双瓣茉莉JsCYP71A基因的克隆和真核表达
2021-08-26袁媛刘予涵李建昆万超张月陈清西伍炳华
袁媛 刘予涵 李建昆 万超 张月 陈清西 伍炳华

摘 要:基于雙瓣茉莉[Jasminum sambac (L.) Aiton]的花、叶转录组,筛选了一个在花朵中高表达且与香气释放规律对应的P450基因进行克隆和表达模式分析。利用cDNA末端快速扩增(RACE)技术获得该基因的5?cDNA序列,与转录组中序列进行拼接并验证,显示该基因是一个含1545 bp的开放阅读框、编码514个氨基酸残基的蛋白。此P450蛋白具有特征性的亚铁血红素结合结构域和半胱氨酸残基,与油橄榄、小叶咖啡等其他植物CYP71A氨基酸序列同源性较高,将其命名为JsCYP71A。遗传进化分析显示,该蛋白与拟南芥P450家族中CYP82亚族CYP82G亲缘关系最近。CYP71A和CYP82G多个成员与萜类香气代谢相关,推测JsCYP71A蛋白有相似功能。JsCYP71A在茉莉花白花苞里的表达量最高,在茎中的表达量较低,在叶片中几乎不表达,茉莉花开放过程中,JsCYP71A在夜间11点和1点表达量高,白天表达量低,与萜类香气物质释放呈正相关。JsCYP71A定位于细胞质中。为获得异源表达蛋白,构建真核表达载体,在酵母BY4742株系中表达JsCYP71A,经Western Blot检测发现株系能成功表达大小约60 kDa的蛋白,为后续JsCYP71A酶学功能分析奠定基础。本研究结果为分析JsCYP71A在茉莉花萜类香气合成代谢中的作用提供参考。
关键词:茉莉花;P450;基因克隆;亚细胞定位;真核表达
中图分类号:S685.16 文献标识码:A
Cloning and Eukaryotic Expression of JsCYP71A from Jasminum sambac
YUAN Yuan, LIU Yuhan, LI Jiankun, WAN Chao, ZHANG Yue, CHEN Qingxi, WU Binghua
College of Horticulture, Fujian Agriculture and Forestry University, Fuzhou, Fujian 350002, China
Abstract: Based on the flower and leaf transcriptome of Jasminum sambac, a P450 gene with high expression in flowers and corresponding to the pattern of volatile fragrance release was screened for cloning and expression pattern analysis. The 5?cDNA sequence of this gene was obtained by RACE technology, which was aligned with the sequence in the transcriptome and verified by PCR. JsCYP71A contained an open reading frame at 1545 bp and encoded a protein with 514 amino acid residues. JsCYP71A had characteristic heme binding domains and cysteine residues. JsCYP71A had a high homology with the amino acid sequence of CYP71A in olive, coffee and other plants, therefore this gene was named as JsCYP71A. Phylogenetic analysis showed that JsCYP71A was closely related to CYP82G in the family of Arabidopsis thaliana P450. Multiple members of CYP71A and CYP82G subfamily were related to the aroma metabolism of terpenoids, so it is speculated that JsCYP71A has similar functions. The expression level of JsCYP71A was the highest in the white bud of jasmine, lower in the stem, and very low in the leaf. During the flowering process of jasmine, the expression level of JsCYP71A was high at 11 oclock and 1 oclock at night and low at daytime, which was positively correlated with the release of flower aroma. JsCYP71A was located in the cytoplasm of the cells. Eukaryotic expression vector of JsCYP71A was constructed and expressed in yeast BY4742 strain, and Western Blot showed that a protein about 60 kDa was successfully expressed. The result would lay a foundation for the subsequent enzymatic function analysis of JsCYP71A and for the future analysis of the role of JsCYP71A in the metabolism of terpenoids of jasmine.
Keywords: Jasminum sambac L.; P450; gene cloning; subcellular localization; eukaryotic expression
DOI: 10.3969/j.issn.1000-2561.2021.07.005
植物细胞色素P450是一种加单氧氧化酶,可切割进入底物活性位点并与血红素铁结合的氧分子的双键断裂,将其中的一个氧原子添加到底物的氧化位点,使底物发生氧化[1-2]。植物P450蛋白能结合和催化分子体积不同、结构多样的底物,并能催化多种类型的化学反应,如烷基的羟化、烯基的环氧化、烃基的氧化、氮/硫/氧位的脱烷基化、氮位的羟化和氧化、硫位的氧化、氧化性脱氨、氧化性的碳碳键断裂等,因而也被称为万能的生物催化剂[3]。P450蛋白广泛参与植物特化化合物,如萜类、苯丙烷类及含氮化合物的合成和代谢过程[4],对这些次生代谢产物的结构多样性起关键作用[5]。苯丙烷的生物合成途径中,己报道的P450蛋白超过16个,如肉桂酸-4-羟化酶(C4H,CYP73)、类黄酮-3?-羟化酶(F3?H,CYP73B)、類黄酮-3?,5?-羟化酶(F3?5?H,CYP75A)、2-羟基异黄酮酶(IFS,CYP93C)、松柏醛-5-羟化酶(coniferylalcohol 5-hydroxylase,CYP84)等[1];萜类化合物生物合成中单萜(monoterpene)、倍半萜(sesquiterpenes)、双萜(diterpenes)、三萜(triterpenoids)等都可成为P450蛋白的底物,如拟南芥中CYP71A和CYP71R专一性地参与了单萜的代谢,CYP71AV、CYP71BA、CYP71BL、CYP706B参与倍半萜代谢,CYP71Z、CYP76M、CYP99A、CYP720B参与双萜代谢,CYP708A和CYP716A参与三萜代谢[1]。棕桐酸、油酸、亚油酸等长链脂肪酸的合成也有P450蛋白参与进行羟化反应[2, 6],表明P450蛋白在植物特化代谢中尤其是萜烯类、苯丙烷类的合成与转化的重要作用正在模式植物中得到逐步明晰。
茉莉花[Jasminum sambac (L.) Aiton]为福州市市花,属于木犀科素馨属灌木,原产于南亚和中亚地区[7]。茉莉花是福州传统名茶“茉莉花茶”窨制的香原[8],同时也是香料工业提取精油浸膏的重要原料[9]。茉莉花香气物质主要为单萜、倍半萜、苯丙烷/苯环类(benzenoids/phenylpropanoids)和其他微量物质[10-11]。尽管对于茉莉花的香气成分以及个别合成酶、水解酶有过诸多研究[12-13],也报道过栽培环境因素对花香的影响以及不同品种间的成分差异[14-15],但是有关茉莉花香气成分尤其是芳樟醇、乙酸苄酯等优势成分的代谢途径的了解并不全面。本研究基于前期茉莉花、叶转录组数据(GenBank登录号GHOY00000000)[16],获得若干在花朵中高量表达且与香气释放规律一致的P450基因。本研究选择其中一个表达量最高的P450序列(c61299_g1),克隆其编码序列并且分析其时空表达模式,并了解其亚细胞定位并在酵母中进行异源表达,为其功能研究提供参考。
1 材料与方法
1.1 材料
1.1.1 植物材料 以三年生双瓣茉莉花为材料。选择刚开放不久的花朵用于基因编码序列克隆,选择第2天会开放的黄花苞、当天会开放的紧实白花苞、盛开的花朵(开放8 h)、衰老的花朵(开放36 h左右)、嫩叶、老叶、嫩茎等7种组织用于分析基因在不同组织中的表达水平(图1)。分析基因在花朵在开放过程中的表达水平时,从17:00(紧实白花苞)到第3天15:00(花朵衰老发紫),每隔2 h采集花朵。所有样品用剪刀剪下后用铝铂纸包好投入液氮中,然后保存在–80 ℃超低温冰箱中。
1.1.2 表达载体、菌株及培养基 Gateway系统植物表达载体pK7FWG2.0(35 S启动子,GFP报告基因)、真核表达载体pDRTxa、农杆菌(Agrobacterium tunmefaciens)GV3101株系、酵母(Saccharomyces cerevisiae)By4742株系[17]为本实验室保存。感受态大肠杆菌(E. coli)DH5α购自北京全式金生物技术有限公司。
用于酵母培养的SD+KHL液体培养基配置参见伍炳华等[17]方法。SD+KHL固体培养基为液体培养基基础上添加20%(W/V)琼脂粉。
1.2 方法
1.2.1 RNA提取及cDNA获得 称取约0.1 g组织(不同器官组织或不同开放时期花苞和花朵),参照TranZol UP试剂盒(北京全式金生物技术有限公司)方法进行RNA进行提取。使用超微量紫外分光光度计(Nano drop 2000)测定所提取RNA的浓度,并使用电泳检测其完整性。使用EasyScript? One-Step gDNA Removal and cDNA Synthesis Su-perMix试剂盒(北京全式金生物技术有限公司)获得第一链cDNA,反转录体系按说明书要求。用于获得扩增编码序列的cDNA反转录程序为42 ℃ 30 min,85 ℃ 5 s。用于荧光定量的cDNA的反转录程序为42 ℃ 15 min,85 ℃,5 s。
1.2.2 茉莉JsCYP71A编码序列扩增 转录组中需克隆的目标P450基因序列仅为3?端序列,因此采用cDNA末端快速扩增(RACE)方法,根据转录组中P450基因序列,设计RACE引物(表1)扩增其5?端序列。PCR的扩增体系按说明书要求。PCR程序:94 ℃ 30 s,92 ℃ 5 min,5个循环;94 ℃ 30 s,70 ℃ 30 s,72 ℃ 3 min,5个循环;94 ℃ 30 s,65 ℃ 30 s,72 ℃ 3 min,1个循环;94 ℃ 30 s,67 ℃ 30 s,72 ℃ 3 min,25个循环。PCR结束后,将PCR产物使用AxyPrepTM PCR Clean up Kit(Axygen)纯化回收、连接到pEASY-blunt Cloning Vector载体(北京全式金生物技术有限公司),并转化感受态DH5α,菌液PCR检测阳性后送测序。将获得的5?序列与转录组中的序列进行拼接,获得编辑全长序列,根据序列设计引物验证其编辑序列。PCR的扩增体系按说明书要求。PCR程序:94 ℃ 5 min,94 ℃ 30 s,56 ℃ 30 s, 72 ℃ 1 min,35个循环;72 ℃ 10 min。将获得的PCR产物连接克隆载体,转化感受态,菌液PCR检测阳性后送测序(铂尚生物技术有限公司)。
1.2.3 生物信息学分析 将测序之后得到的核酸序列用生物软件网的SMS在线工具(http://www. bio-soft.net/sms/index.html)中的DNA Figures进行序列翻译,Protein Figures进行多重序列比对;运用NCBI中的BLASTX进行P450间的同源性比对;使用NCBI中的Conserved Domain Search Service(CD Search)进行结构域分析;用MEGA4软件分析茉莉花P450基因与其他物种中P450基因的亲缘关系;使用Protparam在线软件,对P450蛋白分子量、等位点(pI)、正负电荷量等数据进行预测。
1.2.4 茉莉JsCYP71A的时空表达特征 根据序列设计特异性引物(表1),采用荧光定量PCR分析JsCYP71A在不同组织和开放期间的表达水平。按照TransStart? Top Green qPCR SuperMix试剂盒体系每个样品做3次生物学重复,每个生物学重复进行3次技术重复。程序为:95 ℃ 3 min;95 ℃ 15 s,60 ℃ 45 s,40个循环。基因相对表达水平使用2–ΔΔCT法[18]进行计算,茉莉花JsActin基因[19]为内参基因。
1.2.5 植物表达载体的构建及转化 为分析JsCYP71A的亚细胞定位,将JsCYP71A构建植物表达载体pK7FWG2.0,设计特异性引物(表1),将JsCYP71A基因的编码序列连接到TOPO入门载体(pENTR/SD/D-TOPO, Invitrogen)连接完成后测序正确的TOPO-JsCYP71A与pK7FWG2.0植物表达载体进行LR反应(Gateway LR Clonase II Enzyme mix, Invitrogen),使得JsCYP71A基因的编码序列置于35S启动子下,并与GFP融合。将测序正确的pK7FGW2.0-JsCYP71A通过冻融法转入农杆菌GV3101株系,PCR检测无误后备用。TOPO反应和LR反应体系及程序参见其说明书。
1.2.6 亚细胞定位分析 使用烟草叶片和茉莉花原生质体分析JsCYP71A的亚细胞定位。使用烟草叶片时,将验证转化成功的阳性克隆菌液使用含相应抗生素的LB液体培养基(含50 mg/L的庆大霉素、利福平、壮大霉素)28 ℃、200 r/min的条件下振荡培养过夜,再取出适量菌液接种于50 mL LB液体培养基中继续培养至OD600值为0.8~1.0。5000 r/min离心10 min收集菌体,加入重悬液(1/2 MS+200 μmol/L乙酰丁香酮+10 mmol/L MES),暗培养3 h用无菌注射器向叶背面轻柔地注入菌液,暗培养24~30 h。使用茉莉花原生质体进行亚细胞定位分析时,茉莉花原生质体的提取和转化方法见前期研究[20]。用激光共聚焦显微镜(Leica TCS SP8)观察判断GFP是否表达,观察GFP时Ex=488 nm,Em=507 nm,觀察叶绿素时Ex=488 nm,Em=681 nm。
1.2.7 茉莉JsCYP71A真核表达载体的构建、转化酵母及蛋白提取 根据pDRTxa载体上的酶切位点,设计合适的带酶切位点的引物(表1),以cDNA为模板扩增JsCYP71A基因编码序列,通过酶切连接的方法连接到pDRTxa载体。连接完成后转化到大肠杆菌DH5α感受态细胞,挑取白斑摇菌后送测序。使用PEG、AcLi和鲑鱼精DNA介导的转化酵母方法[21]将pDRTxa-JsCYP71A转化至酵母株系BY4742。
酵母总蛋白的提取参考伍炳华等[17]的方法。介于P450蛋白的亚细胞定位特点,将总蛋白用Storage buffer(20 mmol/L Tris-Cl,0.1 mmol/L EDTA,10% glycerol,100 mmol/L KCl,1 mmol/L DTT,pH 7.5)稀释到40 mL,100 000 ×g,4 ℃超速离心45 min,获得含有包涵体蛋白沉淀并用100 μL缓冲液和20 ?L蛋白酶抑制剂溶解。
1.2.8 SDS-PAGE电泳及茉莉JsCYP71A蛋白的Western Blot检测 将酵母中提取的总蛋白和包涵体蛋白按照Super-Bradford蛋白定量试剂盒(Bio-Rad)测定蛋白浓度,然后进行SDS-PAGE电泳检测(碧云天生物技术)。蛋白样品10 μg,Marker 6 ?L,在10 V,30 mA条件下进行电泳。SDS-PAGE电泳结束后切除浓缩胶,将获得的胶进行染色、脱色,观察考马斯亮蓝染色结果。在凝胶的适当位置做好标记,置于转移Buffer(2.9 g/L甘氨酸,5.8 g/L Tris,0.7 g/LSDS,20%甲醇)中。剪裁与胶大小相同的6张滤纸和1张膜(3 M),在转移槽中从下往上依次放入3张滤纸、膜、胶、3张滤纸,每层之间不能有气泡,在0.8 mA/cm下转移蛋白1.5 h。转膜完成后,取出膜做好标记,用去离子水漂洗1次、TTBS(8.8 g/L NaCl,20 mmol/L Tris-HCl(pH 8.0),0.05% Tween-20)漂洗3次(每次5 min,水平摇床)。将膜依次通过漂洗、封闭、结合一抗(anti-his mouse monoclonal antibody)、结合二抗(goat anti-mouse IgG(H+L),HRP Conju-gate)、漂洗等几个流程[17]。在洗好的膜上均匀涂布200 ?L的显影液放入显影仪器(Bio- rad,molecular imager? ChemiDoc TM XRS+)中进行显影。
2 结果与分析
2.1 茉莉JsCYP71A基因CDS序列克隆的结果
通过RACE技术得到茉莉花此P450序列的5′RACE片段600 bp(图2A),将这段序列与已有序列进行拼接,得到起始密码子和终止密码子,根据起始和终止密码子位置设计扩增ORF的引物,扩增获得1545 bp的片段(图2B)。
此序列的CDS长度为1545 bp,编码514个氨基酸残基(图3)。用Protparam在线软件预测P450基因分子总量为58.5 kDa,等电点为6.75,分子式为C2675H4193N683O748S21。正负电氨基酸的残基数相差不大,带正电的氨基酸64个,带负电的氨基酸65个。其不稳定系数为44.52,属于不稳定蛋白。
2.2 茉莉JsCYP71A基因序列分析
将茉莉JsCYP71A基因P450序列编码的氨基酸序列经Blast比对(表2),发现其与油橄榄、小果咖啡和苦瓜中的cytochrome P450 71A1-like蛋白,以及大麻和河岸葡萄中TMTT合成酶(trimethyltridecatetraene synthase)的相似度较高,将此基因命名为JsCYP71A。将茉莉花JsCYP71A氨基酸序列与拟南芥中P450序列进行的遗传进化分析(图4),发现JsCYP71A与拟南芥CYP82家族最近,其中与CYP82G1亲缘关系最近。
将茉莉花JsCYP71A蛋白和其他物种中相应蛋白进行多重序列比对,发现序列中的42~501 aa为P450超级家族(图5A)。此外,序列含有P450蛋白典型的脯氨酸富集区、Helix C、Helix K、Meander 以及Heme-binding 结构域(图5B)。
2.3 茉莉JsCYP71A基因的空间和时间表达特征
采用实时荧光定量PCR分析了茉莉JsCYP71A基因在不同组织中表达水平。结果显示,JsCYP71A基因在白花苞中的相对表达量最高,达到38.39,其次是衰老的花朵,再其次为嫩茎和老叶,在黄花蕾和嫩叶中的表达量最低,其中JsCYP71A基因在黄花苞的表达量仅有0.13(图6)。
在花朵开放释香过程中,JsCYP71A基因在夜间(如第1天的21:00—3:00,第2天的19:00— 23:00)表达量高,在白天(如第1天03:00— 17:00,第2天01:00—15:00)表达量低。第1天19:00—23:00这段时间为P450表达量的增长期,其相对表达量在首日晚上23:00点之后达到最高,为120.74,随后开始下降,然后保持低表达量,第2天19:00又开始升高,在21:00左右达到第2次高峰,为101.43,而后开始下降,之后维持较低水平(图7)。
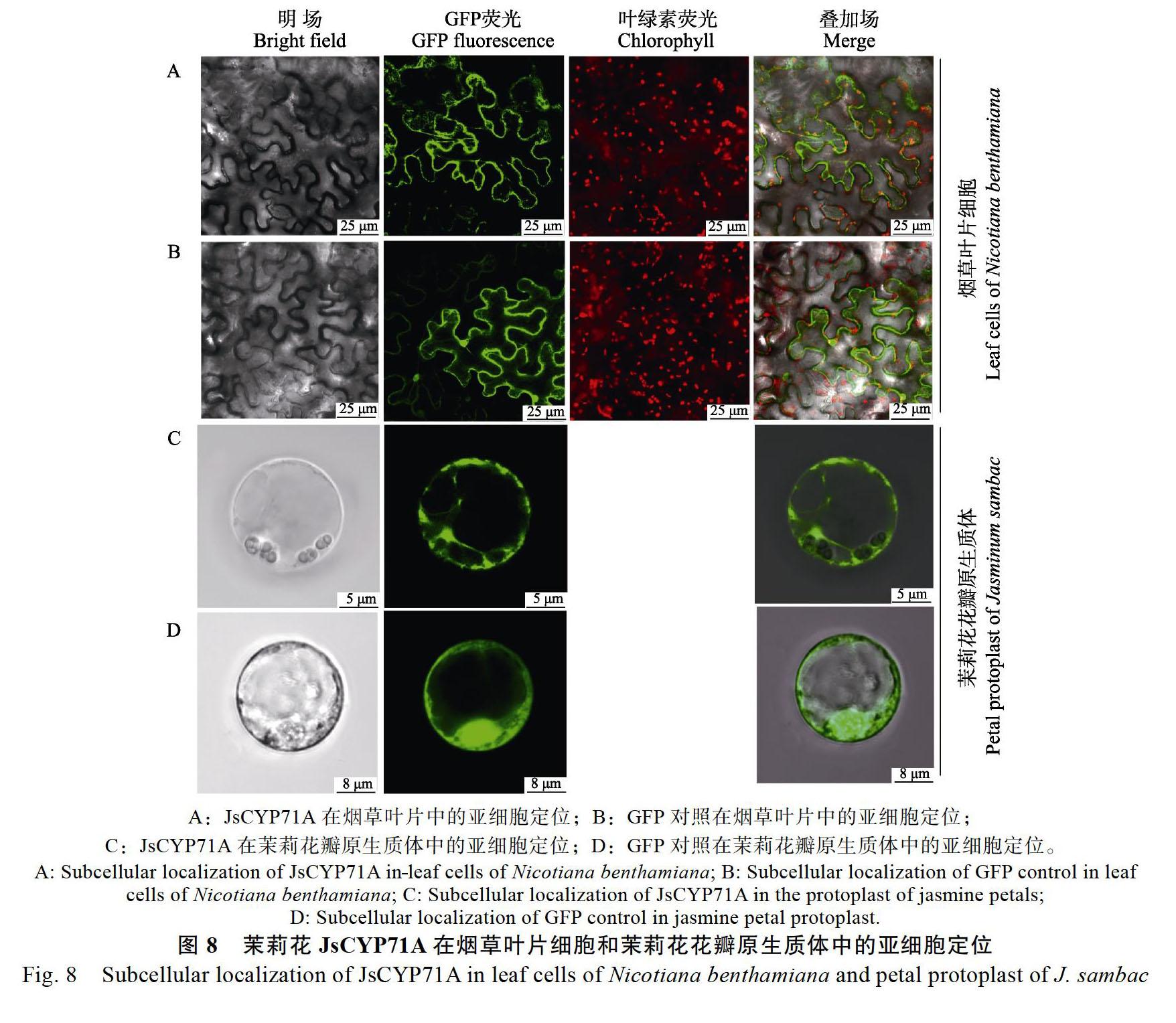
2.4 茉莉JsCYP71A的亚细胞定位
从图8可知,根据烟草叶片中荧光的位置可以判断JsCYP71A在细胞质中表达未在叶绿体中表达,与GFP对照相比,可发现JsCYP71A未在细胞核中表达,即在细胞质中表达;进一步在茉莉花原生质体中进行验证可发现,JsCYP71A的亚细胞定位区域为细胞质中,因此茉莉JsCYP71A的亚细胞定位为细胞质。
2.5 茉莉JsCYP71A在酵母中的表达
根据BSA标准曲线(Y=2.9031X-0.1064,X为酶标仪测定的数据,Y为稀释的BSA标准品浓度)换算转化了JsCYP71A基因以及空载体后酵母中分离的包涵体蛋白浓度分别为0.820 ?g/?L和1.255 ?g/?L。
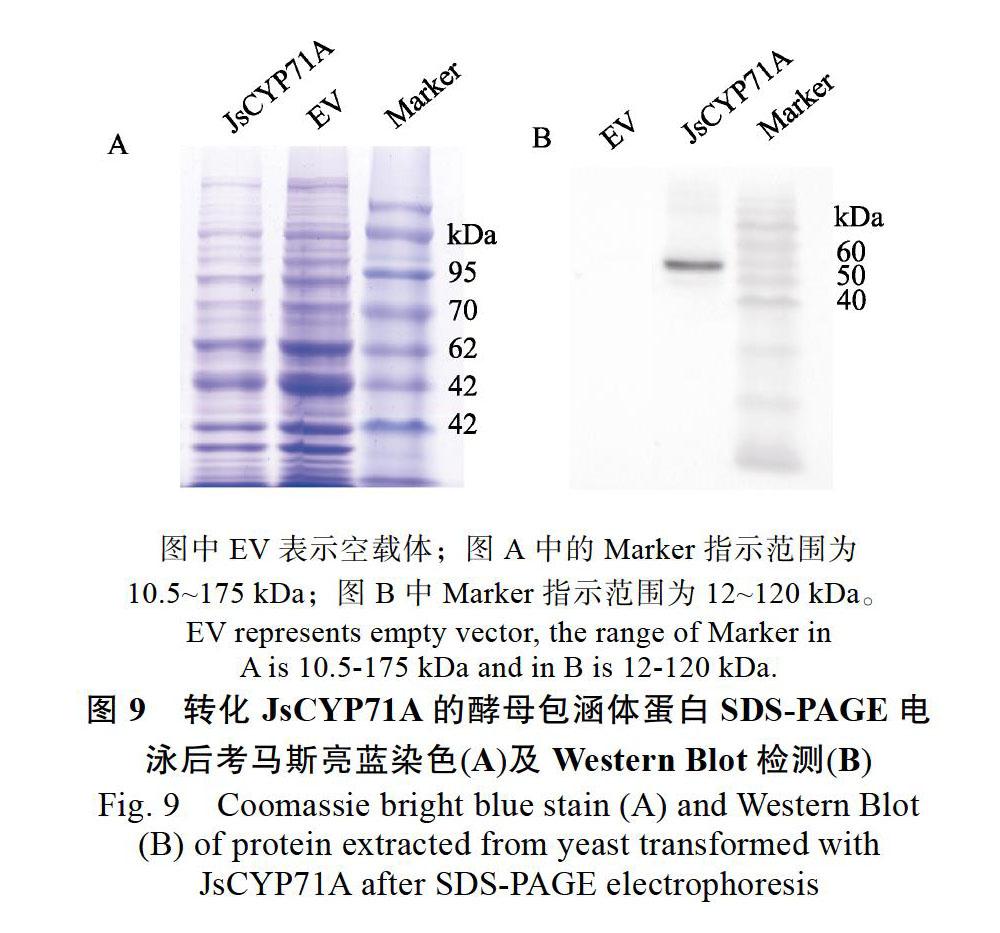
SDS-PAGE胶体考马斯亮蓝染色结果显示从转化了P450的酵母菌液中提取的包涵体蛋白条带均匀分散,条带清晰,表明总蛋白和包涵体蛋白成功得到提取并在凝胶中进行了很好的分离(图9A)。
JsCYP71A蛋白通过生物信息学分析大小为59.39 kDa(包含6个His),Western Blot结果显示,转化了JsCYP71A基因的酵母的总蛋白中在60 kDa处附近出现了条带(图9B),表明提取的总蛋白中确实含有目标JsCYP71A蛋白,即JsCYP71A在酵母中成功表达。
3 讨论
细胞色素P450是高等植物中最大的酶蛋白家族[1],动物中的P450成员一般少于100,但开花植物P450家族庞大,一般超过250個,如在拟南芥(Arabidopsis thaliana)为245个、番茄(Lycopersicon esculentum)457个、水稻(Oryza sativa)343个、葡萄(Vitis vinifera)416个[1]。本研究中,JsCYP71A基因长度1545 bp,编码514个氨基酸,氨基酸序列中有P450超级家族特征序列和脯氨酸富集区、Helix C、Helix K、Meander 以及Heme-binding 结构域基序,初步说明本研究克隆的这个基因是P450基因。
JsCYP71A与同一科植物油橄榄CYP71A相似性和同源性最高。CYP71亚族很多成员参与萜类物质的合成,如CYP71D和CYP71A成员参与薄荷属植物单萜柠檬烯形成后各衍生物的合成[22-24],CYP71B参与了葡萄莎草酮(rotundone)的氧化合成[25],烟草CYP71D参与倍半萜capsidol的合成[26-27]等。此外,JsCYP71A与大麻和河岸葡萄中TMTT(一种半萜)合成酶的相似度较高,TMTT和DMNT是一种在植物防御中十分重要的半萜,在棉花[28]、拟南芥[29-30]、玉米[31]等重要作物中均有报道,这些结果均暗示茉莉JsCYP71A可能具有相似的参与萜类代谢的功能。进一步的遗传进化分析结果表明JsCYP71A与拟南芥CYP82G1亲缘关系最近,拟南芥CYP82G1催化橙花叔醇与香叶基芳樟醇分别转化为挥发性的萜烯同系物DMNT(C11)与TMNT(C16),这二者均为夜间开花的花香物质,也能影响外界刺激调控半萜的合成[29-30]。进一步提示JsCYP71A可能参与了萜类的合成。
茉莉花在夜间开放释香,白天香气较夜间更弱[32],我们分析了JsCYP71A基因在花朵开放48 h过程中的表达水平变化,发现JsCYP71A基因在夜间花朵开放时表达量急速增高,而白天表达低,与茉莉花香气以及萜类香气的释放规律一致[32-33],此外,JsCYP71A基因在当晚会开放的白花苞中表达量最高,在茎、叶等营养器官中的表达量较低,由于花苞开放过程是释放香气物质的主要时期,结合生物信息学分析,推测JsCYP71A可能參与了茉莉花朵开放过程中花朵香气的合成及释放过程。
茉莉花香气的主要组成分为芳樟醇、法呢烯、乙酸芳香酯、甲苯酸甲酯苄酯,其中芳樟醇属于单萜类化合物,占茉莉花单萜类香气物质总含量的90%以上[12],是茉莉花的特征香气组成[34],其含量的变化对茉莉花香气和精油的品质有重要影响。拟南芥中的研究表明芳樟醇生成后,可通过氧化反应生成各类环吡喃型(cyclic pyranoid)或呋喃型化合物的氧化衍生物(furanoid oxygenated derivatives),这些物质可分为挥发性和溶解性两类,其中芳樟醇及其挥发性氧化物是香气的主要成分[35],另一大部分溶解性芳樟醇氧化物以糖苷轭合物的形式存在于植物组织内[36]。这些反应的催化酶属于细胞色素P450家族,包括CYP76C1、CYP76C2、CYP76C3、CYP76C4、CYP71B31[35, 37-38]。相比之下,关于P450在茉莉花萜类香气,特别是芳樟醇合成后代谢的研究尚较少,本研究中JsCYP71A在茉莉花中具有明显的花特异性,并与芳樟醇这类萜类的释放规律具有一致性,暗示JsCYP71A可能参与了萜类的代谢。
茉莉花JsCYP71A蛋白亚细胞定位在细胞质中,作为一种氧化酶,与大部分已报道的P450基因的亚细胞定位一致[25,28,39],为后期JsCYP71A选择真核表达而非原核表达提供基础。后期我们将JsCYP71A在酵母中表达,分离包涵体蛋白并通过Western Blot进行检测,发现JsCYP71A可通过酵母成功表达,为后期测定JsCYP71A催化特定底物,特别是萜类物质的酶学特征分析奠定基础。
参考文献
[1] Hamberger B, Bak S. Plant P450s as versatile drivers for evolution of species-specific chemical diversity[J]. Philosophical Transactions of the Royal Society B Biological Sciences, 2013, 368(1612): 20120426.
[2] 胡之璧, 贺丽虹, 赵淑娟. 细胞色素P450研究进展[J]. 江苏中医药, 2006, 27(10): 81-84.
[3] Bolwell G P, Bozak K, Zimmerlin A. Plant cytochrome P450[J]. Phytochemistry, 1994, 37(6): 1491-1506.
[4] Guengerich F P, Munro A W. Unusual cytochrome P450 enzymes and reactions[J]. Journal of Biological Chemistry, 2013, 288(24): 17065-17073.
[5] Tanaka Y, Brugliera F. Flower colour and cytochromes P450[J]. Philosophical Transactions of the Royal Society B: Biological Sciences, 2013, 368(1612): 20120432.
[6] 赵 剑, 杨文杰, 朱蔚华. 细胞色素P450与植物的次生代谢[J]. 生命科学, 1999, 11(3): 127-131.
[7] Ye Q P, Jin X Y, Zhu X L, et al. An efficient extraction method for fragrant volatiles from Jasminum sambac (L.) Ait[J]. Journal of Oleo Science, 2015, 64(6): 645-652.
[8] 安会敏, 欧行畅, 熊一帆, 等. 茉莉花茶挥发性成分在窨制过程中的变化研究[J]. 茶叶通讯, 2020, 47(1): 67-74.
[9] 陈梅春, 朱育菁, 刘晓港, 等. 茉莉鲜花[Jasminum sambac (L.) Aiton]香气成分研究[J]. 热带作物学报, 2017, 38(4): 747-751.
[10] Bera P, Kotamreddy J N, Samanta T, et al. Inter-specific variation in headspace scent volatiles composition of four commercially cultivated jasmine flowers[J]. Natural Product Research, 2015, 29(14): 1328-1335.
[11] 张 芊. 双瓣茉莉开花过程中挥发性物质变化及花香基因差异表达分析[D]. 福州: 福建农林大学, 2015.
[12] Bera P, Mukherjee C, Mitra A. Enzymatic production and emission of floral scent volatiles in Jasminum sambac[J]. Plant Science, 2017, 256: 25-38.
[13] 孙 君. 茉莉花香气合成相关酶基因克隆与表达分析[D]. 福州: 福建农林大学, 2014.
[14] 沈邦琼, 罗 明, 邱旭明, 等. 双瓣茉莉花品种对比试验[J]. 农业开发与装备, 2018(5): 100, 91.
[15] 李聪聪, 叶晓青, 邓衍明. 不同温度处理对双瓣茉莉开花的影响[J]. 北方园艺, 2018(4): 103-109.
[16] Wang P, Wei P N, Niu F F, et al. Cloning and functional assessments of floral-expressed sweet transporter genes from Jasminum sambac[J]. International Journal of Molecular Sciences, 2019, 20(16): 4001.
[17] Wu B H, Rambow J, Bock S, et al. Identity of a Plasmodium lactate/H+ symporter structurally unrelated to human transporters[J]. Nature Communications, 2015, 6(1): 569-582.
[18] Schmittgen T D, Livak K J. Analyzing real-time PCR data by the comparative C(T) method[J]. Nature Protocols, 2008, 3(6): 1101-1108.
[19] Lu Y, Liu Z Y, Lyu M L, et al. Characterization of JsWOX1 and JsWOX4 during callus and root induction in the shrub species Jasminum sambac[J]. Plants, 2019, 8(4): 79.
[20] 张 娅, 刘晓锋, 张 婧, 等. 茉莉花原生质体瞬时表达体系的建立及应用[J]. 福建农林大学学报(自然科学版), 2019, 48(6): 727-735.
[21] Gietz R D, Woods R A. Transformation of yeast by lithium acetate/single-stranded carrier DNA/polyethylene glycol method[J]. Methods Enzymology, 2002, 350: 87-96.
[22] Mau C J D, Karp F, Ito M, et al. A candidate cDNA clone for (-)-limonene-7-hydroxylase from Perilla frutescens[J]. Phytochemistry, 2010, 71(4): 373-379.
[23] Lupien S, Karp F, Wildung M, et al. Regiospecific cytochrome P450 limonene hydroxylases from mint (Mentha) species: cDNA isolation, characterization, and functional expression of (-)-4S-limonene-3-hydroxylase and (-)-4S-lim?onene-6-hydroxylase[J]. Archives of Biochemistry and Biophysics, 1999, 368(1): 181-192.
[24] Schalk M, Croteau R. A single amino acid substitution (F363I) converts the regiochemistry of the spearmint (?)-limonene hydroxylase from a C6- to a C3-hydroxylase[J]. Proceedings of the National Academy of Sciences, 2000, 97(22): 11948-11953.
[25] Takase H, Sasaki K, Shinmori H, et al. Cytochrome P450 CYP71BE5 in grapevine (Vitis vinifera) catalyzes the formation of the spicy aroma compound (-)-rotundone[J]. Journal of Experimental Botany, 2016, 67(3): 787-798.
[26] Greenhagen B T, Griggs P, Takahashi S, et al. Probing sesquiterpene hydroxylase activities in a coupled assay with terpene synthases[J]. Archives of Biochemistry & Biophysics, 2003, 409(2): 385-394.
[27] H?fer R, Dong L, André F, et al. Geraniol hydroxylase and hydroxygeraniol oxidase activities of the CYP76 family of cytochrome P450 enzymes and potential for engineering the early steps of the (seco)iridoid pathway[J]. Metab Eng, 2013, 20: 221-232.
[28] Liu D, Huang X, Jing W, et al. Identification and functional analysis of two P450 enzymes of Gossypium hirsutum involved in DMNT and TMTT biosynthesis[J]. Plant Biotechnology Journal, 2018, 16(2): 581-590.
[29] Lee S, Badieyan S, Bevan D R, et al. Herbivore-induced and floral homoterpene volatiles are biosynthesized by a single P450 enzyme (CYP82G1) in Arabidopsis[J]. Proceedings of the National Academy of Sciences of the United States of America, 2010, 107(49): 21205-21210.
[30] Rajarammohan S, Pradhan A K, Pental D, et al. Genome-wide association mapping in Arabidopsis identifies novel genes underlying quantitative disease resistance to Alternaria brassicae[J]. Molecular Plant Pathology, 2018, 19(7): 1719-1732.
[31] Richter A, Schaff C, Zhang Z, et al. Characterization of biosynthetic pathways for the production of the volatile homoterpenes DMNT and TMTT in Zea mays[J]. Plant Cell, 2016, 28(10): 2651-2665.
[32] 赵国飞, 罗理勇, 常 睿, 等. 离体茉莉花释香过程中的香气成分特征[J]. 食品科学, 2015, 36(18): 120-126.
[33] 李 鹤, 俞 滢, 陈桂信, 等. 双瓣茉莉花 [Jasminum sambac (L.) Ait]开放过程香气成分的动态变化[J]. 茶叶学报, 2015, 56(1): 29-38.
[34] Khidzir K M, Cheng S F, Chuah C H. Interspecies variation of chemical constituents and antioxidant capacity of extracts from Jasminum sambac and Jasminum multiflorum grown in Malaysia[J]. Industrial Crops and Products, 2015, 74: 635-641.
[35] Boachon B, Junker R R, Miesch L, et al. CYP76C1 (Cytochrome P450)-mediated linalool metabolism and the formation of volatile and soluble linalool oxides in Arabidopsis flowers: a strategy for defense against floral antagonists[J]. Plant Cell, 2015, 27(10): 2972-2990.
[36] Flath R A, Light D M, Jang E B, et al. Headspace examination of volatile emissions from ripening Papaya (Carica papaya L., Solo variety)[J]. Journal of Agricultural and Food Chemistry, 1990, 38(4): 1060-1063.
[37] Ginglinger J F, Boachon B, H?fer R, et al. Gene coexpression analysis reveals complex metabolism of the monoterpene alcohol linalool in Arabidopsis flowers[J]. The Plant Cell, 2013, 25(11): 4640-4657.
[38] H?fer R, Boachon B, Renault H, et al. Dual function of the cytochrome P450 CYP76 family from Arabidopsis thaliana in the metabolism of monoterpenols and phenylurea herbicides[J]. Plant Physiology, 2014, 166(3): 1149-1161.
[39] [39] Banerjee A, Hamberger B. P450s controlling metabolic bifurcations in plant terpene specialized metabolism[J]. Phytochem Review, 2018, 17(1): 81-111.
責任编辑:黄东杰
