2 军曹鱼性腺发育的首周年变化
360 dph军曹鱼精巢可发育至V期,而卵巢只发育至III期早期,由此推测雄鱼的性成熟年龄约为1龄,而此时雌鱼尚未达到性成熟。据报道,自然海区及网箱养殖军曹鱼的最小性成熟生物学年龄分别为:雄性1龄,雌性1.5~2龄[22-24],由此可见,室内水池养殖条件下,军曹鱼性腺发育规律与野生及网箱养殖环境下无明显差异。
通过对军曹鱼精巢的组织学观察发现,精小叶的内缘排列着不规则的精小囊,同一精小囊内的生精细胞发育程度一致,当精小囊内的生精细胞成熟后,囊膜便破裂,将精子释放至精小叶腔中并最终汇集于输精管,这与大部分硬骨鱼类如四指马鲅(Eleutheronema tetradactylum)[25]、黄斑篮子鱼(Siganus oramin)[26]、金钱鱼(Scatophagus argus)[27]、松江鲈(Trachidermus fasciatus)[28]等的生精特点相一致。军曹鱼的精子形成过程与其他硬骨鱼类相同,生殖细胞由最初的精原细胞经转化后形成初级精母细胞,随后通过减数第1次分裂形成次级精母细胞,再经进一步的分裂产生精细胞,最后精细胞经变态形成成熟精子[29]。精巢成熟后精小叶的形状狭长且从精巢的中央向边缘呈辐射状分布,根据此结构特征及生精细胞的发育特点可将其归类为小叶型结构中的辐射型精巢,如鲇形目与鲈形目中大多数种类的精巢均属此类型[30]。
依据不同日龄军曹鱼卵巢的组织学特征可知,I期卵巢主要包含卵原细胞及向初级卵母细胞过渡的第I时相卵细胞,细胞体积相对较小,核内染色质呈明显的细丝状。目前,关于第I时相卵细胞的定义尚未趋于统一,部分学者将其归类至卵原细胞[31-32],还有部分学者将其归类为卵母细胞[27,33],基于细胞特征及有丝分裂的程度,本文将其定义为第I时相卵母细胞。军曹鱼II期卵巢中含有第I时相、第II时相卵母细胞,而III期卵巢中含有第I时相、第II时相、第III时相卵母细胞,卵巢内卵母细胞的发育并不同步,这与黄斑篮子鱼[26]、红鳍笛鲷(Lutjanus erythopterus)[31]、半滑舌鳎(Oplegnathus punctatus)[32]等鱼类似,由此可判定军曹鱼属于分批同步产卵型鱼类,一年可多次产卵。当卵母细胞发育至第II时相中期时,细胞质内开始形成油滴,随着进一步的发育,油滴的数量不断增多并呈环带状分布于细胞核周缘,与银鲳(Pampus argenteus)[34]、灰鲳(Pampus cinereus)[35]和许氏平鲉(Sebastes schlegeli)[36]的研究结果相一致,而金钱鱼(Scatophagus argus)[27]、黑鱾(Girella leonina)[37]和美洲鲥鱼(Alosa sapidissima)[38]中则发现卵母细胞发育至第III时相才开始形成油滴,由此推测,军曹鱼卵母细胞对脂质的积累能力相对较强。
