低温胁迫下不同咖啡生理生化指标的变化及耐寒性评价
2021-08-26黄丽芳李金芹龙宇宙董云萍王晓阳林兴军孙燕陈鹏闫林
黄丽芳 李金芹 龙宇宙 董云萍 王晓阳 林兴军 孙燕 陈鹏 闫林

摘 要:以咖啡主栽品种大粒种、中粒种和小粒种咖啡为材料,进行8 ℃低温胁迫,研究其在胁迫条件下植株的伤害情况和生理生化指标变化,并利用隶属函数法对3种咖啡进行综合评价。结果表明:低温胁迫条件下,3种咖啡叶片受害程度差异显著,小粒种咖啡比大粒种和中粒种咖啡具有明显的耐寒表型;小粒种咖啡的可溶性糖和可溶性蛋白含量在胁迫72 h时,较大粒种和中粒种的高。3种咖啡的抗氧化酶变化趋势都是先升后降,大粒种和中粒种咖啡的抗氧化酶在各处理间都处于较低的水平,小粒种咖啡在胁迫48、72 h时仍表现活跃;咖啡的丙二醛和相对电导率的变化幅度与其抗寒性呈现负相关。通过隶属函数法综合评价,3种咖啡抗寒性由强到弱排序依次为:小粒种>大粒种>中粒种。
关键词:咖啡;抗寒性;生理生化;隶属函数法
中图分类号:S571.2 文献标识码:A
Changes of Physiological, Biochemical and Evaluation of Cold Tole-rance in Coffea
HUANG Lifang1, LI Jinqin2, LONG Yuzhou1, DONG Yunping1, WANG Xiaoyang1, LIN Xingjun1, SUN Yan1, CHEN Peng1, YAN Lin1*
1. Spice and Beverage Research Institute, Chinese Academy of Tropical Agricultural Sciences / Key Laboratory of Genetic Resources Utilization of Spice and Beverage Crops, Ministry of Agriculture and Rural Affairs / Hainan Provincial Key Laboratory of Genetic Improvement and Quality Regulation for Tropical Spice and Beverage Crops, Wanning, Hainan 571533, China; 2. College of Tropical Crops, Yunnan Agricultural University, Puer, Yunnan 665099, China
Abstract: Coffea liberica, Coffea canephora and Coffea arabica, treated with 8 ℃, were used to study the leaf physiological responses, and the cold tolerance of the cultivars was comprehensively evaluated using membership function. Under low temperature stress, C. arabica showed stronger cold-tolerance than C. liberica and C. canephora, and the damage degree of the leaves was significant. The content of soluble sugar and protein of C. arabica was higer than of that of C. liberica and C. canephora under 8 ℃ for 72 h. The antioxidant enzymes of the cultivars increased first and then decreased. The antioxidant enzymes of C. liberica and C. canephora were in a low level among all treatments, while that of C. arabica was still active under 48 and 72 h stress. Malondialdehyde and relative electrical conductivity was negatively correlated with cold resistance.The Evaluation on the cold tolerance of the cultivars was in the order C. arabica>C. liberica>C. canephora.
Keywords: coffea; cold resistance; physiology and biochemistry; subordinate function (SF)
DOI: 10.3969/j.issn.1000-2561.2021.07.017
咖啡起源于非洲中北部熱带亚热带地区,为茜草科(Rubiaceae)咖啡属(Coffea)多年生常绿灌木或小乔木,为世界三大饮料作物(咖啡、茶叶、可可)之一。咖啡属有124种,其中商业栽培的主要有阿拉比卡咖啡(Coffea arabica L.,俗称小粒种咖啡)、罗布斯塔咖啡(Coffea Canephora Pierre,俗称中粒种咖啡)和利比利卡咖啡(C. liberica Bull ex Hiern,俗称大粒种咖啡)。咖啡富含可溶性蛋白、脂类、蛋白质、糖类、天然物质和天然解毒物,研究发现咖啡主要成分咖啡因、绿原酸、阿魏酸等,有阻止因高脂食物导致的体重增加、记忆障碍认知损伤[1]、预防老年痴呆阻止细胞死亡[2]和有效抵抗抑郁症[3]的功效。咖啡树种植2~3年左右就会有一定的收获,管理成本较低,总体产值高[4],是我国重要的热带经济作物之一,在国内具有不可替代的特色和优势,对促进热区政府税收、企业增效和农民增收发挥着重要的作用[5]。
温度是限制植物生长发育的主要因素之一,也是限制咖啡向高纬度、高海拔推广的主要限制因子。较高海拔种植的咖啡树病虫害轻,咖啡果发育期变长,咖啡品质提高[6-7],但高海拔地区霜冻时有发生,咖啡树易受寒害。咖啡适宜生长在平均温度18 ℃~25 ℃气候环境下,最适宜花芽发育温度为16 ℃~20 ℃,气温过低不利于开花或授粉,气温过高不利于正常开花,花量少进而导致结果少[8-9]。目前关于烟草[10]、玉米[11]、茶树[12]等的抗寒性研究相继报道,但是咖啡抗寒方面的研究却很少,也没有选育出高抗寒性的咖啡品種,抑制了咖啡产业的发展[13]。国外主要从种质资源的生理生化方面进行了初步的抗寒鉴定。Ramalho等[14-15]研究了冷胁迫条件下不同基因型咖啡的矿物质含量动态变化,以及不同冷胁迫时期咖啡植株的氧化应激反应等。Fortunato等[16]以小粒种和中粒种以及2份杂交种的咖啡苗为材料,研究了在冷条件下抗氧化系统的生物化学及分子特性,结果表明,中小粒种杂交种的冷驯化能力最强,小粒种次之。国内学者研究了咖啡在受寒时的防寒御寒措施,以及寒害后的防治方法[17-19]。然而,关于咖啡种质资源抗寒性鉴定,低温胁迫对咖啡苗生理生化的影响尚未见报道。因此,本研究选取生产上常见的3个不同种的咖啡作为试材,研究其在低温胁迫下生理生化指标的变化,利用隶属函数法综合评价其抗寒性,为咖啡抗寒品种的选择提供理论依据,也为进一步探索咖啡的抗寒机理提供基础。
1 材料与方法
1.1 材料
供试材料为1年生大粒种、中粒种和小粒种3种咖啡苗,株高50.0~60.0 cm,茎粗约1.0 cm,分枝对数2对。小粒种咖啡为云南省主栽品种‘卡蒂姆(CIFC7963),中粒种咖啡为海南省主栽品种‘热研4号咖啡,大粒种咖啡为‘大韩,材料均来源于中国热带农业科学院香料饮料研究所,材料统一栽植在泥炭土和沙土混合基质的塑料盆中,选取生长正常,无病虫害和机械损伤的苗木放入人工培养箱中,每种咖啡各选取15株,设置3个重复,每个重复5株,进行低温胁迫处理。
1.2 方法
试验设置8 ℃低温胁迫处理,以25 ℃为对照(CK),光照强度为10 000 lx,光周期为昼夜12 h/12 h,相对湿度保持在70%左右。分别在胁迫12、24、36、48、72 h时采集叶片并迅速置于液氮中冷冻,–80 ℃冰箱保存,用于测定叶片生理指标;各处理3次重复。
生理指标测定:采用南京建成生物技术研究所测定试剂盒分别测定叶片的过氧化物酶(POD)、总超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、脯氨酸(Pro)、可溶性糖、可溶性蛋白和丙二醛(MDA)含量等生理生化指标,具体操作参照说明书,每个处理测定3次重复。
相对电导率测定:选择8 ℃处理0、24、48、72 h后的第三或第四片完全展开叶片,避开主脉剪成0.2 cm长宽的碎片,称取0.2 g碎片加入20 mL去离子水,抽真空,放入三角瓶中并盖上封口膜,在室温下振荡8 h,振荡频率80 r/h。采用DDS-11A型电导仪测定浸泡液的电导率R1,再将盛有浸泡液的三角瓶置于沸水浴30 min,冷却至室温,再测定浸泡液的电导率R2,3次重复。相对电导率按以下公式计算:
相对电导率R=(R1/R2)×100%。
1.3 数据处理
试验数据采用SPSS 25.0软件进行统计分析,生理生化指标用LSD法进行多重比较。采用模糊综合评判方法中的隶属度公式综合评价不同咖啡品种的抗寒性。参照陈明辉等[20]和何雪银等[21]的方法计算隶属函数值和平均隶属度,分析评价不同咖啡品种抗寒性。其中,与抗寒性呈正相关指标用公式(1)计算,与抗寒性呈负相关指标用公式(2)计算:
隶属函数=(Xi–Xmin)/(Xmax–Xmin) (1)
反隶属函数=1–(Xi–Xmin)/(Xmax–Xmin) (2)
公式中,Xi为指标测定值,Xmin、Xmax为该指标内的最小值和最大值。隶属函数平均隶属度值越大,则抗寒性越强。
2 结果与分析
2.1 低温胁迫对咖啡叶片表型的影响
由图1可以看出,通过观察咖啡叶片的寒害表型发现,8 ℃低温处理对不同咖啡品种叶片具
有差异性,在处理12 h时3种品种咖啡叶片受害较轻,主要表现为叶片轻度受害,出现萎焉性状,大粒种叶片在处理24 h时受害较其他2个品种要严重,其叶片局部变褐;处理36 h时,大粒种咖啡和小粒种咖啡叶肉组织轻微褐化,中粒种叶片从主脉附近变褐,叶片轻度失水;处理48 h和72 h时,大粒种叶片褐化程度加重、叶片严重失水,出现卷曲、发黄;中粒种咖啡从80%的叶片褐化到整片褐化,失水严重,变干;小粒种咖啡叶片60%中度褐化,轻微的失水。从8 ℃处理对3种咖啡叶片的表型可以看出小粒种咖啡叶片受害较轻,中粒种和大粒种咖啡叶片受害较重。
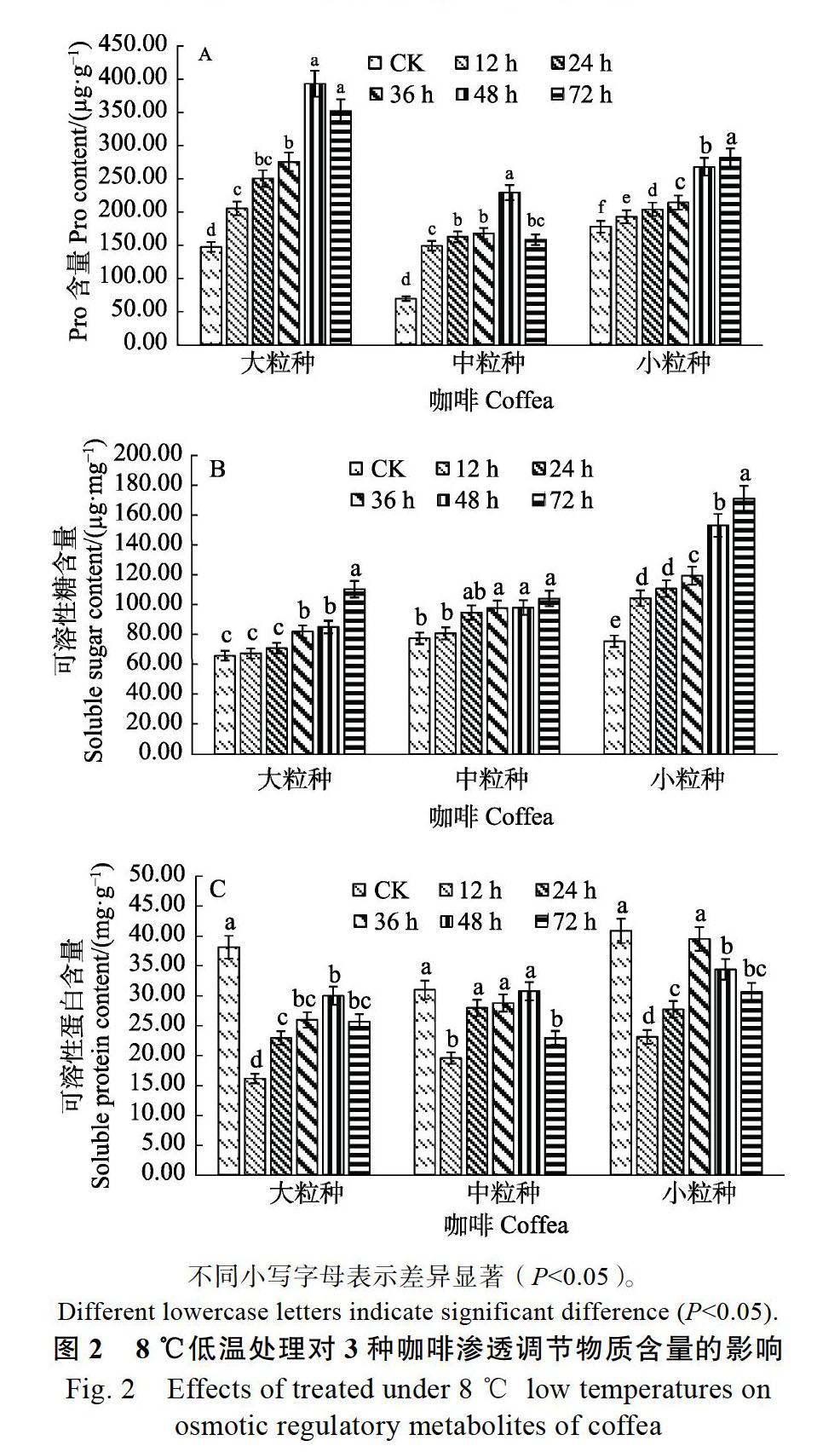
2.2 低温胁迫对咖啡叶片渗透调节物质含量的影响
由图2A可知,经8 ℃低温胁迫后,3种咖啡品种叶片Pro含量均显著性升高,随着低温胁迫
时间的延长其变化趋势和幅度均不相同,大粒种和中粒种咖啡Pro含量的变化趋势是先升后降,均在48 h时达到393.17和228.79的峰值,与CK相比,差异均达显著(P<0.05),而小粒种咖啡的变化趋势为持续升高,处理组均与CK差异显著(P<0.05);在处理72 h时,3个品种较CK相比分别增加了1.36、1.29和0.58倍,可以看出,大粒种和中粒种在受到胁迫后会产生大量的Pro来平衡细胞代谢,但随着时间的延长,植物受害较为严重,导致Pro含量的合成受到影响。
低温胁迫对3种咖啡叶片可溶性糖含量的测定结果见图2B,随着8 ℃处理时间的延长,3种咖啡的可溶性糖含量均呈上升趋势,在72 h时达到最大值,与CK相比均呈显著状态(P<0.05)。从3种咖啡的可溶性糖含量的上升幅度中可以看出,小粒种咖啡在遇到低温胁迫后,能够产生较多的糖来提高叶片细胞液的渗透浓度从而起到降低冰点提高抗寒性的作用。大粒种和中粒种咖啡在处理12 h和24 h时与CK差异不显著,说明在低温胁迫过程中,这2种咖啡合成可溶性糖的速度较慢,使得植株受害较为严重;大粒种、中粒种和小粒种咖啡在8 ℃处理72 h的可溶性糖含量较CK相比,分别上升了67.55%、34.42%和126.23%,说明小粒种咖啡具有较强的抗寒性。
图2C可溶性蛋白含量变化表明,3种咖啡在低温胁迫过程中的可溶性蛋白含量趋势的变化趋势相同,均是先下降后上升再下降,处理12 h时都为最低值,大粒种和中粒种在48 h时达第二峰值,小粒种在36 h时达到第二峰值,说明此时对3种咖啡造成严重胁迫,超过咖啡叶片可溶性蛋白含量的调节能力;大粒种48 h与CK差异显著(P<0.05),中粒种和小粒种咖啡的第二峰值与CK差异不显著,而处理72 h时较CK相比差异均达显著(P<0.05),分别降低了23.73%、25.96%和25.05%。
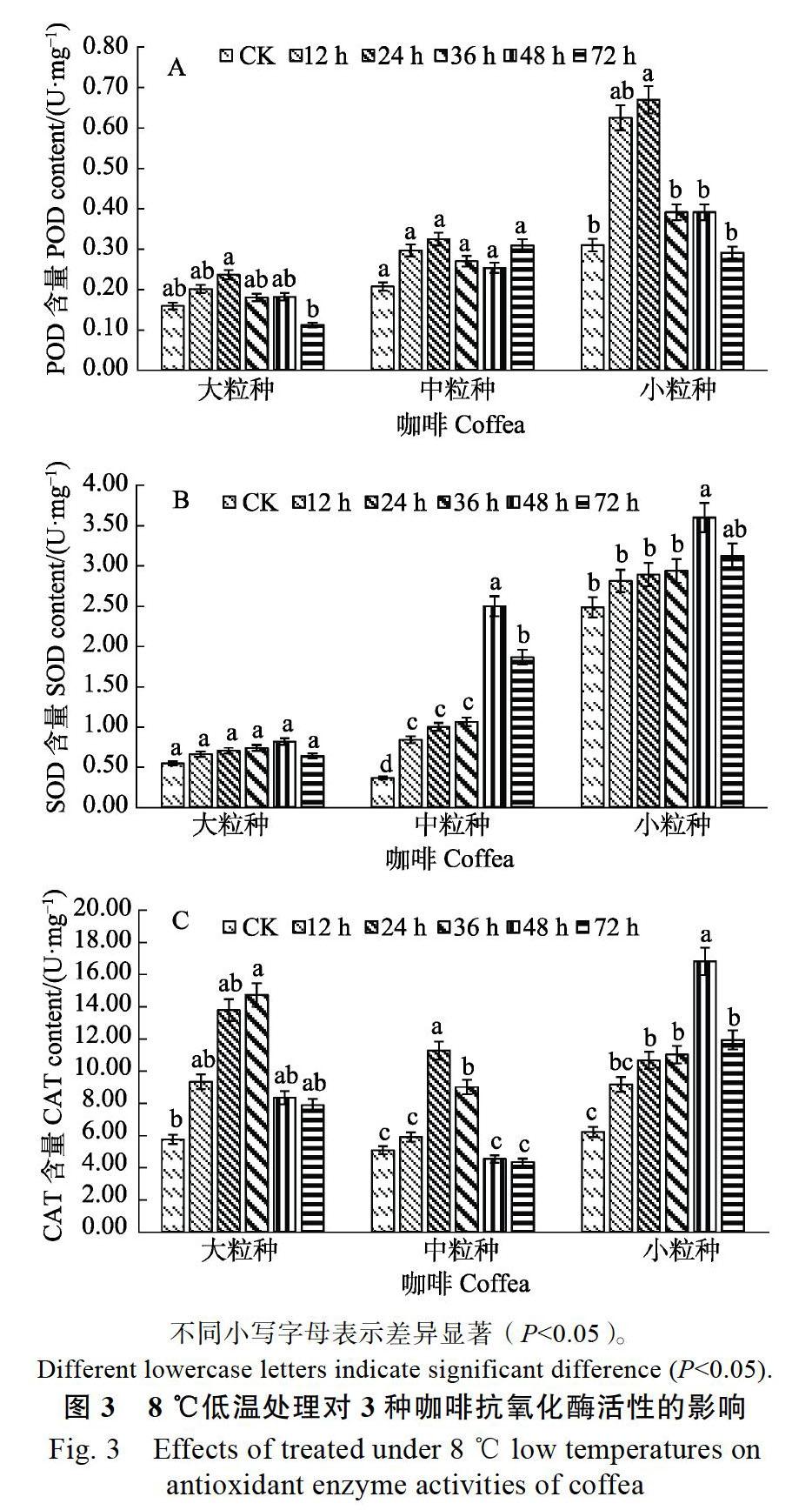
2.3 低温胁迫对咖啡叶片抗氧化酶活性的影响
由图3A可知,8 ℃低温胁迫下POD活性变化可以看出,3种咖啡的变化趋势和变化幅度不同。随着低温处理时间的延长,小粒种咖啡的POD活性急剧增加,在24 h时达峰值0.63(U/m),显著高于CK(P<0.05),比CK提高了1.16倍,大粒种和中粒种咖啡也在24 h时出现峰值,分别比CK提高了0.49、0.57倍,大粒种低温处理24 h时较72 h相比呈显著差异(P<0.05),中粒种咖啡低温处理组较CK相比均未达显著差异。
从图3B可以看出,在8 ℃低温处理下,随着胁迫时间的延长,3种咖啡的SOD活性较对照CK相比均有不同程度的增加,其变化趋势均是先升后降,且都在48 h时达到峰值;在低温胁迫处理时间延长的情况下,中粒种咖啡的变化幅度较大,低温处理组均显著高于CK(P<0.05);小粒种咖啡的变化幅度次之,在低温处理48 h时,SOD含量最高,为3.60 U/mg,与CK达显著差异(P<0.05);大粒种咖啡的变化幅度最小,处理之间差异均不显著。大粒种、中粒种和小粒种SOD活性在低温处理72 h时,较CK相比分别提高了16.80%、415.59%和25.82%。
由图3C可知,在低温处理中,3种咖啡叶片CAT活性均呈先升后降的趋势,大粒种咖啡上升幅度比较大,而中粒种咖啡上升幅度最小。从CAT活性变化中可知,大粒种、中粒种和小粒种咖啡分別在36 h、24 h和48 h时达到峰值,且均显著高于CK(P<0.05),中粒种的峰值较为往前,在低温胁迫后中粒种咖啡产生了大量的CAT来分解低温过程中所产生的过氧化氢(H2O2),随着低温处理时间的延长,H2O2的产生速度超过了中粒种咖啡CAT的合成速度,因此导致CAT活性降低,说明中粒种咖啡的抗寒性相对较弱。小粒种咖啡在低温处理72 h时与CK差异达显著(P<0.05),其他2种咖啡差异均不显著(P< 0.05)。
2.4 低温胁迫对咖啡叶片细胞膜的影响
由图4A可知,经过8 ℃低温胁迫后,3种咖啡叶片MDA含量呈持续上升趋势,其中大粒种咖啡的MDA含量变化幅度比中粒种和小粒种的
变化幅度大,大粒种和中粒种咖啡的所有低温处
理均显著高于CK(P<0.05),低温处理72 h与CK相比,大粒种、中粒种和小粒种分别增加了177.78%、113.40%和76.52%,从上升幅度中可以看出,在8 ℃低温处理下,大粒种咖啡受害情况较为严重,MDA积累较为严重。
相对电导率是反映咖啡叶片在逆境条件下细胞原生质膜透性的重要指标。由图4B可知,随着低温胁迫处理时间的延长,3种咖啡叶片的相对电导率均有不同程度的上升,在处理24 h时,大粒种和中粒种咖啡的增加幅度较大,相对电导率的值为84.57%和67.38%,比CK分别升高了94.55%和101.53%,小粒种的变化幅度较小,比CK升高了17.74%;3种咖啡低温处理72 h的相对电导率与CK差异显著(P<0.05),分别增加了177.68%、170.07%和198.99%。
2.5 应用隶属函数法对3种咖啡抗寒性的综合评价
MDA含量和相对电导率对咖啡品种抗寒性起到反向作用,其值越高,抗寒性越低,用反隶属函数法,而其他指标对咖啡抗寒起正向作用。采用隶属函数法,3种咖啡抗寒指标的平均隶属度,小粒种咖啡的最高为0.54,说明小粒种咖啡的抗寒性较强,大粒种和中粒种咖啡为0.30和0.28,说明这2种咖啡的抗寒性相近,都较弱。抗寒性综合评价:为小粒种>大粒种>中粒种。
3 讨论
脯氨酸(Pro)作为植物细胞体内的渗透调节物质,在植物抵抗低温胁迫时主要起了平衡细胞代谢,促进蛋白质水合,维持细胞运输,调节渗透压等作用[22]。在8 ℃低温过程中,3种咖啡的Pro含量显著上升,其中大粒种和中粒种72 h处理较48 h处理Pro含量略微下降,说明在受到胁迫后会产生大量的Pro来平衡细胞代谢,但随着时间的延长,植物受害较为严重,导致Pro含量的合成受到影响。小粒种咖啡的Pro含量持续上升,处理间差异达到显著水平,说明小粒种咖啡渗透调节能力较强,很大程度的抵御胁迫所带来的危害。可溶性糖和可溶性蛋白是影响植物细胞渗透调节的重要物质,在低温胁迫过程中,可溶性糖和可溶性蛋白含量可以增强细胞保水能力,非结冰水增加,在一定程度上可抵抗低温对细胞造成的损伤[23-24];本研究中3种咖啡的可溶性蛋白和可溶性糖含量均有不同程度的上升,随着处理时间的延长含量越来越高,表明咖啡进行了蛋白质合成和可溶性糖含量的积累来抵御胁迫,其中可溶性蛋白的变化趋势是降-升-降,说明随着处理时间的延长,该物质含量降低,细胞渗透率升高,造成咖啡植株失水。
低温胁迫下植物会产生能够破坏细胞成分如蛋白质、脂类、膜、核酸等的活性氧[25],植株会通过产生POD、SOD、CAT等低分子量抗氧化酶,使得氧自由基维持在一个稳定的水平上,从而阻止活性氧对植物的伤害。本试验中,经过低温胁迫处理后,3种咖啡叶片的POD、SOD、CAT活性呈先升后降趋势,说明低温诱导咖啡抗氧化酶活性的提高,从而缓解低温带来的损害,但随着胁迫作用的不断增强,抗氧化系统遭到破坏,酶活性下降,膜脂过氧化程度加大[26]。与POD和CAT相比,咖啡SOD防御系统低温耐受时间更长,在低温过程中,3种咖啡的SOD活性均在处理48 h时达到峰值,POD活性及中粒种咖啡CAT活性在24 h时达到峰值,中粒种的POD和CAT活性峰值较为往前,说明在低温胁迫前期中粒种咖啡产生了大量的保护酶来抵抗低温所带来的影响,随着时间的延长,保护酶系统遭到破坏,酶活性降低,这也说明中粒种咖啡的抗寒性相对较弱。
在逆境条件中植物器官会破坏细胞膜的组成物质和导致细胞膜透性增大,电解质外渗,相对电导率升高,生理代谢功能紊乱,发生膜脂过氧化[27]。丙二醛是膜脂过氧化的主要产物,其含量累积可作为植物受伤害的程度。本研究中通过相对电导率和MDA两个指标来反映叶片受寒害程度。在8 ℃低温处理下,随着处理时间的延长,MDA含量呈显著性上升,在处理24 h时,大粒种和中粒种相对电导率增加幅度较大,这与刘英等[28]和孙凌霄等[29]的研究结果一致;在观察咖啡叶片的寒害表型发现,小粒种植株受寒程度明显低于另外2种咖啡,而此时MDA含量变化幅度较小;大粒种和中粒种MDA含量变化较大,其叶片变褐、失水严重,说明MDA含量能够明显呈现出植株受害情况,大粒种和中粒种叶片受害程度较小粒种咖啡严重。
在低温胁迫过程中,3种咖啡的抗氧化系统和渗透调节物质等均有较显著的差别,抗寒性较弱的大粒种和中粒种咖啡随着低温胁迫持续进行,细胞结构被破坏,植株的抗氧化酶系统和代谢失活,植株抗寒性表现出明显降低趋势。本研究得出的咖啡抗寒性渗透物质含量、抗氧化酶活性、丙二醛及相对电导率变化应答规律,以及利用隶属函数法综合评价咖啡的抗寒性,为后续开展咖啡抗寒品种的培育及抗寒基因的挖掘等提供可靠的依据。
参考文献
[1] Moy G A, McNay E C. Caffeine prevents weight gain and cognitive impairment caused by a high-fat diet while elevating hippocampal BDNF[J]. Physiology Behavior, 2013(109): 69-74.
[2] Chu Y F, Chang W H, Richard M B, et al. Crude caffeine reduces memory impairment and amyloid β1-42 levels in an Alzheimers mouse model[J]. Food Chemistry, 2012, 135(3): 2095-2102.
[3] 周莉娜, 南 彩, 王惠玲, 等. 咖啡主要成分在抑郁癥中的研究进展[J]. 国际精神病学杂志, 2018, 45(1): 13-14, 25.
[4] 崔 莹. 生态咖啡种植管理与卫生安全管理分析[J]. 现代园艺, 2020, 43(9): 214-215.
[5] 黄家雄, 黄 琳, 吕玉兰, 等. 中国咖啡产业发展前景分析[J]. 云南农业科技, 2018(6): 4-7.
[6] 萧自位, 张洪波, 田素梅, 等. 云南咖啡生豆品质研究[J]. 食品工业, 2019, 40(2): 29-33.
[7] 黄家雄, 吕玉兰, 程金焕, 等. 不同海拔对小粒种咖啡品质影响的研究[J]. 热带农业科学, 2012, 32(8): 4-7.
[8] 吴家耀. 咖啡花的形态学研究–影响咖啡开花的因素[J]. 云南教育学院学报, 1996(2): 74-77.
[9] 王 芳, 李峪曦, 蒋路平, 等. 低温胁迫下植物分子响应机制研究进展[J]. 世界林业研究, 2020, 33(6): 15-21.
[10] 张嘉雯, 卢绍浩, 赵 喆, 等. 外源褪黑素对低温胁迫下烟草幼苗生理指标的影响[J]. 中国农业科技导报, 2020, 22(9): 78-86.
[11] 侯 雯, 杜 卓, 王 丽, 等. 外源褪黑素对低温胁迫下玉米幼苗生长和生理特性的影响[J]. 中国糖料, 2020, 42(2): 33-37.
[12] 徐义群, 胡 哲, 胡月朦, 等. 干旱-低温交叉适应性对茶树抗寒性的影响[J]. 安徽农业大学学报, 2020, 47(1): 1-6.
[13] 黄家雄, 李贵平. 中国咖啡遗传育种研究进展[J]. 西南农业学报, 2008(4): 1178-1181.
[14] Ramalho J C, Fortunato A S, Goulao L F, et al. Cold-induced changes in mineral content in leaves of Coffea spp. identification of descriptors for tolerance assessment [J]. Biologia Plantarum, 2013, 57(3): 495-506.
[15] Ramalho J C, DaMatta F M, Rodrigues A P, et al. Cold impact and acclimation response of Coffea spp. plants[J]. Theoretical and Experimental Plant Physiology, 2014, 26(1): 5-18.
[16] Fortunato A S, Lidon F C, Santos P B, et al. Biochemical and molecular characterization of the antioxidative system of Coffea sp. under cold conditions in genotypes with contrasting tolerance[J]. Journal of Plant Physiology, 2010, 167(5): 333-342.
[17] 周 华, 李锦红, 杨世华, 等. 小粒种咖啡寒害树截干复壮试验初报[J]. 热带农业科技, 2004, 27(4): 13-16.
[18] 何 麗. 云南咖啡防寒御寒措施[J]. 重庆工商大学学报(自然科学版), 2006, 23(2): 154-156.
[19] 孙 燕,董云萍. 咖啡寒害及其防治[J]. 农村实用技术, 2015(7): 36-37.
[20] 陈明辉, 程世平, 张志录, 等. 低温胁迫下不同果蔗品种光合及荧光特性的变化及耐寒性评价[J]. 热带作物学报, 2018, 39(3): 465-471
[21] 何雪银, 文仁来, 吴翠荣, 等. 模糊隶属函数法对玉米苗期抗旱性的分析[J]. 西南农业学报, 2008, 21(1): 52-56.
[22] 欧 欢, 王振磊, 王新建, 等. 不同品种扁桃花蕾抗寒性评价[J]. 干旱区资源与环境, 2018, 32(9): 169-174.
[23] 马媛媛, 肖 霄, 张文娜. 植物低温逆境胁迫研究综述[J]. 安徽农业科学, 2012, 40(12): 7007-7008, 7099.
[24] 张迎辉. 低温胁迫下福建山樱花的生理响应与抗寒基因的表达[D]. 福州: 福建农林大学, 2014.
[25] Hernandez J A, Ferrer M A, Jimenez A, et al. Antioxidant systems and O2.-/H2H2 production in the apoplast of pea leaves. Its relation with salt-induced necrotic lesions in minor veins[J]. Plant Physiology, 2001, 127(3): 817-831.
[26] 曹 慧, 王孝威. 水分胁迫下短枝型苹果叶片膜脂过氧化和保护酶系统的变化[C]//中国园艺学会. 中国园艺学会第九届学术年会论文集, 重庆: 中国科学技术出版社, 2001: 79-82.
[27] 高京草, 王长柱, 王进国, 等. 枣树抗寒性测定方法研究[J]. 西北林学院学报, 2011, 26(5): 72-75.
[28] 刘 英, 曾琪瑶, 曾炳山, 等. 南洋楹无性系抗寒性研究[J]. 中南林业科技大学学报, 2020, 40(5): 7-12.
[29] 孙凌霄, 金晓玲, 胡希军, 等. 广玉兰新品种‘碧翠的抗寒性评价及抗寒性指标筛选[J]. 湖南生态科学学报, 2020, 7(1): 33-39.
责任编辑:白 净
