基于DNA条形码的长江口鱼类浮游生物形态分类研究
2021-09-01刘守海杨旖祎何彦龙季晓王宇坛张海景毛如意蒋晓山程祥圣
刘守海,杨旖祎,何彦龙,季晓,王宇坛,张海景,毛如意,蒋晓山,程祥圣*
( 1.国家海洋局东海环境监测中心,上海 201206;2.自然资源部海洋生态监测与修复技术重点实验室,上海 201206;3.国家海洋局东海海洋环境调查勘察中心,上海 200137)
1 引言
鱼类浮游生物(Ichthyoplankton)是鱼卵、仔鱼和稚鱼的统一简称,鱼卵和仔、稚鱼是鱼类生活史早期发育的重要阶段。作为鱼类种群补充的重要来源,它们的种类分布和数量变化是评估海域鱼类产卵场、亲鱼资源量和渔业资源补充量最直接有效的数据资料之一。仔、稚鱼既是生物能的消费者,又是生物能的转换者,是海洋食物链中的重要环节之一,基于此,鱼类浮游生物已成为海洋生物多样性与海洋生态学的重要研究对象[1]。准确鉴定鱼类浮游生物是开展这些研究的基础。鱼类发育过程的形态变化非常复杂,因此,如何系统准确地找出各发育阶段的形态变化、找出最基本的特征是这一基础的前提。通过传统形态学方法来认识鱼类早期发育变化的方法有3种[2]:第1种是人工授精法,通过对某种成鱼进行人工授精获得的样品与天然采集到的样品进行比较来鉴别种类[3-4],这种方法鉴定的种类是绝对可靠的,但往往受海上条件所限,在海上进行现场人工授精是比较困难的;第2种方法是动态研究法,即用大量不同大小的标本,按发育期先、后进行追踪观察,比较形态和器官的发育差异,以形态进行分类[5-8];第3种方法是静态研究法,是以单个个体完整性为重点,追寻个体形态发育阶段的主要特征,其优点是仅用少数标本便可进行鉴定分类,但此方法要求鉴定者非常熟悉各科、属、种的幼体形态特征[9]。
随着科学技术的进步,鱼类浮游生物鉴定工作的准确性得到很大的提升,如扫描电镜技术的发展让研究者可以通过电子显微镜观察鱼卵的卵膜结构来进行种类的鉴定[10-12]。近年来,随着DNA条形码(DNA barcoding)技术在鱼类鉴定研究中广泛应用[13],更多学者尝试利用DNA条形码对鱼类浮游生物进行种类鉴定。Pegg等[14]利用COⅠ基因对澳大利亚南部大堡礁的仔鱼进行了鉴定。Valdez-Moreno等[15]利用DNA条形码对墨西哥尤卡坦半岛附近海域的成鱼和仔鱼(早期鱼类)进行种类鉴定研究。Ko等[16]应用DNA条形码评估了5家研究机构仔鱼形态鉴定结果的精确度,通过DNA条形码鉴定到种水平的仔鱼占69%。该研究认为,DNA条形码技术是种类鉴定的最好方法之一。周美玉等[17]将从形态学特征鉴定为2种类型鱼卵和3种类型仔、稚鱼的样品,应用DNA条形码技术分析鉴定出鱼卵种类为3种、仔、稚鱼种类为4种。刘守海等[18]通过DNA条形码技术鉴定调查所获得的7目10科样品,鉴定到种的比例高达94.7%。周晓梦等[19]应用DNA条形码和形态对短吻红舌鳎(Cynoglossus joyneri)鱼卵、仔鱼进行了鉴定。以上结果说明,DNA条形码应用于鱼卵和仔、稚鱼的鉴定可大大提高种类鉴定结果的精确性,这些研究大大提高了鱼类浮游生物的鉴别准确性。
长江口水域由于长江径流带来了大量的营养物质,孕育了丰富的饵料资源,使之成为多种经济鱼类的产卵、育幼和索饵的重要场所[20]。长江口的鱼类浮游生物研究始于20世纪80年代[21-24],长江口共有鱼类浮游生物56科123种,其中鉴定到种为93种,鉴定到属为6种,鉴定到科为17种,未定种为7种。以上调查皆通过形态获得种类鉴定结果,且有一定比例的种类未定到种。本文以2016年5月和8月及2017年8月长江口及其邻近海域的仔、稚鱼样品为材料,采用形态学与DNA条形码相结合的形式进行鱼类浮游生物分类研究,获得了长江口春夏季鱼类浮游生物种名录,通过摄影、绘图等手段,记录并描述报道较少种类的鱼类早期发育阶段的形态特征,为河口鱼类育幼场的研究提供了科学依据,也为鱼类早期发育阶段资料的积累探索了新途径。
2 材料与方法
2.1 样品采集和形态学鉴定
于2016年5月、8月以及2017年8月在长江口及其邻近海域设置采样站点(图1),按照《海洋调查规范》(GB/T 12763.6-2007)使用浅水Ⅰ型网(网长为145 cm、网口内径为50 cm、网口面积为0.5 m2、筛绢孔径为0.505 mm)和大型浮游生物网(网长为280 cm、网口内径为80 cm、网口面积为0.5 m2、筛绢孔径为0.505 mm)在站点分别进行垂直拖网和水平拖网。采集到的标本现场进行初步处理,用无水乙醇保存。
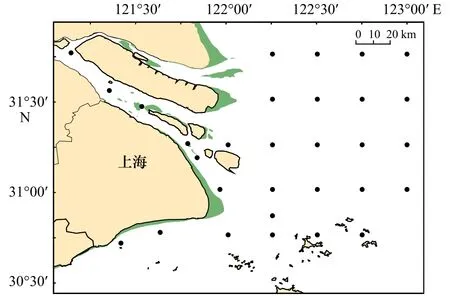
图1 长江口海域采样站位图Fig.1 Sampling stations in the Changjiang River Estuary
在室内Nikon SMZ 25体视显微镜(尼康,日本)下,根据鱼卵和仔、稚鱼形态特征,对样品进行初步鉴定[2,25]。用NIS-Elements D软件进行拍照并获取体长、头长、眼径等参数。从形态鉴定结果中,按1~10、10~50、50~100、大于 100的数量组,分别取1个、3个、5个、10个个体进行DNA条形码分子实验。
2.2 DNA条形码分析方法
鱼卵取单个整粒鱼卵,体长小的仔鱼取整条,较大的仔、稚鱼取尾部。取肌肉约100 mg,使用海洋动物组织基因组DNA 提取试剂盒(北京天根生化科技有限公司)提取总DNA。使用通用引物F1:TCR ACY AAY CAY AAA GAY ATY GGC AC和 R1:TAG ACT TCW GGG TGR CCR AAG AAT CA扩增仔、稚鱼的线粒体DNA 的COⅠ部分序列,PCR总反应体系为50 μL,其中包括:10×PCR 缓冲液 5 μL、dNTP 4 μL(浓度为2.5 mmol/L)、上下游引物各 2 μL(浓度为 10 mmol/L)、Taq DNA 聚合酶 0.8 μL(浓度为 5 U/μL)、模板 DNA 1 μL,加双蒸水至总体积 50 μL。样品在 AG-22331 型PCR仪上进行扩增,94℃预变性5 min;94℃ 变性30 s、52℃ 退火 45 s、72℃ 延伸 1 min,35 个循环;72℃ 延伸10 min;4℃保存。扩增产物使用1.0%的琼脂糖凝胶电泳检测后送至上海杰李生物技术有限公司胶回收双向测序后拼接。
2.3 数据处理
利用DNAStar 软件包对测得的序列和GenBank下载序列进行校对,用Clustal X对所有序列进行比对排序并去除两端冗余列。将确定有效的COⅠ基因序列在 Bold System(Barcode of Life Data System)数据库和 GenBank中进行BLAST分析来物种鉴定。采用遗传相似度不低于99%的为同一物种,92%~99%为同一属,85%~92%为同一科的标准对本调查海域的鱼类浮游生物进行种类鉴定[13,16],对于比对到不同物种上的序列,进一步通过构建系统发育关系树来确定物种。结果按照《拉汉世界鱼类名典》[26]进行同种异名的校正。
3 结果与分析
3.1 2016−2017 年长江口及邻近海域鱼类浮游生物种类组成
2016年5 月、8月以及2017年8月,调查共获得鱼类浮游生物9目19科55种,其中鉴定到种的有45种,鉴定到属的有5种,鉴定到科的有5种,鉴定到种的比例为81.8%。鲈形目种类数最多有33种,其次是鲤形目有6种,鲱形目和鲽形目含有相同种类数都为4种,鲻形目2种,鲀形目2种,鲉形目2种,鳗鲡目1种,仙女鱼目1种。在鲈形目中,以虾虎鱼科和石首鱼科最多,两者占鲈形目种类的近80%(表1)。
从种类组成的季节分布来看,鱼类浮游生物类群的季节变化不明显,其中,以鲤形目的鲤科,鲱形目的鳀科,鲈形目的石首鱼科和虾虎鱼科为主。但在种类上有明显的季节变化,夏季鱼类浮游生物种类数多于春季。在5月和8月均出现的种类仅有凤鲚(Coilia mystus)、日本鳀(Engraulis japonius)和小黄鱼(Larimichthys polyactis)。仅在5月出现的种类有16种,以淡水的鲤科和咸淡水的虾虎鱼科为主,包括鲤科的鲢鱼(Hypophthalmichthys molitrix)、青鱼(Mylopharyngodon piceus)、鳊鱼(Parabramis peknensis),虾虎鱼科的斑点竿虾虎鱼(Luciogobius guttatus)、睛尾蝌蚪虾虎鱼(Lophiogobius ocellicauda)、斑尾刺虾虎鱼(Acanthogobius ommaturus)等。仅在8月出现的种类有36种,以虾虎鱼科(14个)、石首鱼科(6个)和鲤科(3个)为主。包括鲤科的寡鳞飘鱼(Pseudolaubuca engraulis)、银飘鱼(Pseudolaubuca sinensis),石首鱼科的棘头梅童鱼(Collichthys lucidus)、皮氏叫姑鱼(Johnius belangerii)、叫姑鱼(Johnius grypotus)、鮸(Miichthys miiuy),虾虎鱼科的阿部氏鲻虾虎鱼(Mugilogobius abei)、髭缟虾虎鱼(Tridentiger barbatus)、孔虾虎鱼(Trypauchen vagina)、拉氏狼牙虾虎鱼(Odontamblyopus lacepedii)等。
3.2 形态描述
我国已知海洋鱼类已超3 000种,但资料可查的鱼卵与仔、稚鱼种类不超过300种[2,27]。在传统方法研究下,一些仔、稚鱼常见种或隐存种[18],如龙头鱼(Harpadon nehereus)、前鳞龟鰉(Chelon affinis),没有得到认识。仔、稚鱼发育是连续变化的一个过程,目前有很多种类只有某个阶段的形态特征描述及图谱[28],如四指马鲅(Eleutheronema tetradactylum)、少鳞鱚等。因此,本文选择之前报道较少种类进行形态特征描述,并绘制手绘图,获得分类鉴定资料。
3.2.1 龙头鱼Harpadon nehereus(Hamilton 1822 )
(1)分类地位
仙女鱼目Aulopiformes,狗母鱼科Synodontidae,龙头鱼属Harpadon。
(2)参考序列
JX534239,KP112261。
(3)形态特征
全长为5.97 mm,体长为5.82 mm。卵黄囊已全部吸收。头长为0.83 mm,占体长的14.3%,眼径为0.29 mm,占头长的34.9%。眼变黑,口、咽、消化道已经相通,消化道细长,自胸鳍基至肛门处的消化道背缘两侧有3对椭圆形黑色素斑块,3对色素斑呈等间隔分布。肛门位于体中央稍后。肛前距为4.11 mm,占体长的70.6%。肛后尾部腹缘有1个菊花状黑色素。尾端脊索平直。背部鳍膜起始于颅顶,然后向后一直延伸到尾部鳍膜,无色透明,腹部鳍膜起点略低于背部鳍膜(附图 A1a)。
全长为 9.75 mm,体长为 9.24 mm。头长为 1.25 mm,占体长的13.5%,眼径为0.47 mm,占头长的37.6%。口裂增大。胸鳍基至肛门前共有5对椭圆形黑色素斑块,在原来第1对、第2对之间和原来第3对后面,各增加1对色素斑。肛门位于体中央稍后。肛前距为6.84 mm,占体长的74.0%。肛后尾部腹缘有1个菊花状黑色素。脊索向上翘。臀鳍鳍条出现,尾鳍有20~22 鳍条(附图 A1b)。
全长为30.55 mm,体长为26.35 mm。头长为3.94 mm,占体长的15.0%,眼径为1.31 mm,占头长的33.2%。肛门前共有6对椭圆形黑色素斑块,胸鳍前增加1对黑色素斑。肛门位于体中央稍后。肛前距为19.34 mm,占体长的73.4%。肛后尾部腹缘菊花状黑色素消失。第一背鳍10鳍条,脂鳍出现,腹鳍鳍条形成。尾鳍基底出现4个较小斑块状色素(附图A1c)。
全长为40.37 mm,体长为34.27 mm。头长为6.73 mm,占体长的19.6%,眼径为1.25 mm,占头长的18.6%。两侧至肛门前共有7对黑色素斑。肛门位于体中央稍后。肛前距为25.81 mm,占体长的75.3%。第一背鳍12鳍条,脂鳍始点在臀鳍中间位置。腹鳍15鳍条。臀鳍8鳍条。尾鳍基底4个黑色素块消失2个,仅剩2处较小斑块状色素(附图A1d)。
(4)研究概况
有关龙头鱼早期生活史的研究,Delsman[29]记录了0.8 mm的龙头鱼鱼卵;冲山宗雄[9]对日本海域体长为25~30 mm的龙头鱼稚鱼的形态作过描述。本文描述龙头鱼从仔鱼到稚鱼(体长为5.82 ~34.27 mm)的个体发育形态,补充了仔、稚鱼形态特征变化的描述。
3.2.2 前鳞龟鰉Chelon affinis(Günther 1861)
(1)分类地位
鲻形目Mugiliformes,鲻科Mugilidae,龟鰉属Chelon。
(2)参考序列
JF911709。
(3)形态特征
全长为26.54 mm,体长为21.82 mm。头长为6.51 mm,占体长的29.8%,眼径为2.13 mm,占头长的32.7%。上下颌有色素,自头后至尾柄的体侧中轴以上星状、树枝状和菊花状色素不规则分布。臀鳍基部分布树枝状和菊花状色素。肛门位于体中央稍后。肛前距为15.00 mm,占体长的68.7%。所有鳍条都已形成。背鳍2个,第一背鳍II鳍棘5鳍条,第二背鳍II鳍棘7鳍条。臀鳍起始位置比第二背鳍起点略靠前,臀鳍II鳍棘 12 鳍条(附图 A2)。
(4)研究概况
冲山宗雄[9]对日本海域体长为4.1~12.0 mm的前鳞龟鰉形态作过描述。国内未见前鳞龟鰉早期生活史的研究报道。本文描述了体长为21.82 mm前鳞龟鰉稚鱼的形态。
3.2.3 四 指 马 鲅Eleutheronema tetradactylum(Shaw 1804)
(1)分类地位
鲈形目Perciformes,马鲅科Polynemidae,四指马鲅属Eleutheronema。
(2)参考序列
KC878730。
(3)形态特征
全长为11.18 mm,体长为9.88 mm。体前部呈椭圆形,头较大,头长为3.02 mm,占体长的30.6%,眼径为0.69 mm,占头长的22.8%。吻钝圆,上颌骨有左右对称的1对黑色素。肛门位置约为体中部,肛前距为5.68 mm,占体长的57.5%。脊索尾椎骨向上弯曲。背鳍2个,第一背鳍VI鳍棘,第二背鳍I鳍棘15 鳍条。臀鳍位置与第二背鳍相对应,臀鳍II鳍棘17鳍条。腹部胸鳍下方有1个黑色素。臀鳍基有1排黑色素,始于臀鳍中部,约10个附图(A3)。
(4)研究概况
有关四指马鲅早期生活史的研究,多以人工培育下四指马鲅早期发育的形态及生长的观察为主[30-32]。王晓东[33]描述了全长为13.4 mm四指马鲅稚鱼的形态,各鳍鳍条数目基本稳定,鳞片开始出现。本文较详细描述了体长为9.88 mm四指马鲅稚鱼的形态,发现臀鳍基有1排黑色素,始于臀鳍中部,约10个。
3.2.4 少鳞鱚Sillago japonica(Temminck et Schlegel 1843)
(1)分类地位
鲈形目Perciformes,鱚科Sillaginidae,鱚属Sillago。
(2)参考序列
AP017438。
(3)形态特征
全长为13.89 mm,体长为12.71 mm。头长为3.35 mm,占体长的26.4%,眼径为1.15 mm,占头长的34.3%。头部眼后方有数个小块状黑色素,下颚、腹部正中线、臀鳍基底有点状黑色素分布,尾柄背缘、腹缘及体侧正中线有斑块状黑色素分布,尾鳍基底出现2丛黑色素。肛门位于体中央略前。肛前距为6.07 mm,占体长的47.8%。第一背鳍VIII鳍棘,第二背鳍19鳍条,胸鳍、腹鳍、臀鳍和尾鳍鳍条已形成(附图A4)。
(4)研究概况
有关少鳞鱚早期生活史的研究,水戸敏[34-35]对日本福冈外海的少鳞鱚进行了人工授精、孵化实验,详细描述了鱼卵和仔、稚鱼的发育与形态;冲山宗雄[9]对体长为2.6~16.5 mm的少鳞鱚仔、稚鱼的形态作过描述;王晓东[33]描述了全长为6.5~16.5 mm少鳞鱚仔稚鱼的形态。本文描述了体长为12.71 mm少鳞鱚稚鱼的形态,发现尾柄背缘、腹缘及体侧正中线有斑块状黑色素分布,尾鳍基底出现2丛黑色素。
3.2.5 日 本 须 鳎Paraplagusia japonica(Temminck &Schlegel 1846)
(1)分类地位
隶属于鲽形目Pleuronectiformes,舌鳎科Cynoglossidae,须鳎属Paraplagusia。
(2)参考序列
JQ639066。
(3)形态特征
全长为6.08 mm,体长为5.92 mm。头长为1.42 mm,占体长的24.0%,眼径为0.36 mm,占头长的25.4%。卵黄囊仍未完全消失,卵黄囊下方有一簇色素。口钝圆,上、下颌及鳃盖骨形成。颅顶上方棘冠出现2条长棘条,长度分别为1.13 mm和1.49 mm。体部背缘有5个丛状黑色素,每丛5~6个树枝状黑色素或点状黑色素,腹缘有1列不规则排列的黑色素分布。肛门位于体中央略前。肛前距为2.51 mm,占体长的42.4%。尾部鳍膜上出现弹性丝(附图A5)。
(4)研究概况
有关日本须鳎早期生活史的研究,男卓志[36]报道了日本若狭湾西部及其邻近海域直径为0.8 mm左右的日本须鳎卵和体长为5.95~12.30 mm的日本须鳎仔、稚鱼形态;丘台生[37]描述了我国台湾沿海体长为4.0~7.9 mm的日本须鳎仔稚鱼形态;冲山宗雄[9]对体长为10.2~12.3 mm的日本须鳎仔稚鱼的形态作过描述;万瑞景和张仁斋[25]描述了1984年6月采自东海海域的体长6.80 mm的日本须鳎仔鱼形态,体侧背缘自头后至尾端有10余个丛状黑色素,体侧的腹缘也有9个丛状黑色素分布。本文描述了体长为5.92 mm日本须鳎仔鱼的形态,发现体部背缘有5个丛状黑色素,每丛5~6个树枝状黑色素或点状黑色素,腹缘有1列不规则排列的黑色素分布。
4 讨论
DNA条形码技术是一项通过标准化的短基因片段的序列差异来对物种进行鉴定的技术。相对于传统的形态分类方法,DNA条形码技术最大的优势在于它能准确鉴定不同发育阶段的个体、形态相似度较高的物种和因缺少形态学分类数据而导致很难进行区分的个体[38]。这项技术运用到鱼类浮游生物的鉴定中,快速解决了很多传统形态学分类方法无法解决的问题[28]。物种判别标准是分子鉴定研究中最重要的因素,根据Ward等[39]、Ko等[16]、周美玉等[17]的研究结果,本文采用遗传相似度不低于99%的为同一物种、92%~99%为同一属、85%~92%为同一科的标准对本调查海域的鱼类浮游生物进行种类判定。大多数的种类能够准确鉴定到种(表1)。部分样品序列因与某一物种相似度不高而未鉴定到种,如有序列与数据库中的寡鳞飘鱼相似度最大为98%,故鉴定到飘鱼属。有些样品序列与两个种类的相似度均为99%~100%,且这些种类在本调查区域均有出现[40-41],故只将其鉴定到科或属的水平。如最后鉴定为鲻虾虎鱼属的序列,其与数据库中的阿部氏鲻虾虎鱼和粘皮鲻虾虎鱼(Mugilogobius myxodermus)相似度为99%以上,且二者在本调查海域均有分布记录。本研究中样品序列与大鳍弹涂鱼(Periophthalmus magnuspinnatus)和青弹涂鱼(Scartelaos histophorus)相似度均为99%的序列,因此无法准确鉴定,故将其鉴定为虾虎鱼科[42]。此外,本研究鉴定的序列,与数据库中黑鳃梅童鱼(Collichthys niveatus)(HM219223)和小黄鱼相似度达99%以上,根据Cheng等[43]研究可知,黑鳃梅童鱼序列疑为形态鉴定有误,故本研究将其鉴定为小黄鱼。因此数据库的完整性与可靠性对鉴定结果的准确性至关重要。
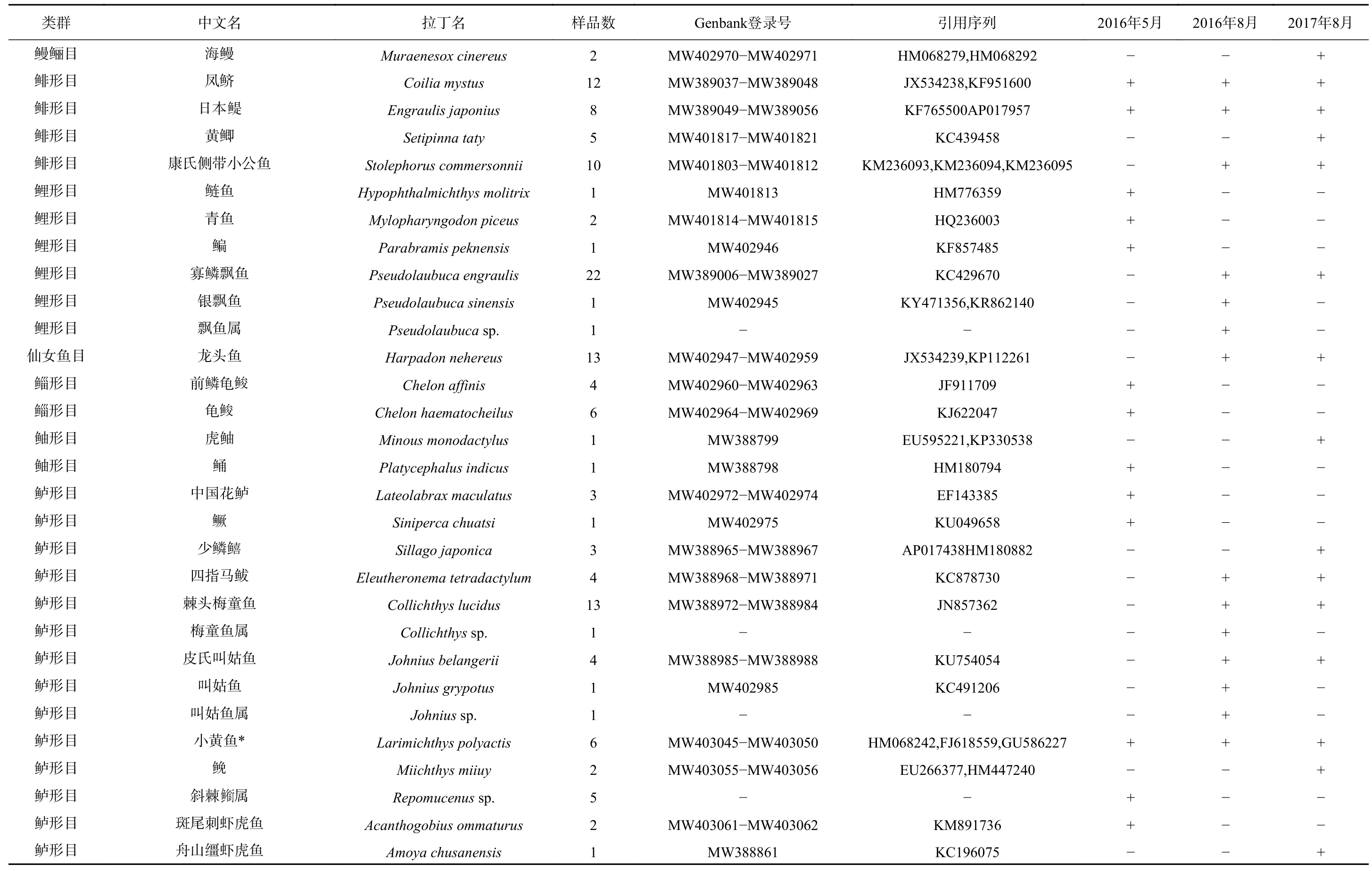
表1 长江口鱼类浮游生物种名录(2016年春季(5月)、夏季(8月)和2017年夏季(8月))Table 1 List of ichthyoplankton species of Changjiang River Estuary(spring (May),summer (August) 2016 and summer (August) 2017)
长江口鱼类浮游生物在种类上有明显的季节变化,而类群无明显的季节变化。本研究中,仅在5月出现的种类有16种,有36种仅在8月出现。在温带和亚热带河口,鱼类浮游生物的数量在夏季达到峰值,且具有优势种种类季节变动模式[23]。本研究中,河口鱼类浮游生物主要类群为虾虎鱼科和鲤科,但同一类群里种类有季节变化,如同为鲤科,5月出现的种类为鲢鱼、青鱼、鳊鱼,8月出现的种类为寡鳞飘鱼、银飘鱼。根据生态位竞争排斥原理,生态上类似的种很少能共存于一个生态位中,因而生活在一起的各个物种都必须有它们自己独特的生态位[44],即相同科或者相同属鱼类在时间上出现明显的错位分布,因为生态位接近,时间错位分布可以减少对空间和食物资源的竞争[45]。这一现象在长江口成鱼也有出现,鲳科鲳属的银鲳(Pampus argenteus)(出现于 5-8月)和灰鲳(Pampus cinereus)(出现于 8-9 月),尾数有明显的时间错位分布现象[45]。成鱼的繁殖策略和产卵区域水体环境的季节变化是河口鱼类浮游生物种类季节变动的主要因素之一[46-47]。
本研究采用传统形态学和现代分子生物学方法相结合的研究思路,在DNA条形码技术的辅助鉴定下,收集了鱼类浮游生物形态特征,积累了鱼类早期发育的形态分类信息[28]。本研究首次描述了隐存种[18]前鳞龟鰉稚鱼、河口常见优势种[48]龙头鱼仔稚鱼阶段的形态特征;报道了少鳞鱚、四指马鲅和日本须鳎等种类在不同发育阶段或不同体长的特征。这些工作完善了鱼类浮游生物的分类鉴定资料,为鱼类早期发育阶段分类资料的积累探索了新途径。
致谢:何日光、彭小家、陈耀辉等同志在室内样品和数据处理等方面做了大量工作,谨致谢忱。特别感谢陈莲芳研究员对本文审阅并提出宝贵意见。
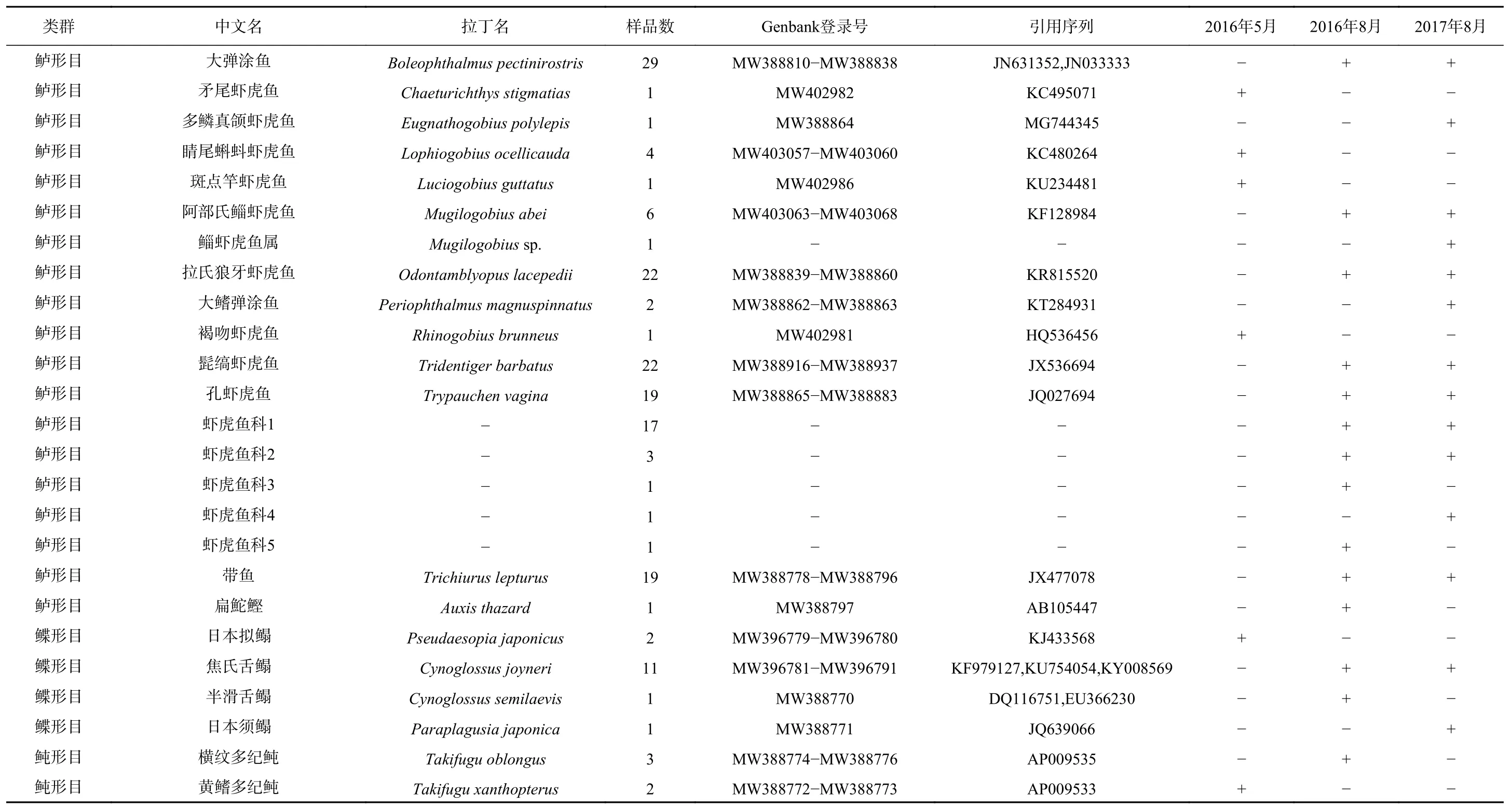
续表1
附录1

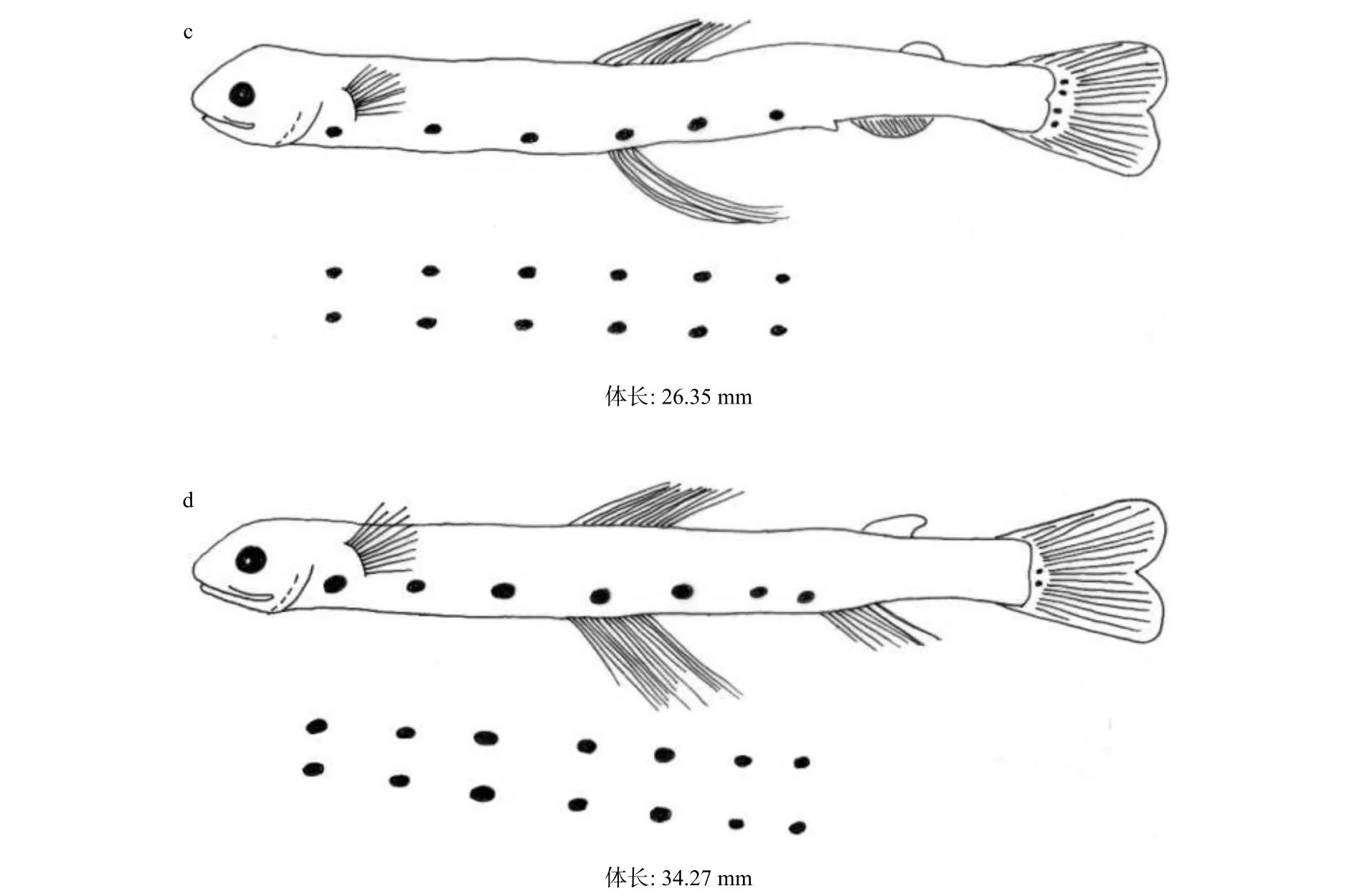
图 A1 龙头鱼仔稚鱼Fig.A1 Larvae and juvenile stages of Harpadon nehereus

图 A2 前鳞龟鰉稚鱼Fig.A2 Juvenile stage of Chelon affinis

图 A3 四指马鲅稚鱼Fig.A3 Juvenile stage of Eleutheronema tetradactylum

图 A4 少鳞鱚稚鱼Fig.A4 Juvenile stage of Sillago japonica

图 A5 日本须鳎仔鱼Fig.A5 Larvae stage of Paraplagusia japonica
