牦牛GPRIN3基因的克隆及组织表达分析
2021-08-20柴志欣武志娟王吉坤钟金城信金伟
向 娅,柴志欣,武志娟,王吉坤,钟金城,信金伟
(1.西南民族大学青藏高原动物遗传资源保护与利用四川省重点实验室,四川 成都 610041;2.西南民族大学青藏高原动物遗传资源保护与利用教育部重点实验室,四川 成都 610041;3.省部共建青稞和牦牛种质资源与遗传改良国家重点实验室,西藏 拉萨 850000)
G蛋白的全称是鸟嘌呤核苷酸结合蛋白(GTPBinding protein),G蛋白偶联受体(GPCR)是膜整合蛋白中最多的一类,为细胞表面受体超家族.在动物中,G蛋白主要在细胞通讯过程中发挥作用,传递细胞外信号,如激素、神经递质、离子和脂质,以及感觉刺激,如信息素、口味和增味剂[1-3].涉及G蛋白偶联受体(GPCRs)的信号通路可调节哺乳动物的多种生理过程,是许多药物治疗的主要靶点.G蛋白在结构上的差异主要源于α亚单位,α亚单位的多样性实现了G蛋白对多种生理功能的调节[4-5],根据α亚单位可将G蛋白分为四个亚家族:①Gαs(激活腺苷酸环化酶),产生重要胞内信使cAMP,激活依赖于cAMP的蛋白激酶A,还可激活磷脂酶C和钙通道;②Gαi(抑制腺苷酸环化酶),调控部分离子的转运和磷脂肌醇的代谢,参与促有丝分裂原蛋白激酶级联和Hh信号传导途径,通常把Gt和Go也归入该家族;③Gαq,介导磷脂酶C的激活和多种胞内第二信使的产生及蛋白激酶C的激活;④Gα12/13,与凝血酶受体偶联,参与激活MAPK且不依赖于Rho因子,还可介导Rho因子依赖的传导信号到细胞骨架的过程[6].Gαi/o调节蛋白可诱导G蛋白调节诱导神经突增生家族(G protein-regulated inducer of neurite outgrowth family,GPRIN)介导GPCR与顺序细胞内靶标之间的通讯[7].
GPRIN家族是具有组成型活性形式的共表达信号传导分子,包括GPRIN1、GPRIN2和GPRIN3三个亚型,这三个成员间表现出显著的同源性.GPRIN抗体已广泛应用于神经生物学和信号转导领域的研究,Gαo可以刺激神经分支细胞,但其在体内的作用机制尚不清楚,GPRIN可能是Go调节神经生长的下游作用因子,Gαo-GRIN相互作用可调节神经突增生,使细胞通过GPCR-Gαi/O介导的信号分化为神经突起.研究发现,Gαi/O-GPRIN信号在小鼠神经系统发育中具有重要作用,能够调节神经元的定向迁移,Gαo-GPRIN途径可以显著调控共表达神经元在胚胎阶段的迁移和分化,从出生后到成熟阶段,发挥定位功能并维护特定的神经系统,在神经元中特异表达并定位于树突和轴突且与Gαo相互作用的GPRIN蛋白很可能参与信号转导途径和神经元分化过程[7-9].还有研究表明,GP RIN1介导了μ阿片受体的功能[10],与激活Gαo蛋白的Cdc42结合,导致细胞形态发生显著变化[8].GPRIN2也有类似的现象[9],GPRIN1和GPRIN2被称为GPCR信号的替代(腺苷酸环化酶)介体[7].GPRIN3的作用定义不多,通过BLAST搜索发现它为脊椎动物所特有,在大脑中高表达,最近被标记为β-arrestin-2的伴侣,可调节多巴胺受体脱敏,影响选择性行为任务[11].
GPRIN3的研究主要集中在人类疾病方面,杨婵[12]研究表明,参芪复方可能通过同为差异mRNA和差异lncRNA靶基因的GPRIN3基因调控细胞增殖凋亡,从而抑制炎症反应,防治糖尿病大血管(胸主动脉)病变.Karadurmus[13]等研究发现,GPRIN3是纹状体中D2R功能的假定选择性控制器,在纹状体相关行为和细胞功能中发挥关键作用,这为解决与D2R相关的纹状体功能障碍(精神分裂症,帕金森氏病等)提供了新方向.目前尚未见牦牛GP RIN3基因相关研究报道,故本研究通过PCR技术克隆麦洼牦牛GPRIN3基因CDS区序列,分析其核苷酸、氨基酸序列,预测其蛋白结构和功能,利用RT-qPCR技术分析其组织表达情况,旨在探讨牦牛GP RIN3基因的分子特性.
1 材料与方法
1.1 试验动物
于四川省阿坝藏族羌族自治州红原县选取雄性麦洼牦牛3头(0.5岁).屠宰后快速采集心脏、肝脏、肺脏、肾脏、臀肌等组织,DEPC水冲洗后,置于液氮中保存备用.
1.2 试验试剂
北京天根生化科技有限公司:动物组织DNA提取试剂盒、RNA反转录试剂盒、DH5α感受态细胞、DNA纯化回收试剂盒;TaKaRa公司:2000 bp Marker、pMD19-T Vector cloning kit、TB Green Premix Ex Taq.
1.3 提取基因组DNA
使用动物组织DNA提取试剂盒获得牦牛肺脏组织基因组DNA,loading buffer染色,10 g/L琼脂糖凝胶电泳检测DNA样品质量;NanoDrop 2000分光光度计测定其浓度及OD260nm/280nm值,OD260nm/280nm值在1.8~2.0方可使用,置于-20℃保存备用.
1.4 总RNA提取及cDNA合成
利用Trizol法提取牦牛各组织总RNA.采用NanoDrop 2000分光光度计测定RNA浓度及OD260nm/OD280nm(Ratio,R)值,R在1.8~2.0时方可采用.当R<1.8时,说明溶液有蛋白或其他有机物污染;当R>2.2时,说明RNA已经水解成单核酸.配制1.0%琼脂糖凝胶,5μL RNA样品加1μL 6×Loading buffer,混匀,180 V电泳5 min,于凝胶成像仪上拍摄,检测RNA样品质量,并反转录成cDNA,置于-20℃保存备用.
1.5 基因克隆及荧光定量PCR
根据GeneBank收录的野牦牛GPRIN3基因序列(XM_005897777)及牦牛GAPDH(XM_014482068)基因序列,采用Primer 5.0设计特异性引物(见表1),由成都擎科梓熙生物技术有限公司合成.

表1 GPRIN3基因引物序列Table 1 GPRIN3 gene primer sequences
以牦牛耳样基因组DNA为模板进行PCR扩增.反应体系、程序见表2、表3.PCR产物经10 g/L琼脂糖凝胶电泳检测,紫外灯下切胶,根据DNA纯化回收试剂盒说明书纯化回收PCR产物,4℃存放.将PCR回收产物与载体进行连接、转化、复苏、倒置培养,具体操作参照向娅等[14]基因克隆步骤.荧光定量PCR反应体系、程序见表2、表3,每个样品3个生物学重复,3个技术重复.

表2 PCR反应体系Table 2 PCR reaction system

表3 PCR反应程序Table 3 PCR reaction program
1.6 生物信息学分析
分析麦洼牦牛基因组DNA序列,预测其蛋白结构和功能(表4).

表4 DNA和蛋白质序列数据分析工具Table 4 DNA and protein sequence data analysis tools
2 结果与分析
2.1 PCR扩增结果
以10 g/L琼脂糖凝胶电泳检测提取的牦牛基因组DNA样品,结果显示,条带单一清晰(见图1A),可用于后续实验;用紫外分光光度计检测其浓度并记录,-20℃保存备用.
以麦洼牦牛耳样DNA为模板进行PCR扩增,产物经10 g/L琼脂糖凝胶电泳检测,有目的条带且条带单一,长度为690、717、909、751 bp(见图1B),与预期片段大小一致.
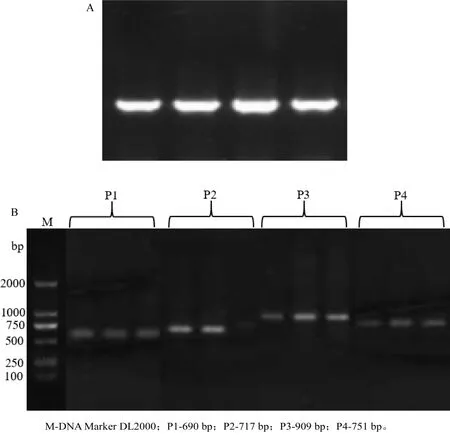
图1 牦牛基因组DNA琼脂糖凝胶电泳(A)和GPRIN3基因PCR扩增结果(B)Fig.1 Agarose gel electrophoresis of genome DNA in yak(A)and PCR amplification results of GPRIN3 gene(B)
2.2 氨基酸序列比对
测序拼接获得麦洼牦牛GPRIN3基因CDS区序列长度为2343 bp,起始密码子为ATG,终止密码子为TGA,编码780个氨基酸残基,序列提交GenBank得到登录号MT978152.将麦洼牦牛与GenBank中野牦牛(XM_005897777)GPRIN3的核苷酸序列比对显示(见图2),共有4处碱基突变,分别位于第309、413、1099、1212位点,一致性为99.83%;氨基酸序列比对显示,共3个位点发生突变,分别是第103(G→R)、366(T→M)、404(E→K)位点,一致性为99.62%;第413位的碱基突变属于同义突变.

图2 牦牛、野牦牛GPRIN3基因氨基酸序列比对Fig.2 Sequence comparison ofdiduced amino acid sequence of GPRIN3 genebetween yak and wild yak
2.3 系统进化树分析
从NCBI数据库中下载8个不同物种GPRIN3基因的CDS区序列,利用MEGA 7.0软件分析牦牛、野牦牛、普通牛等8个物种GP RIN3基因编码区核苷酸序列并构建系统进化树.结果显示,牦牛与野牦牛遗传距离最近,其次是普通牛,再与野猪、犬聚为一类,与小鼠、人亲缘关系较远,与黑猩猩的亲缘关系最远(见图3),说明GPRIN3基因编码区在哺乳动物间高度保守.

图3 GPRIN3基因系统进化树Fig.3 Phylogenetic tree of GPRIN3
2.4 蛋白理化性质预测与分析
蛋白理化性质预测结果显示,牦牛GPRIN3蛋白分子式为C3481H5697N1053O1190S29,分子质量为82.270 ku,等电点为7.18,为偏碱性蛋白,负电荷氨基酸残基(Asp+Glu)、正电荷氨基酸残基(Arg+Lys)均为85个.由氨基酸组成可知(见表5),Ser(S)含量最丰富,占比13.20%,Trp(W)、Tyr(Y)含量最低,均只占0.3%.不稳定指数为60.07,说明牦牛GPRIN3蛋白结构不稳定;脂肪系数为63.12;总平均亲水系数(GRAVY)为-0.644,与ExPASy-ProtScale预测结果相符,牦牛GPRIN3氨基酸序列中大多数氨基酸为亲水性残基,此蛋白属于亲水性蛋白.
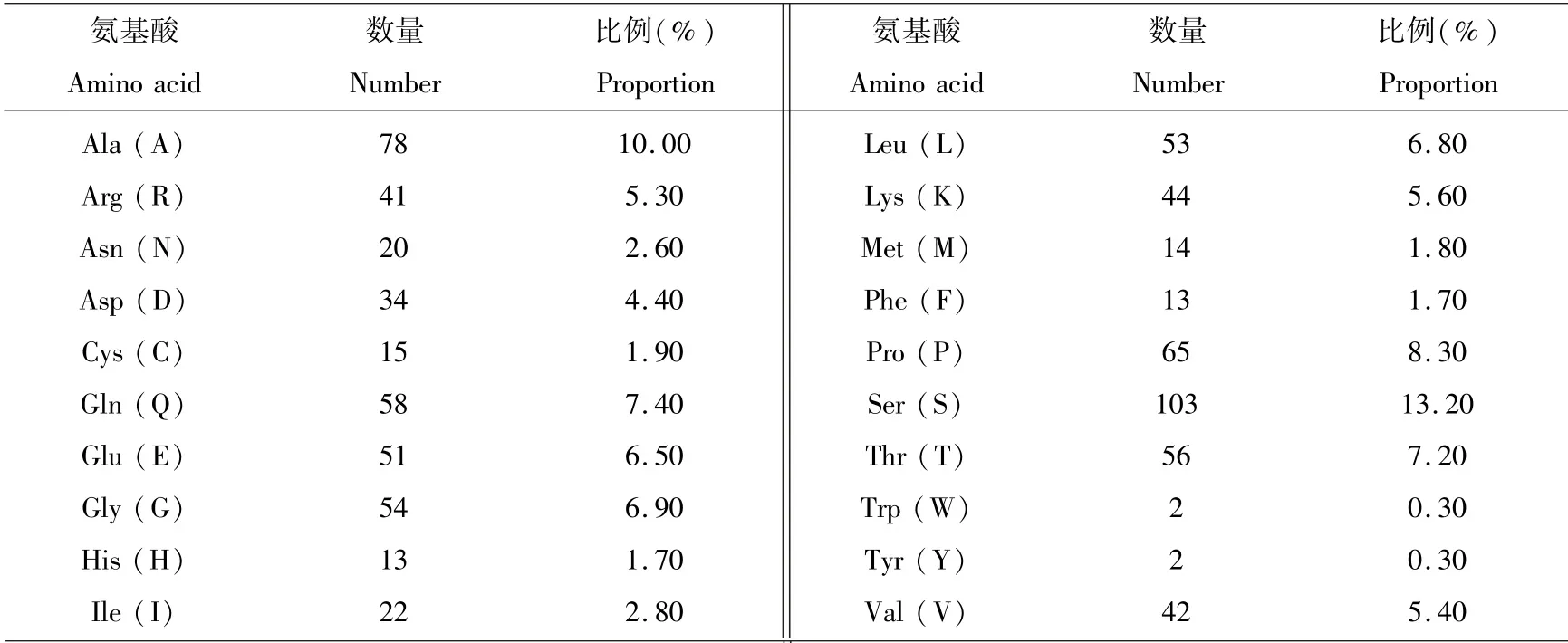
表5 麦洼牦牛GPRIN3基因编码蛋白的氨基酸组成Table 5 The amino acid composition of the protein encoded by the Maiwa yak GPRIN3 gene
2.5 蛋白结构与功能预测
2.5.1 跨膜结构及信号肽预测
跨膜结构预测显示,GPRIN3蛋白的氨基酸均位于细胞膜表面,无跨膜结构域;结合ExPASy-TMPred软件分析结果,牦牛GPRIN3蛋白在第384~404位氨基酸处存在1个由内而外的跨膜螺旋,但仅为明显的TM片段,为非跨膜蛋白(见图4).信号肽预测结果表明,牦牛GPRIN3蛋白不含信号肽,为非分泌型蛋白(见图5).

图4 麦洼牦牛GPRIN3基因编码蛋白跨膜结构域预测Fig.4 Transmembrane domain prediction of Maiwa yak GPRIN3 protein
图5 麦洼牦牛GPRIN3基因编码蛋白信号肽预测Fig.5 Signal peptide prediction of Maiwa yak GPRIN3 gene encoding protein
2.5.2 磷酸化位点及糖基化位点预测
磷酸化位点预测结果显示,阈值为0.5时,牦牛GPRIN3蛋白存在74个Ser磷酸化位点,32个Thr磷酸化位点,无Tyr磷酸化位点(见图6).糖基化位点预测结果表明,GPRIN3蛋白含有152个O-糖基化位点,3个N-糖基化位点(第271、422、652位)(见图7).
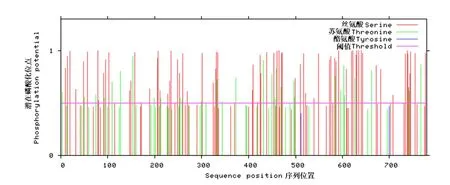
图6 麦洼牦牛GPRIN3基因编码蛋白磷酸化位点预测Fig.6 Phosphorylation sites prediction of Maiwa yak GPRIN3 gene encoding protein

图7 麦洼牦牛GPRIN3基因编码蛋白N-糖基化位点预测Fig.7 N-glycosylation site prediction of Maiwa yak GPRIN3 gene encoding protein
2.5.3 二级结构及三级结构预测
二级结构预测显示,牦牛GPRIN3蛋白无规则卷曲(Cc)含量最高,共562处,占二级结构的72.05%;α螺旋(Hh)有158处,占比20.26%;延伸链(Ee)42处,占二级结构的5.38%;β转角(Tt)含量最低,仅2.31%(见图8).

图8 牦牛GPRIN3蛋白二级结构预测长线为α-螺旋(Hh),中长线为延伸链(Ee),短线为β-转角(Tt),最短线为无规则卷曲(Cc)Fig.8 Secondary structure prediction of yak GPRIN3 proteinThe long line isα-helix(Hh),the middle and long line is the extended chain(Ee),the short line is theβ-turn(Tt),and the shortest line is the random coil(Cc)
基于PDB数据库预测GPRIN3蛋白的三级结构模型(见图9),牦牛GPRIN3蛋白与2g0f.1.A模型相似性最高,三级结构以无规则卷曲为主,与二级结构预测结构相符.
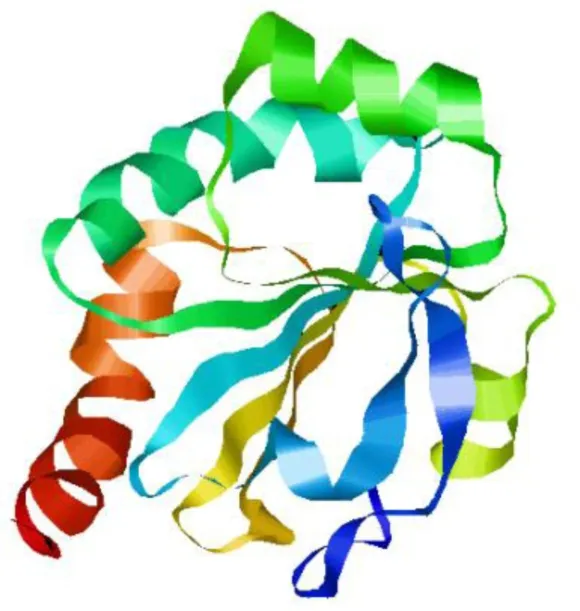
图9 牦牛GPRIN3蛋白三级结构预测Fig.9 Tertiary structure prediction of yak GPRIN3 protein
2.5.4 保守结构域预测
保守结构域预测显示,牦牛GPRIN3蛋白含有GRIN-C超家族结构域,GRIN-C超家族代表G蛋白调节神经突生长诱导物的C末端(见图10).
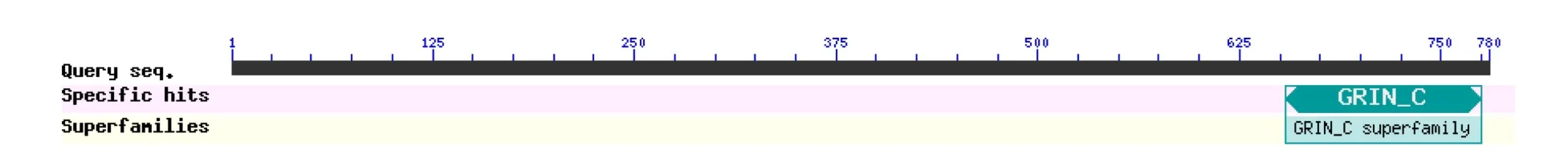
图10 牦牛GPRIN3基因编码蛋白保守结构域预测Fig.10 Conserved domain prediction of yak GPRIN3 gene encoding protein
2.5.5 蛋白功能分析
蛋白亚细胞定位预测显示,牦牛GPRIN3蛋白主要分布于细胞核(82.6%)中,13.0%分布于细胞骨架中,在分泌系统的囊泡中仅占4.3%.蛋白功能分类预测表明,牦牛GPRIN3蛋白可能在辅因子的生物合成、运输和结合、翻译途径中发挥重要作用(见表6),同时在电压激活离子通道过程中发挥功能(见表7).

表6 牦牛GPRIN3基因编码蛋白功能分类分析Table 6 Functional category analysis of yak GPRIN3 gene encoding protein
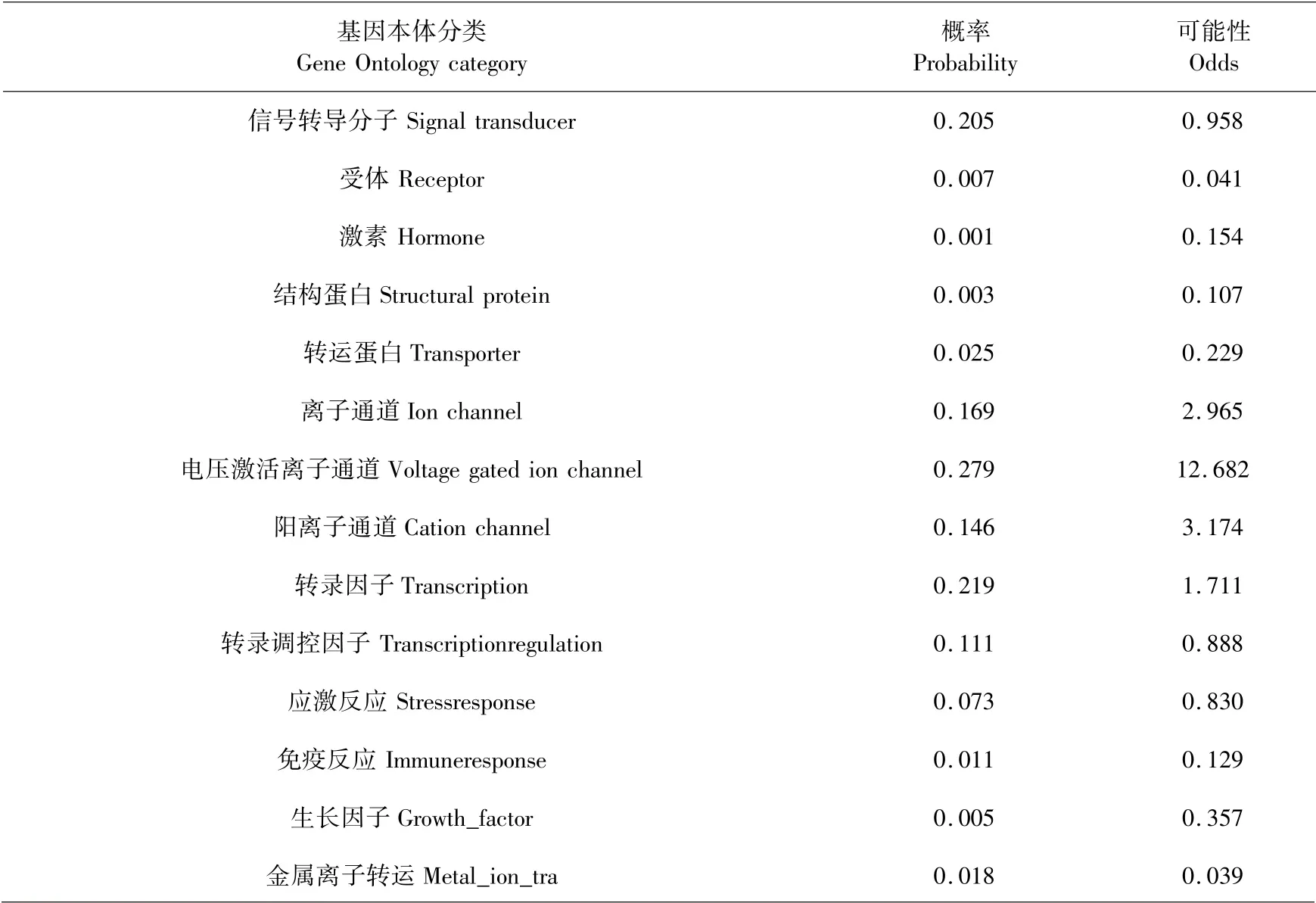
表7 牦牛GPRIN3基因本体分类分析Table 7 Gene Ontology category analysis of yak GPRIN3 gene encoding protein
2.5.6 蛋白互作分析
分析牦牛GPRIN3蛋白的互作关系发现,GPRIN3与WIZ蛋白相关性最高(0.721)(见图11),WIZ是一种蛋白质向导,可将EHMT1和EHMT2组蛋白甲基转移酶链接到CTBP核心加压机制,可参与EHMT1-EHMT2异二聚体的形成和稳定,属于C2H2型锌指蛋白家族.与GPRIN1蛋白相关性为0.688,GPRIN是G蛋白调节神经突生长的诱导物1,与神经突生长有关.SYNGAP1(0.468)是突触RasGTPase活化蛋白1,突触后信号PSD的主要成分,可在AMPAR膜运输和突触可塑性的NMDAR依赖性控制中发挥重要作用,可调节AMPAR介导的微型兴奋性突触后电流.
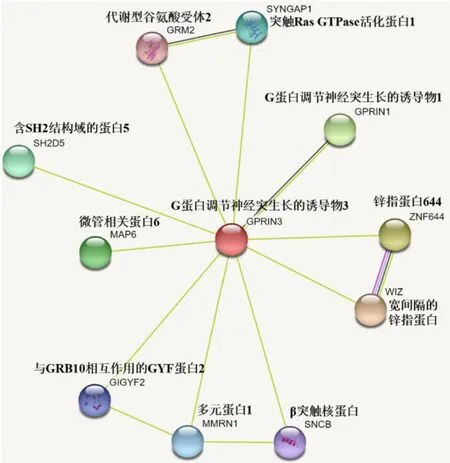
图11 GPRIN3蛋白网络互作分析Fig.11 GPRIN3 protein network interaction analysis
2.6 牦牛GPRIN3 mRNA组织表达分析
2.6.1 标准曲线及线性范围
将PCR原液10倍梯度稀释的6个浓度(10-1~10-6)作为模板,每个浓度3个重复进行实时荧光定量PCR,根据扩增曲线得到Ct值,QuantStudio软件自动拟合出标准曲线(见图12).GP RIN3基因扩增效率为100.826%,相关系数为0.990,表明建立的标准曲线线性关系很好,标准线性方程为:y=-3.302x+1.260.
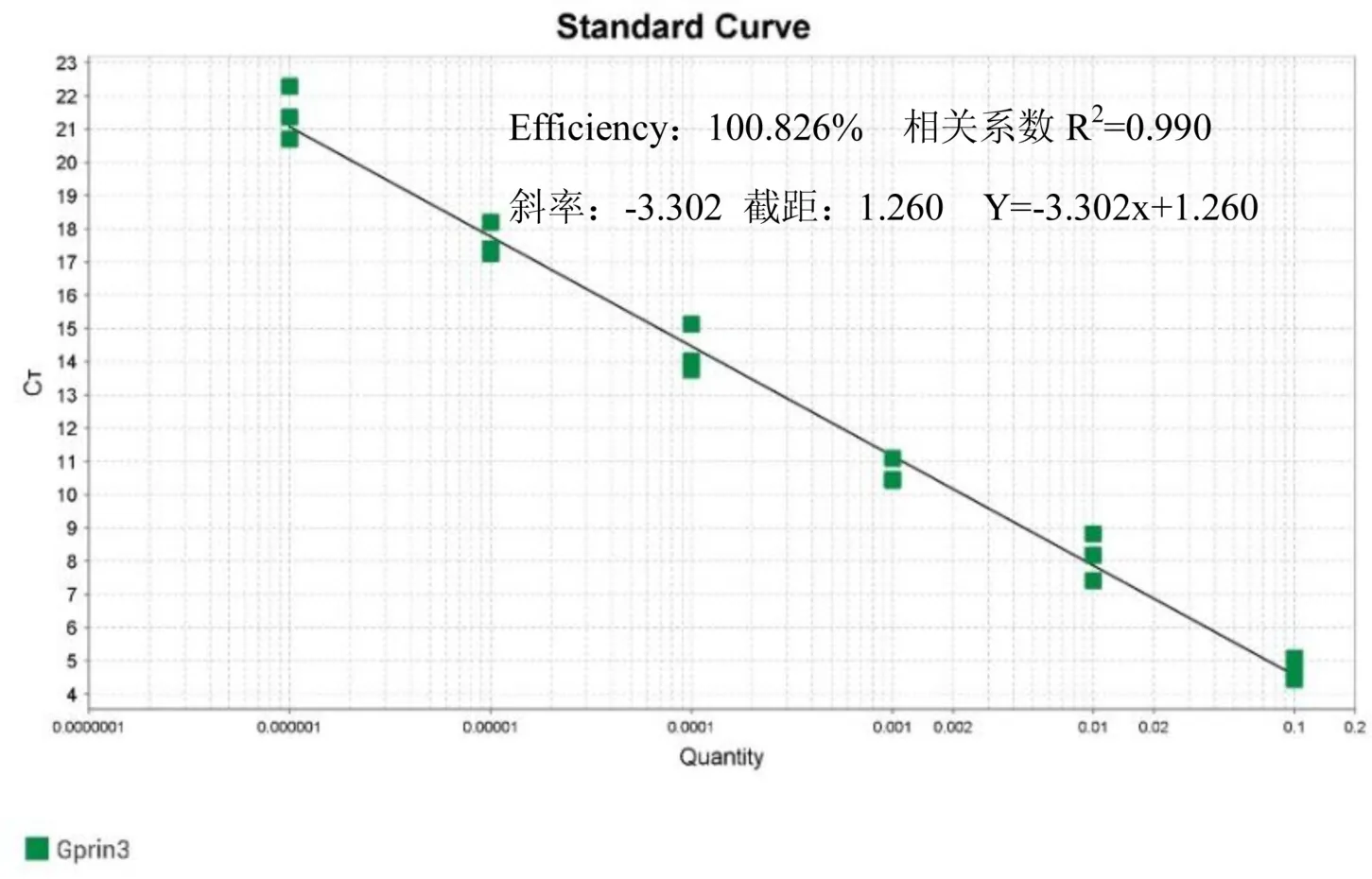
图12 GPRIN3基因的标准曲线Fig.12 Standard curve of GPRIN3 gene
2.6.2 荧光定量PCR结果
荧光定量结果显示,GPRIN3基因在麦洼牦牛心脏、肝脏、肺脏、肾脏和臀肌组织中均有表达,在肺脏组织中表达量最高,与肝脏组织差异显著(P<0.05),与肾脏、心脏组间差异极显著(P<0.01),在臀肌组织中表达量极低,且与其他组织均差异极显著(P<0.01)(见图13).
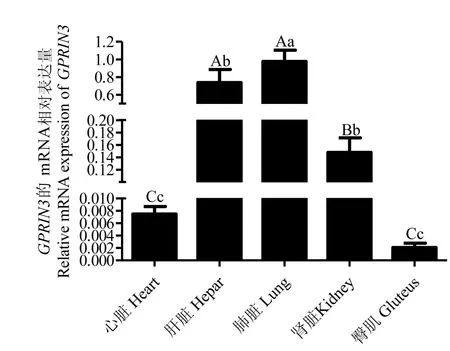
图13 GPRIN3基因在麦洼牦牛不同组织的mRNA表达量大写字母表示差异极显著(P<0.01),小写字母表示差异显著(P<0.05);字母不同表示差异显著,字母相同表示差异不显著(P>0.05)Fig.13 mRNA expression of GPRIN3 gene in different tissues of Maiwa yakCapital letters means the difference is extremely significant(P<0.01),lowercase letters means the difference is significant(P<0.05);different letters means the difference is significant,and the same letters means the difference is insignificant(P>0.05)
3 讨论
大量研究表明,G蛋白主要在细胞通讯过程中发挥作用,传递细胞外信号,而细胞通讯异常可能导致牦牛生长发育障碍甚至死亡,GPRIN3基因可能作为影响牦牛生长发育的候选基因发挥作用.本研究对麦洼牦牛GP RIN3基因进行了克隆、生物信息学分析及组织表达分析研究,初步探索该候选基因在其各组织中的表达情况.
本研究中麦洼牦牛与野牦牛GPRIN3基因的核苷酸、氨基酸序列比对结果显示,其与野牦牛核苷酸序列相比,共有4处碱基突变,序列一致性为99.83%;氨基酸序列比对显示,4处碱基突变1处为同义突变,3处属于错义突变,分别是第103(G→R)、366(T→M)、404(E→K)位点,一致性为99.62%.在中枢神经系统中甘氨酸(Gly)与谷氨酸(Glu)均为激动剂,可让动物因超兴奋死亡;甘氨酸在CNS中广泛分布,在神经信号传递及参与各种生理和病理反应中发挥重要作用[15].因此,推测甘氨酸、谷氨酸的突变可能会影响牦牛神经信号传导进而影响牦牛相关激素的分泌,从而影响牦牛生长发育.系统进化树结果显示,牦牛与野牦牛遗传距离最近,其次是普通牛,再与野猪、犬聚为一类,与黑猩猩的亲缘关系最远,说明GPRIN3基因编码区在哺乳动物间高度保守.
本研究牦牛GPRIN3蛋白中丝氨酸(13.2%)、丙氨酸(10.0%)、脯氨酸(8.0%)含量较高.丝氨酸(Ser)在脂肪和脂肪酸的新陈代谢、肌肉生长、细胞膜的制造加工、肌肉组织和包围神经细胞的鞘的合成中均发挥重要作用[16];D-丝氨酸同时也是重要的神经递质,可调控中枢系统[17].谭茹尹等[18]研究发现,包括人类在内,高等动物的高级中枢存在区域性高浓度的D-丝氨酸,并发挥着重要的神经递质作用,与癫痫、精神分裂症、焦虑症、成瘾等神经精神疾病的发生发展有密切关系.脯氨酸(Pro)是动物胶原蛋白的重要组成成分,进入肽链后可发生羟基化作用.根据伊利诺斯大学一项最新的研究,脯氨酸重复对于理解许多天然蛋白的结构和功能很重要;且脯氨酸重复片段越长,泡沫状的蛋白就越大,研究人员推测这些发现有助于神经组织退化疾病的治疗[19].Karadurmus[13]等研究表明,G PRIN3在间接中棘神经元(indirect medium spiny neurons,iMSN)中通过调控细胞树突状排列,维持兴奋性和D2R依赖性行为.GPRIN3产生的D2R信号微调为治疗需要补充多巴胺的疾病(例如帕金森氏病)或必须减少多巴胺的紧张性疾病(例如精神分裂症)提供了新思路,但其具体机制尚未明确.故后续的研究可以此为思路,探索GPRIN3产生D2R信号微调对病例情况的影响,为预防牦牛神经性疾病提供参考.
亚细胞定位预测显示,GPRIN3蛋白主要分布于细胞核、细胞骨架和分泌系统的囊泡中,且在细胞核中分布最多,可发挥核定位作用.保守结构域预测显示,GPRIN3蛋白含有GRIN-C超家族结构域.对牦牛GPRIN3蛋白进行功能分类预测,结果表明,GPRIN3蛋白可能在辅因子的生物合成、运输和结合、翻译途径中发挥重要作用,同时在电压激活离子通道过程中发挥功能.结合保守结构域预测结果,GPRIN蛋白可能在调节辅因子的生物合成、运输和结合及细胞通讯等过程发挥重要作用.
麦洼牦牛主要生活在高海拔、低压缺氧的环境,为适应高原极端恶劣生态环境,肺脏、肝脏等机体主要呼吸器官发生显著变化,实时荧光定量PCR结果表明,GP RIN3在麦洼牦牛肺脏组织中表达量最高,与肝脏组织差异显著(P<0.05),与肾脏、心脏组间差异极显著(P<0.01),在臀肌组织中表达量极低.目前,GPR IN3基因在家畜中的研究较少缺乏可供参考比对的数据和文献,通过本研究对牦牛GPRIN3基因的克隆和组织表达分析可为该基因在家畜中的研究提供可参考的数据.
4 结论
本研究克隆获得麦洼牦牛GP RIN3基因长度为2343 bp的开放阅读框,编码780个氨基酸残基;系统进化树结果显示,牦牛与野牦牛、普通牛亲缘关系最近;存在GRIN-C保守功能结构域,是G蛋白调节诱导的神经突增生C末端结构域,推测GPRIN3可能调节辅因子的生物合成、运输和结合,并在细胞通讯等过程中发挥重要作用;实时荧光定量PCR结果表明,G PRIN3在牦牛肺脏组织中表达量最高,本研究为今后进一步探讨牦牛G PRIN3基因及其编码蛋白的结构和功能提供理论基础.
