Alternative models of cancer stem cells: The stemness phenotype model, 10 years later
2021-07-30VivekKaushikYogeshKulkarniKumarFelixNeelamAzadAnandKrishnanIyerJuanSebastianYakisich
Vivek Kaushik, Yogesh Kulkarni, Kumar Felix, Neelam Azad, Anand Krishnan V Iyer, Juan Sebastian Yakisich
Vivek Kaushik, Yogesh Kulkarni, Kumar Felix, Neelam Azad, Anand Krishnan V Iyer, Juan Sebastian Yakisich, School of Pharmacy, Department of Pharmaceutical Sciences, Hampton University, Hampton, VA 23668, United States
Abstract The classical cancer stem cell (CSCs) theory proposed the existence of a rare but constant subpopulation of CSCs. In this model cancer cells are organized hierarchically and are responsible for tumor resistance and tumor relapse. Thus,eliminating CSCs will eventually lead to cure of cancer. This simplistic model has been challenged by experimental data. In 2010 we proposed a novel and controversial alternative model of CSC biology (the Stemness Phenotype Model, SPM).The SPM proposed a non-hierarchical model of cancer biology in which there is no specific subpopulation of CSCs in tumors. Instead, cancer cells are highly plastic in term of stemness and CSCs and non-CSCs can interconvert into each other depending on the microenvironment. This model predicts the existence of cancer cells ranging from a pure CSC phenotype to pure non-CSC phenotype and that survival of a single cell can originate a new tumor. During the past 10 years, a plethora of experimental evidence in a variety of cancer types has shown that cancer cells are indeed extremely plastic and able to interconvert into cells with different stemness phenotype. In this review we will (1) briefly describe the cumulative evidence from our laboratory and others supporting the SPM; (2) the implications of the SPM in translational oncology; and (3) discuss potential strategies to develop more effective therapeutic regimens for cancer treatment.
Key Words: Cancer; Stem cells; Stemness; Plasticity; Chemotherapy; Interconversion
INTRODUCTION
The biological properties of cancer cells have profound implications for all areas of oncology research ranging from preclinical studies to advanced clinical trials. It is not a surprise that numerous conceptual models of cancer cell biology have been proposed with the ultimate goal to develop effective therapies that not only extend survival but lead to a definitive cure. In the past decades the discovery of the potential abilities in self-renewal and differentiation of normal stem cells has opened a new horizon in medicine and important concepts were extrapolated to neoplastic cells. Although the concept of cancer stem cell (CSC) is not new since the general idea of tumors driven by a subset of cells endowed with stem-like properties was postulated by Rudolf Virchow in 1855[1] it has gained tremendous momentum after the isolation of putative cancer stem-like cells (CS-LCs) in a variety of cancer types including brain tumors[2-5], breast cancer[6-9], colon[10], hepatic[11], pancreatic[12], thyroid[13,14], bladder[15,16],cervical[17-20], ovarian[21-24], urothelial[25-28], renal[29-31] and chordoma[32,33]. In general, putative CS-LCs were isolated from every type of fresh tumor specimens and cancer cell lines. These discoveries quickly led to a new paradigm, the so called“Cancer Stem Cell Theory” (CSCT), that is fundamentally not completely different from the original models proposed by Virchow and his contemporaries[1]. In this model there is a hierarchical organization where a subset of CSCs can irreversibly differentiate into all types of non-CSCs. Prior to the modern CSCT, that started with the first isolation of putative CS-LCs in 1997[34] the clonal stochastic model (cSM)postulated in 1976[35] was popular among oncologists. The cSM proposed that all transformed cells in the tumor have carcinogenic potential and are able to proliferate and produce the same cells. The cSM is a non-hierachical model. From the clinical point of view, according to the cSM, to cure cancer all cancer cells should be eliminated since any cancer cell is potentially tumorigenic. On the contrary, according to the modern CSCT, to cure cancer or at least to obtain a significant outcome, it should be enough to eliminate only the rare subpopulation of CSCs. The idea of a rare subpopulation of CSCs as driving element in cancer development, evolution and heterogeneity, has overridden the previous cSM model[7] and catapulted research of therapeutic strategies based on CSCs targeting, such as the targeting of CSC niche,CSC signaling pathways, and CSC mitochondria and, metabolism[7,36-38].
Although the modern CSCT was an attractive concept it was found soon to be insufficient to reconcile experimental findings with a hierarchical rigid model. As a consequence several alternative plasticity models of CSCs such as “Stemness Phenotype Model”[39], the “complex system model”[40], the “Dynamic CSC model”[41], and the “Dedifferentiation model”[42] were proposed as early as 2010. These models share some similarities, but a detailed comparison is beyond the scope of this article. For review see Cruzet al[43]. The aim of this short review is to update and highlight key predictions of the Stemness Phenotype Model (SPM).
THE SPM
In short, the SPM was originally proposed as a “One compartment model” where there is only one cancer cell type. These cells are cells with different stemness phenotype due to random biological variation. The stemness depends on the microenvironment where the cells grow and can range from a phenotype resembling a non-CSC to a pure CSC. In other words, the SPM proposed that there are no true different subpopulations of CSCs and non-CSCs but a single cell type that can interconvert into each other depending on the microenvironmental conditions. An immediate prediction of this model is that there are cells having “intermediate phenotypes”between both extreme phenotypes[39]. Other key prediction of the SPM is that the survival of a single cell might induce tumor relapse and therefore any effective therapy will must be able to eliminate 100% of cancer cells at once in order to prevent regrowth[43].
MICROENVIRONMENTALLY-DRIVEN INTERCONVERSION BETWEEN CSCS AND NON-CSCS
In the SPM microenvironmentally-driven interconversion between CSCs and non-CSCs is a key process that explains the characteristic found in tumors such as the existence of intratumoral heterogeneity and chemoresistance. Evidence of intratumoral heterogeneity due to interconversion between cancer cell phenotypes was likely observed long before the first isolation of putative CSCs. For instance, in 1987 it was reported that several different cell phenotypes coexist in the human breast cancer cell line MCF7[44]. Similarly, it was known by 1998 that the human lung carcinoma cell line DLKP contains 3 distinct subpopulations and that two of them can interconvert to the third one[45]. At present, the findings of these two examples can easily be explained by interconversion but at that time the concept of stemness was not common in the literature. Definitive evidence of microenvironmentally-driven interconversion between CSCs and non-CSCs phenotypes were already available after the isolation of putative CSCs (characterized by stemness markers) when it was found that cells that were considered non-CSCs could interconvert into CSCs. For instance, (1) some CD44−Du145 prostate cancer cells (100% purity) could give rise to CD44+cells in culture[46];and (2) non-SP MCF7 breast cancer cells when recultured after being sorted contained SP cells indicating that the non-SP fraction gave rise to a new SP subpopulation[47]. A direct conversion from a non-CSC phenotype to a CSC phenotype was demonstrated in breast and prostate cancer cells when it was observed that exposure to conditioned media stimulated non-CSCs to become CSCs and that IL6 was enough to drive this conversion[48]. Other examples include the ability of some mature leukemia cells to de-differentiate and reacquire clonogenic and leukemogenic properties[49] and the dedifferentiation of glioma cells to glioma stem-like cells by therapeutic stress[50].Additional evidence of non-CSCs conversion into CSCs were found in osteosarcoma[51], lung[52], pancreatic[53], colon[54] and breast cancers[55].In vitrodata from our lab and others demonstrated that phenotypic changes due to changes in culture conditions are rapid and reversible[56,57]. For instance, cancer cells can become(within three days) highly resistant to conventional anticancer drugs when switched from anchorage-dependent (adherent cells) culture conditions into anchorageindependent (floating cells) culture conditions. Chemosensitivity was quickly restored(within three days) when floating cells were cultured back as adherent cells. Under these conditions, a reversible change in the expression of proteins from multiple pathways was observed demonstrating complex and quick phenotypic adaptations to changing environment[56,57].
THE EXISTENCE OF MULTIPLE SUBPOPULATIONS OF CANCER CELLS
The SPM predicts the existence of multiple subpopulations of cancer cells ranging from a “pure non-CSC phenotype” to a “pure CSC phenotype”. This prediction was confirmed in the non-small cell lung adenocarcinoma (NSCLA) cell lines A549 and H441. It was found that NSCLA cells contain multiple, interconvertible, phenotypically distinct subpopulations (e.g., non-SP, SP, CD133posand ALDHhigh) that exhibit distinct self-renewal and metastatic gene expression patterns[52].
These findings clearly demonstrated that cancer cells are actually extremely plastic and that microenvironmental conditions can influence and drive the bidirectional interconversion between CSCs and non-CSCs phenotypes. Recently several theoretical multi-phenotypic models that include, interconversion and cellular plasticity has been useful in predicting and validating this new paradigm[58-60].
THE ABILITY OF A SINGLE CANCER CELL TO REPOPULATE A TUMOR
The ability of any given cancer cell regardless of its phenotype to reconstitutein vivothe complex intratumoral heterogeneity of any cancer is the ultimate prediction of the SPM.In vivoevidence suggesting that any cancer cell is potentially tumorigenic were available long before any alternative model of CSCs were published. Perhaps the most convincing data was published in 2007 demonstrating that each of the 67 single C6 glioma (Including CD133-) cells plated per miniwell was able to generate a clone and subclones, which subsequently gave rise to a xenograft glioma in the BALB/C-nude mouse[61]. Recently, it has been reported that all 16 subpopulations of highly heterogeneous GBM cultures carry stem cell propertiesin vitro. These cells undergo stochastic state transitions, they showed reversible phenotypic adaptationin vivoand they all formed tumors. More importantly, the authors showed that the phenotypic heterogeneity could also be recreated by single cells of different phenotypic profiles[62].
CLINICAL IMPLICATIONS
Mathematical models of cancer biology are providing insight of strategies for cancer elimination. Simple mathematical models considering two populations of cells: CSCs,which can divide indefinitely, and differentiated cancer cells, which do not divide and have a limited lifespan predict that neither inhibition of CSCs proliferation alone nor stimulation of CSCs differentiation is sufficient for cancer cure[63].
Mathematic modelling ofin vitrogrowth of heterogeneous cell cultures in the presence of interconversion from differentiated cancer cells to CSCs also demonstrated that by targeting only the CSCs subpopulation will not be enough to eradicate cancer and that the chemotherapeutic elimination ofin vitrocultures of heterogeneous cancer cells will be effective only if it targets all cancer cell types[64]. From the clinical point of view, the SPM seems to bring back the field of cancer treatment research to the early days of the cSM. The overall clinical implications of both the SPM and the cSM are essentially the same: they both predict that to cure cancer all cancer cells should be eliminated. However, these two models are conceptually very different and, it can be predicted that to achieve complete elimination of all cancer cells (if we ever achieve that goal) it will require a different approach. It is likely then that a successful chemotherapy regime will require numerous anticancer therapies, each of them targeting a “spectrum” of cancer cell subpopulations that in turn can create serious toxicity issues. The next big challenge in the oncotherapy field will be to develop a safe(low or non-toxic) therapeutic regime that can be administered simultaneously to deplete all cancer cells at once.
REDUCING SYSTEMIC TOXICITY BY SEQUENTIAL CHEMOTHERAPY
In complex, highly heterogeneous tumors the eradication of all cancer cells at once will likely require high doses of anticancer agents +/- radiation/immunotherapy that will severely limit its practical application due to toxicity issues. One alternative to circumvent this problem is to administer them sequentially. Sequential cancer treatment with chemotherapy followed by radiotherapy + high dose chemotherapy followed by autologous peripheral blood stem cell transplantation (APBSCT) has been employed with relatively good outcomes in several cancers such as mantle cell lymphoma[65] and relapsed/refractory acute myeloid leukemia[66]. Sequential multimodalities regimes are being increasingly utilized to treat patients carrying different types of cancers such as gastric cancer[67], pancreatic cancer[68], leukemia[69], non-small cell lung cancer[70] and, breast cancer[71]. Sequential anthracyclineand taxane-based neoadjuvant chemotherapy represents the standard therapeutic approach for the majority of patients with early-stage triple negative breast cancer[72].Novel sequential treatment are also currently investigated at the preclinical level[73].We have demonstrated that a first step treatment with Hydroxyurea left few DBTRG.05MG glioma cells arrested in a senescent-like state. In the second step,salinomycin at low concentration eliminated 100% of these senescent-like cells[74].These cells can grow in suspension as neurospheres, in which the Hedgehog pathway is activated[75]. Thisin vitroexample of sequential chemotherapy suggests that it would be possible to eliminate all cancer cells at once with lower, and therefore less,toxic concentrations.
In the cSM, tumor heterogeneity appears as consequence of random genetic changes concomitant with clonal selection. At some point during its development a tumor may have different genetically defined subpopulations growing in specific microenvironments. Due to the irreversible nature of genetic mutations, specific subpopulations can be permanently eliminated with specific anticancer treatments. For instance,sequential chemotherapy steps with different anticancer drugs can potentially eliminate one or few subpopulations per step which will eventually lead to a cure when the last subpopulation is eliminated (Figure 1). It is important to point out that,due to the high genetic instability of cancer cells, any time gap between steps increases the chances of generating new genetic clones (new cancer cell subpopulations) and thus increases the chances of tumor relapse. According to the SPM any time gap between steps increases the chances of regenerating the cancer cell subpopulation(s)eliminated in previous steps by interconversion. We have demonstratedin vitrothat cancer cells are extremely plastic and they can undergo cycles of phenotypic changes within few days[57]. According to the SPM, multistep treatment regimes may only work if there is no time gap or just gap between steps or if the interconversion process is inhibited (Figure 2).
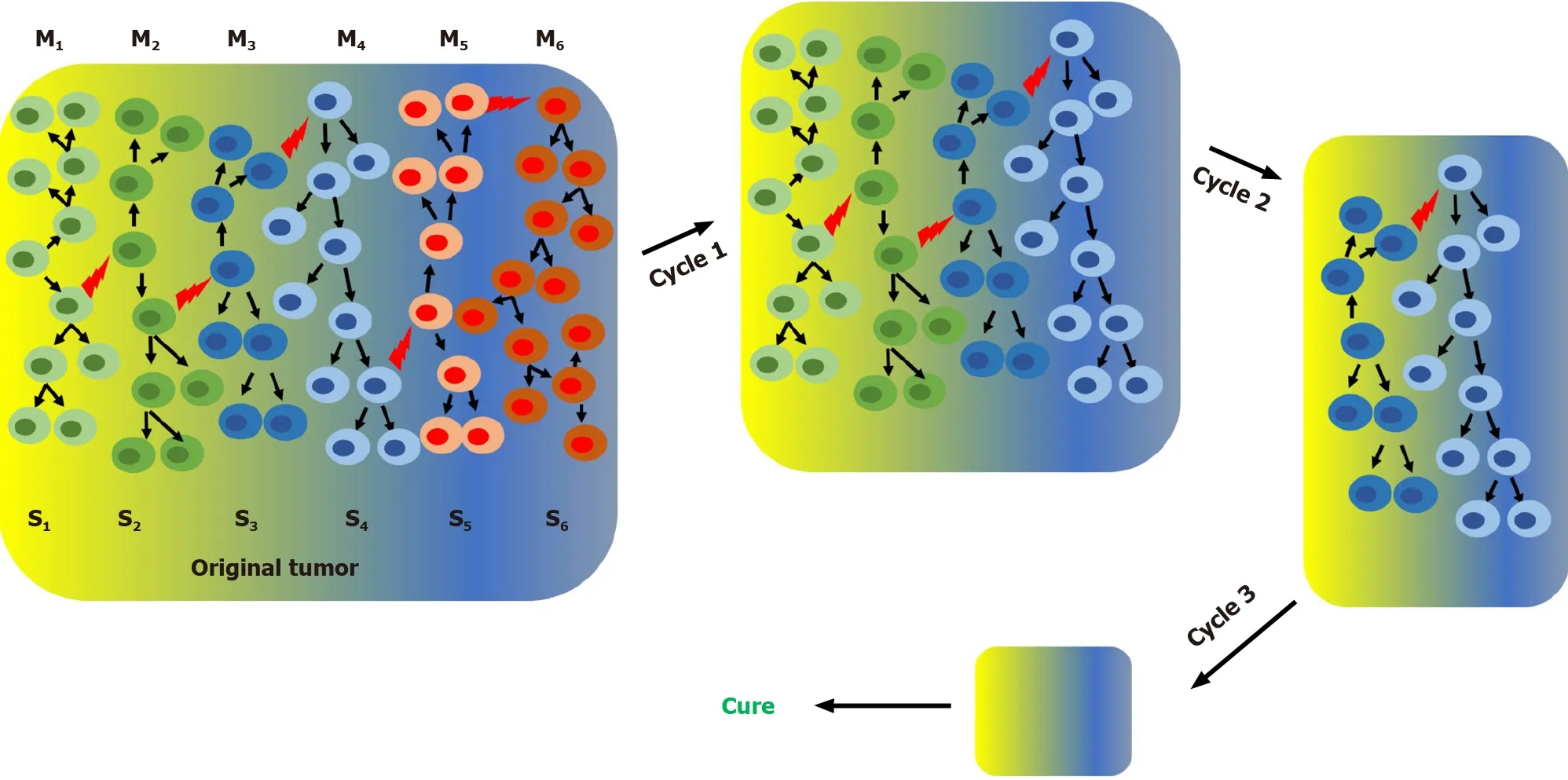
Figure 1 Clonal stochastic model and effect of treatment with sequential chemotherapy cycles (or steps) on tumor progression. According to the clonal stochastic model a complex heterogeneous tumor may contain at a certain time, different subpopulations growing in different microenviroments. Clonal proliferation of single primordial cancer cell originates the original subpopulation (S1). During tumor grow random mutations (represented by red lightning bolts)originates all different subpopulations (S1-S6). M1-M6 represent different microenvironments. Since this model is unidirectional repeated cycles of chemotherapy with no gap can gradually deplete specific cell subpopulations, reducing the tumor size and eventually, if all cancer cells are eliminated, lead to a cure.
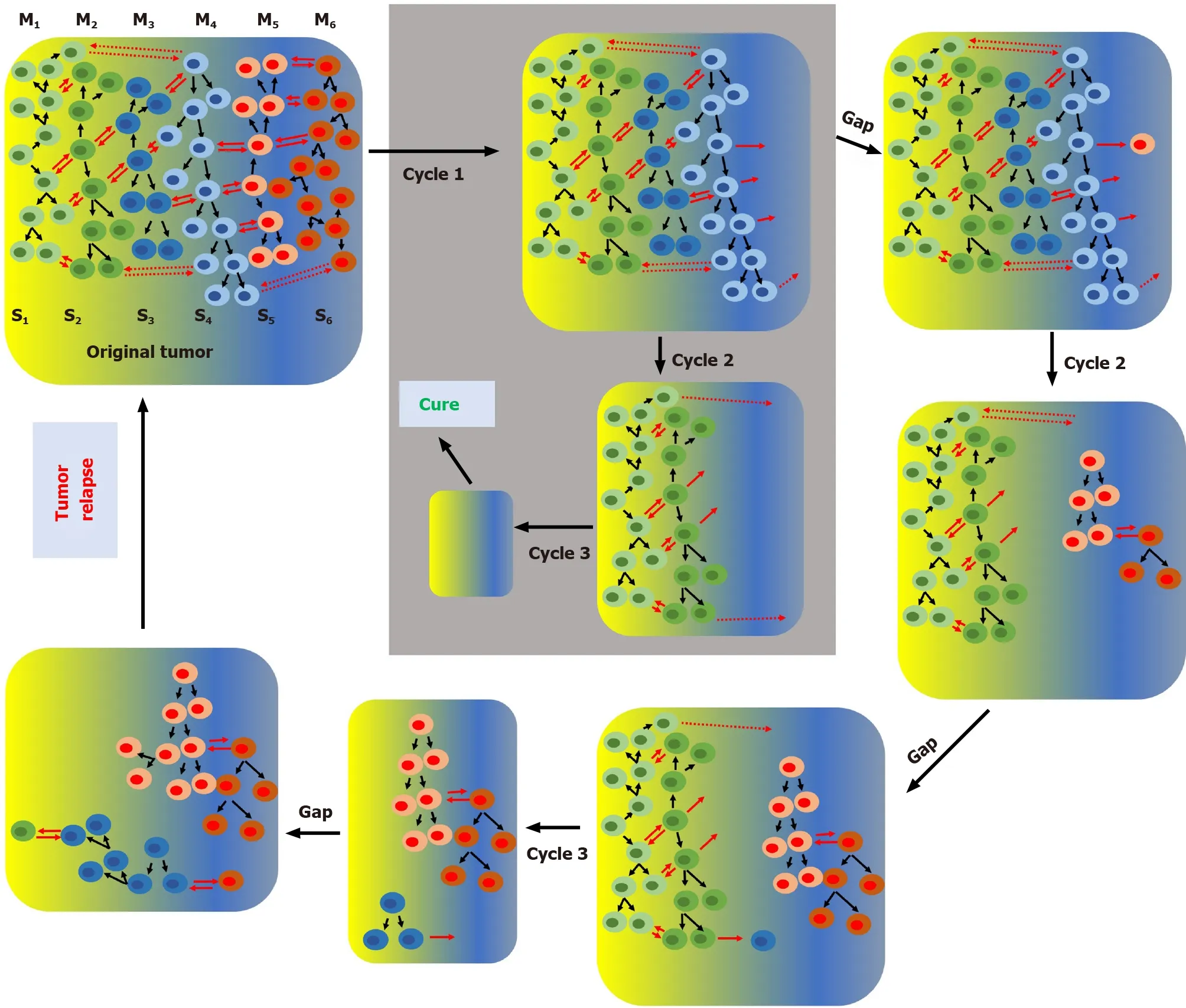
Figure 2 The stemness phenotype model and effect of treatment with sequential chemotherapy cycles (or steps) on tumor progression.According to the stemness phenotype model a complex heterogeneous tumor may contain at a certain time, different subpopulations growing in different microenvironments. Clonal proliferation of a single primordial cancer cell originates the original subpopulation (S1). During tumor growth the original subpopulation can interconvert (represented by red arrows) in other phenotypes originating all different subpopulations (S1-S6). M1-M6 represent different microenvironments. Since this model is bidirectional repeated cycles of chemotherapy can gradually deplete specific cell subpopulations, reducing the tumor size and eventually, if all cancer cells are eliminated, lead to a cure. This scenario (shown inside the grey box) can only occur if interconversion is prevented (e.g., when there is no gap between chemotherapy cycles). Any gap between cycles would allow by interconversion the regeneration of cells sensitive to the previous cycle and eventually, lead to tumor relapse.
THE SPM AND BEYOND
The SPM does not exclude the coexistence of other mechanisms contributing to intratumoral heterogeneity. For instance, in addition to microenvironmentally-driven interconversion between CSCs and non-CSCs, genetic mutations due to genomic instability of cancer cells may create new subpopulations of CSCs in the same tumor[43]. Key concepts from other models not only expand our knowledge of cancer biology but can be useful in designing a curative treatment. For instance, in addition to blocking interconversion, it could be helpful to prevent the generation of new clones originated by stochastic genetic mutations. One of the hallmark of cancer cells is“avoidance of apoptosis” following, for instance, DNA damage[76]. In this context,drugs that promote apoptosis induced by DNA-damage such as PARP alone or in combination with ATR inhibitors[77,78], can potentially reduce the generation of new genetic clones.
CONCLUSION
During the last decade the SPM and similar alternative models of cancer biology have expanded our understanding of cancer biology and new therapeutic targets and biological processes have been identified. For instance, targeting key factors involved in the process of interconversion opens the opportunity to block the conversion of a non-CSCs phenotype into a CSC phenotype and thus reducing chemoresistance and tumor relapse. The SPM, initially proposed for gliomas in a conceptual review article in 2010[39] was quickly extrapolated to other types of tumors[43]. During the last ten years, extensive experimental evidence was published supporting the notion that microenvironmentally-driven interconversion between CSCs and non-CSCs is a key process leading to intratumoral heterogeneity, that in turn is responsible for chemoresistance and tumor relapse. Additionally, key predictions of the SPM – the ability of any given cancer cell to reconstitutein vivothe complex intratumoral heterogeneity has been demonstrated experimentally in gliomas. The SPM has been demonstrated to be a useful working model of cancer biology that should be taken in consideration when developing new cancer treatment modalities. In addition, the SPM is a model of cancer biology (at the cellular level) that does not necessarily exclude key concepts from other cancer models and therefore has the potential to integrate them into more complex tumoral (at the tissue level) models that can be experimentally tested for developing novel treatments.
杂志排行

World Journal of Stem Cells的其它文章
- Epigenetic modulators for brain cancer stem cells: Implications for anticancer treatment
- Mechanisms involved in selecting and maintaining neuroblastoma cancer stem cell populations, and perspectives for therapeutic targeting
- Roles of mitochondrial unfolded protein response in mammalian stem cells
- Stem cell therapies in tendon-bone healing
- Exosomal microRNAs from mesenchymal stem/stromal cells:Biology and applications in neuroprotection
- Immunotherapy against programmed death-1/programmed death ligand 1 in hepatocellular carcinoma: Importance of molecular variations, cellular heterogeneity, and cancer stem cells