Effects of shear stress on differentiation of stem cells into endothelial cells
2021-07-30YanHuangJiaYiQianHongChengXiaoMingLi
Yan Huang, Jia-Yi Qian, Hong Cheng, Xiao-Ming Li
Yan Huang, Jia-Yi Qian, Hong Cheng, Xiao-Ming Li, Key Laboratory for Biomechanics and Mechanobiology of Ministry of Education, Beijing Advanced Innovation Center for Biomedical Engineering, School of Biological Science and Medical Engineering, Beihang University,Beijing 100083, China
Abstract Stem cell transplantation is an appealing potential therapy for vascular diseases and an indispensable key step in vascular tissue engineering. Substantial effort has been made to differentiate stem cells toward vascular cell phenotypes,including endothelial cells (ECs) and smooth muscle cells. The microenvironment of vascular cells not only contains biochemical factors that influence differentiation but also exerts hemodynamic forces, such as shear stress and cyclic strain.More recently, studies have shown that shear stress can influence the differentiation of stem cells toward ECs. A deep understanding of the responses and underlying mechanisms involved in this process is essential for clinical translation. This review highlights current data supporting the role of shear stress in stem cell differentiation into ECs. Potential mechanisms and signaling cascades for transducing shear stress into a biological signal are proposed. Further study of stem cell responses to shear stress will be necessary to apply stem cells for pharmacological applications and cardiovascular implants in the realm of regenerative medicine.
Key Words: Shear stress; Stem cells; Cell differentiation; Endothelial cells; Mechanotransduction
INTRODUCTION
Cardiovascular disease continues to be the leading cause of death globally, with an increase every year[1]. Cell-based therapies have the potential to provide new solutions for treating vascular diseases[2]. Endothelial cells (ECs) derived from donors or differentiated from stem cells are required for various clinical applications, such as promoting angiogenesis in ischemic areas or re-endothelialization of tissue-engineered grafts. Patient-derived primary ECs are limited in number, have donor variabilities,and theirin vitrophenotypes and functions can deteriorate over time[3]. This necessitates the exploration of alternative EC sources. Many methods have been tried to differentiate various stem cells toward ECs[4], such as exogenous growth factors,co-cultivation, and gene transfer. Exogenous growth factors mainly include vascular endothelial growth factor (VEGF), basic fibroblast growth factor (bFGF), epidermal growth factor (EGF), insulin-like growth factor, transforming growth factor and platelet-derived growth factor (PDGF)[5]. In recent years, many studies have shown that in addition to chemical factors, physical stimulation is an important way to regulate the differentiation of stem cells into ECs[6].
The biomechanical patterns of blood flow in vessels are complex[7]. Vascular cells are exposed to hemodynamic forcesin vivo, including flow shear stress and cyclic stretch caused by blood flow[8]. ECs are in direct contact with blood and particularly sensitive to changes in shear stress. Shear stress can affect the morphology, orientation,metabolic activities, and homeostasis of ECs by affecting receptor regulation and signal transmission in ECs (phosphoinositide, and Ca2+and K+ion channels,etc.).Shear stress is a key regulator of EC function and maintenance of vascular homeostasis. Given the crucial role of shear stress in the differentiation of ECsin vivo, it is generally accepted that replicating shear stress in cell culture could be crucial for differentiating stem cells toward the endothelial phenotype[9]. More recently, studies have shown that shear stress can influence the differentiation of stem cells toward ECs.A deep understanding of the responses and the underlying mechanisms involved in this process is essential for clinical translation.
In this review, we present an overview of the role of shear stress in EC oriented differentiation of several types of stem cells, with special reference to experiments conducted using mesenchymal stem cells (MSCs), endothelial progenitor cells (EPCs),embryonic stem cells (ESCs), and induced pluripotent stem cells (iPSCs). We also elucidate potential mechanisms and signaling cascades by which stem cells sense shear stress and transduce the effects of mechanical signals into biological signals. We address various aspects associated with stem cell differentiationviafluid shear stress in the hope of bringing the prospect of regenerative medicine a step closer from bench to bedside. Various receptors and signaling pathways could be pharmacological targets for which novel drugs could be developed in cardiovascular regenerative medicine.
SHEAR STRESS PROMOTES ENDOTHELIAL DIFFERENTIATION OF STEM CELLS
MSCs
MSCs are among the most promising and suitable stem cell types for vascular tissue engineering. MSCs, also known as mesenchymal stromal cells, were first identified in the bone marrow stroma but are also suggested to be present in other tissues. Chenet al[10] proposed that shear stress was able to promote nuclear localization and upregulate expression of β-catenin related to cardiovascular development, EC protection and angiogenesis in MSCs. Many studies have demonstrated that different types of MSCs are able to differentiate into vascular ECs when they are stimulated with shear stress[2] (Table 1).
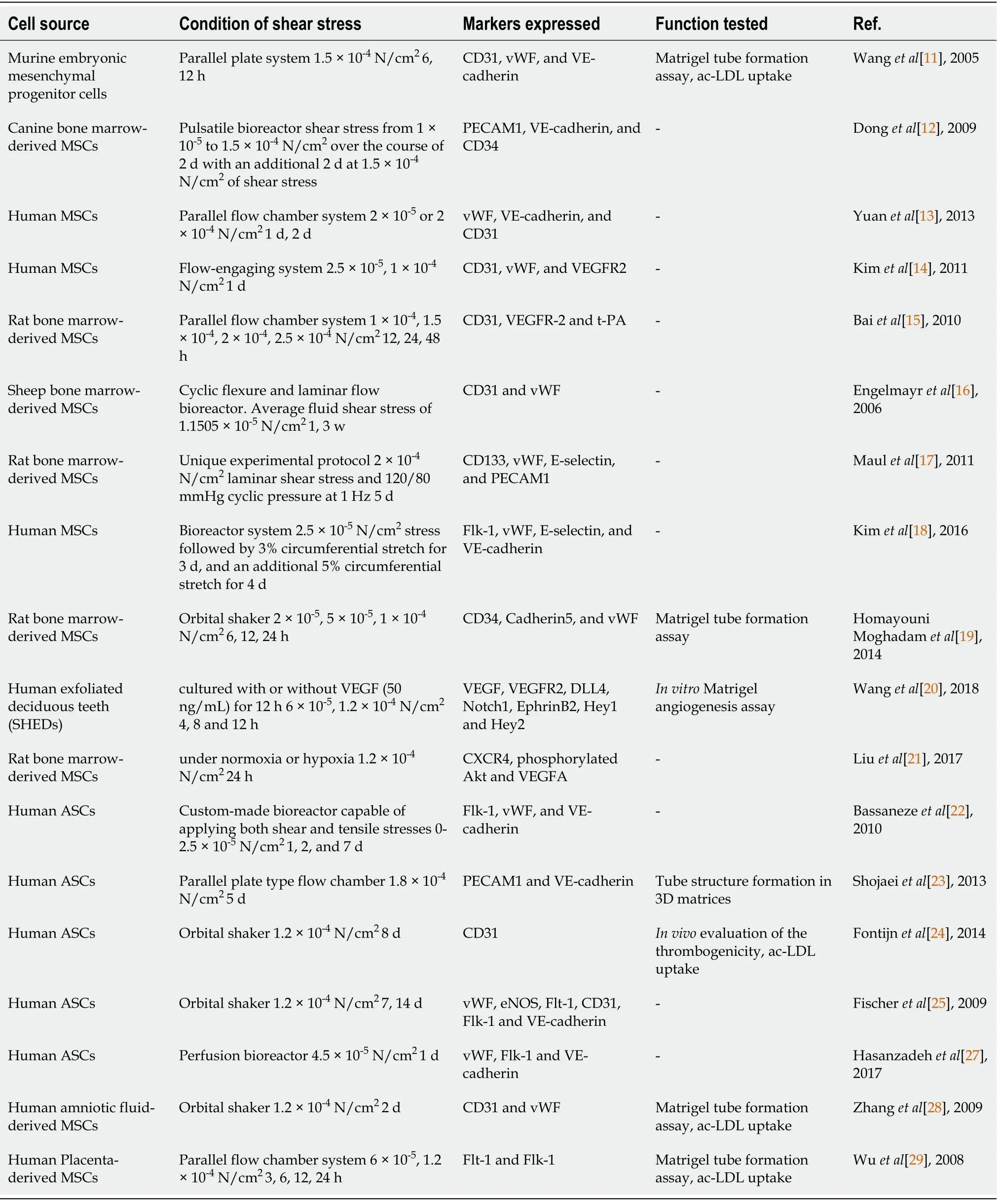
Table 1 Studies on shear stress regulating mesenchymal stem cell to endothelial differentiation
It appears that shear stress promotes endothelial differentiation while downregulating smooth-muscle-cells-oriented differentiation. An early study treated murine embryonic mesenchymal progenitor cells with 1.5 × 10-4N/cm2of shear stress using a parallel plate system. The system consisted of an active pump (for steady flow), a gravity head (for steady flow), and parallel-plate flow chambers. In flow chambers,fluid flowed through the chambers through a conduit, creating a constant hydrostatic pressure that forced culture medium to pass through the cells cultured on the lower plate, generating a steady shear stress. They found upregulated EC markers including CD31, von Willebrand factor (vWF), and vascular endothelial-cadherin (VE-cadherin,CD144), enhanced acetylated-low density lipoprotein LDL (ac-LDL) uptake, and increased tubule formation on Matrigel with 12 h of shear stress treatment. The study also showed mRNA expression of vascular smooth muscle cell markers, including PDGF receptor and PDGF, was downregulated relative to the static control[11]. Donget al[12] seeded canine bone-marrow-derived MSCs onto poly-ε-caprolactone and lactic acid scaffolds and applied them to shear stress from 1 × 10-4to 1.5 × 10-4N/cm2over the course of 2 d with an additional 2 d at 1.5 × 10-4N/cm2of shear stress provided by a pulsatile bioreactor. Similar to the structure of the parallel plate system mentioned above, the bioreactor consisted of a peristaltic pump, a compliance chamber, a glass culture reservoir (for medium circulation and air exchange), and a culture chamber.MSCs seeded on the tubular scaffolds were installed horizontally in the culture chamber. Mean shear stress (τmean) can be calculated by the equation: τmean= 4 μQ/πr3 where μ is the viscosity of the culture medium, Q is the flow rate of the bioreactor, and r is the radius of the scaffolds. They observed a significant increase in the expression of EC markers platelet/EC adhesion molecule 1 (PECAM1, CD31), VE-cadherin, and CD34, and a significant decline in the protein levels of α-smooth muscle actin (α-SMA)and calponin compared to static controls.
Physiological shear stress levels are advantageous for driving MSCs into an endothelial phenotype. In the previous two independent experiments, the magnitude of shear stress applied to stem cells was 1.5 × 10-4N/cm2. There were also other studies using higher or lower shear stresses. Yuanet al[13] individually loaded human MSCs with a steady laminar shear stress of 2 × 10-5N/cm2or 2 × 10-4N/cm2for 2 d, and then statically cultured for 5 d. They found that shear stress (2 × 10-4N/cm2) can induce cells to express vWF, VE-cadherin and CD31. Kimet al[14] reported that human MSCs exposed to a shear stress of 2.5 × 10-5or 1 × 10-4N/cm2for 1 d expressed CD31, vWF,and VEGF receptor 2 [VEGFR2, fetal liver kinase-1 (Flk-1)]. At low shear stress, CD31 was significantly expressed whereas vWF and VEGFR2 expression was only slightly higher than that, under 1 × 10-4N/cm2. In our previous work[15], we exposed rat bone-marrow-derived MSCs to a wide range of shear forces (from 1 × 10-4to 2.5 × 10-4N/cm2) and shear force durations (12-48 h). MSCs demonstrated a significant increase in expression of CD31, VEGFR-2, and tissue-type plasminogen activator (t-PA) at shear stress levels that were ≤ 1.5 × 10-4N/cm2, while higher and/or prolonged magnitude shear stress resulted in rapid decrease in EC oriented differentiation of MSCs.
Coupled mechanical stimuli, relevant to the vasculature, can differentiate MSCs toward ECs. Engelmayret al[16] reported that cyclic flexure and laminar flow (average fluid shear stress of 1.1505 × 10-5N/cm2) induced sheep bone-marrow-derived MSCs to express the endothelial-associated markers CD31 and vWF. Maulet al[17] tried 2 ×10-4N/cm2laminar shear stress and 120/80 mmHg cyclic pressure at 1 Hz and found that the rat bone-marrow-derived MSCs under this shear stress increased expression levels of endothelial-specific genes including prominin 1 (CD133), vWF, E-selectin, and PECAM1. Kimet al[18] demonstrated that 2.5 × 10-5N/cm2shear stress followed by 3%circumferential stretch for 3 d, and an additional 5% circumferential stretch for 4 d upregulated expression of several EC markers such as Flk-1, vWF, E-selectin, and VEcadherin.
The combination of biochemical and mechanical stimuli promotes MSC differentiation toward ECs. Homayouni Moghadamet al[19] used fluid shear stress as a mechanical inducer and platelet lysate and estradiol as chemical inducers. Their findings indicated that 1 × 10-4N/cm2fluid shear stress in combination with 5%platelet lysate directed MSCs to differentiate toward CD34+cells, indicating the initiation of endothelial differentiation of MSCs. In addition, when stem cells fromhuman exfoliated deciduous teeth (SHED) cultured with or without VEGF (50 ng/mL)for 12 h after shear stress (6 × 10-5, 1.2 × 10-4N/cm2for 4, 8 and 12 h), mRNA expression of angiogenic markers VEGF, VEGFR2, and CD31 was increased significantly[20]. Also, our group’s previous study showed that the combined stimulation of shear stress and VEGF resulted in more EC-oriented differentiation of
MSCs in comparison to any individual stimulation[15]. Laminar shear stress (1.2 × 10-4N/cm2for 24 h) facilitated rat bone-marrow-derived MSCs cultured in the endothelial growth medium (EGM) for endothelial maturation under both normoxic and hypoxic conditions[21].
Furthermore, MSCs derived from adipose tissue have been shown to differentiate into ECs under shear stress. Adipose tissue is a readily available source of multipotent adult stem cells for use in tissue engineering and regenerative medicine. Bassanezeet al[22] demonstrated that even though laminar shear stress (1 × 10-4N/cm2up to 96 h),produced by a cone plate system, failed to induce EC markers (CD31, vWF and Flk-1)in human adipose tissue-derived stem cells (ASCs), it stimulated nitric oxide (NO)-dependent VEGF production. Another study found that 0-2.5 × 10-5N/cm2cyclic shear stress increased expression levels of Flk-1, vWF, and VE-cadherin[23]. Fontijnet al[24]found that SOX18 transduced human ASCs, reorganized under conditions of shear stress 1.8 × 10-4N/cm2for 5 d in EGM2 plus bFGF displayed VEGF-induced chemotaxis and formed tubular structures in 3D matrices in an matrix-metalloproteinase(MMP)-7-dependent manner, suggesting that shear stress can activate differentiation of ASCs into ECs.
Synergy between biochemical factors and shear stress has been shown. Fischeret al[25] exposed human ASCs to EC growth supplement medium (for up to 3 wk) and physiological shear force (1.2 × 10-4N/cm2for up to 8 d). The combination synergistically promoted expression of CD31 and ac-LDL uptake. Human ASCs subjected to shear stress and VEGF expressed specific endothelial markers, including vWF,endothelial NO synthase (eNOS), fms-like tyrosine kinase-1 (Flt-1), CD31, Flk-1 and VE-cadherin[26,27]. In addition to bone-marrow-derived MSCs and adipose-derived MSCs, MSCs derived from other sources, such as amniotic fluid[28] and human placenta[29], can differentiate into ECs under shear stress[6].
Taken together, these studies suggest that different types of MSCs are able to differentiate into vascular ECs when they are stimulated with physiological shear stress.Mechanical and biochemical influences synergize in order to increase the expression of EC markers and EC functionality.
EPCs
Circulating EPCs are adult stem cells that play a central role in endothelial repair and blood vessel formation[30]. Hemodynamic conditions can influence localization and cell lineage differentiation of adult vascular progenitor cells[31]. EPCs are mobilized from bone marrow into peripheral blood, attach to existing ECs, and then migrate across the endothelium into tissues where they proliferate, differentiate, and form new blood vessels[32].
Shear stress has been shown to promote EPC differentiation into ECs[33].Yamamotoet al[34] reported that laminar shear stress (1 × 10-6N/cm2to 2.5 × 10-5N/cm2) accelerated cell proliferation and expression of Flk-1, Flt-1 and VE-cadherin,and upregulated capillary-like tube formation of human peripheral-bloodmononuclear-cell-derived EPCs[34]. Yeet al[35] exposed human EPCs separated from cord blood to shear stress of 5 × 10-5N/cm2by using a parallel-plate coculture flow chamber. They found that shear stress increased the expression of endothelial markers CD31 and vWF[35]. Moreover, exposure of human CD34+hematopoietic progenitor cells to shear stress increased expression of VEGF-R2, eNOS, and a VEGFR2 promoterdriven reporter gene[36]. To identify the effects of shear stress on EPC differentiation,EPCs were exposed to 5 × 10-4, 1.5 × 10-4and 2.5 × 10-4N/cm2laminar shear stress for up to 24 h or 1.5 × 10-4N/cm2laminar shear stress for 5, 10 and 20 h, which resulted in upregulation of bothin vitroendothelial differentiation andin vivoreendothelialization capacity of human EPCs in a nude mouse model in magnitude-dependent and timedependent manners[37]. Cuiet al[38] treated EPCs with different levels of shear stress(2 × 10-5, 6 × 10-5, 1.2 × 10-4and 2 × 10-4N/cm2). The results showed a dose-dependent increase in the shear-stress-induced gene expression of CD31 and vWF in EPCs above 2 × 10-5N/cm2. Chenget al[39] exposed human umbilical-cord-blood-derived EPCs to laminar shear stress of 1.5 × 10-4N/cm2using a parallel plate flow chamber system.Shear stress enhanced EPC differentiation toward ECs and inhibited smooth muscle cells differentiation[39].In vitroandin vivoassays revealed that shear stress upregulated expression levels of vWF and CD31 in EPCs, with sub-sequently increasedin vivoreendothelialization after arterial injury[40].
Further studies investigated whether EPCs differentiate into arterial or venous ECs in response to shear stress[3]. Obiet al[41] have demonstrated that controlled levels of shear stress in a flow-loading device increased the expression levels of the arterial EC markers ephrinB2, Notch1/3, Hey1/2, and activin receptor-like kinase 1, but decreased expression of the venous markers EphB4 and neuropilin 2. Exposure of human umbilical cord blood derived EPCs to shear stress of 1 × 10-6-5 × 10-5N/cm2increased the surface protein expression rate of the endothelial markers including VEGFR1 (Flt-1), VEGFR2, VE-cadherin, tyrosine kinase with immunoglobulin and EGF homology domain-2 (Tie2), vascular cell adhesion molecule 1, integrin αv/β3, Eselectin, eNOS, MMP-9 and VEGF. Likewise, in the study of Suzukiet al[42], shear stress (1.5 × 10-4N/cm2) augmented human peripheral-blood-mononuclear-cellderived EPCs to express of CD31 and vWF as well as ephrinB2, which is a marker for arterial ECs[42].
Shear stress not only has a clear effect on mature ECs, but also significantly promotes the homing of EPCs to the site of endothelial injury, encourages EPC differentiation into arterial ECs, and induces EPCs to resist thrombosis and anti-atherosclerosis (Table 2).
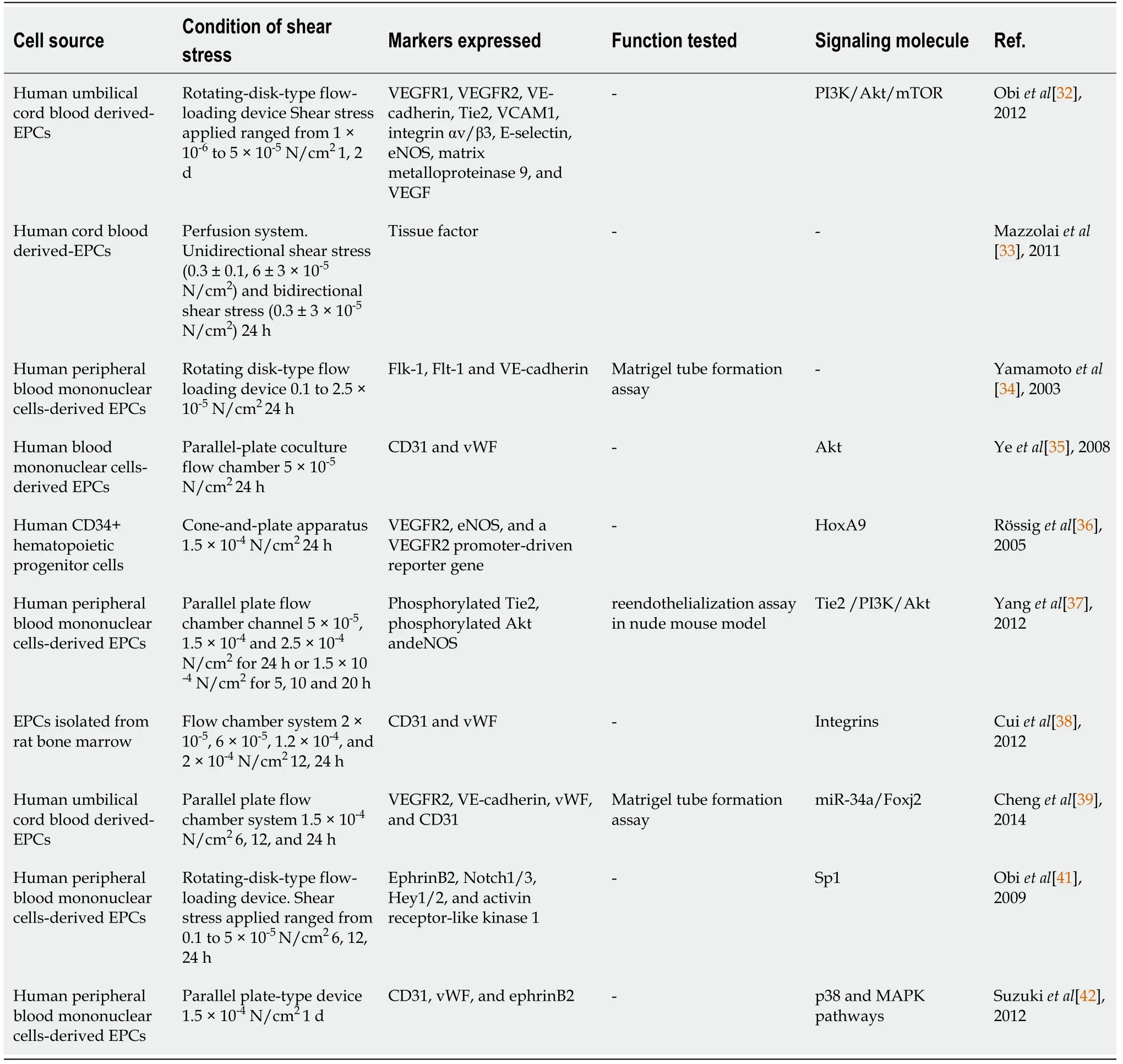
Table 2 Studies on shear stress regulating endothelial progenitor cell to endothelial differentiation
ESCs
In addition to MSCs and EPCs (adult progenitor/stem cells), ESCs have been shown to differentiate into ECs under shear stress. Shear stress is involved in endothelial differentiation during embryonic development and is key to the maintenance of a healthy endothelium[2]. Several groups have provided strong evidence that pulsatile/non pulsatile shear stress promotes phenotypic differentiation of ESCs, ESC-derived stem cell antigen-1-positive (Sca-1+) cells or ESC-derived Flk-1+into vascular ECs[43].
An early study treated mouse ESCs with a laminar shear stress of 1 × 10-4N/cm2using a cone-plate apparatus that consisted of a rotating cone placed in a tissue culture dish filled with fluid[44]. In the cone-plate system, fluid moved along the azimuth angle with the rotation of the cone and along the radial direction due to centrifugal force. Therefore, shear stress was produced at the plate. They found strongly activated transcription from the VEGFR2 promoter and early induction of endothelial markers,including VEGFR2 and PECAM1. Other cardiovascular markers, such as SMA, smooth muscle protein 22-α, myocyte enhancer factor-2C, and α-sarcomeric actin also appeared after exposure to laminar shear stress. When plated on Matrigel-coated plates, they formed tubular-like structures. Ahsan and Nerem[45] also loaded mouse ESCs with a steady laminar flow of 1.5 × 10-4N/cm2using a parallel plate flow chamber for 2 d. ESCs exposed to shear stress expressed increased levels of endothelial marker proteins (Flk-1, VE-cadherin, and PECAM1) and formed chord-like structures in anin vitroMatrigel assay. A similar result was reported on mouse ESCs by Nsiahet al[46]. The application of 2 d of fluid shear stress at 5 × 10-5N/cm2during early differentiation of mouse ESCs promoted expression of endothelial marker genes Flk1, VEcadherin and PECAM1, compared with statically cultured ESCs. To systematically investigate the effects of several mechanical parameters, Wolfe and Ahsan[47] applied laminar shear stress of 1.5 × 10-5, 5 × 10-5, and 1.5 × 10-4N/cm2to ESCs using a parallel plate bioreactor system. They found that all of them can increase the number of Flk1+,endothelial PECAM1+, and hematopoietic CD41+cells[47].
Flk-1 and/or Sca-1 are progenitor markers, usually used to isolate vascular progenitors from stem cell populations undergoing spontaneous differentiation[36].Zenget al[48] demonstrated that both mouse ESCs and Sca-1+ cells exposed to laminar shear stress (1.2 × 10-4N/cm2) increased expression of PECAM1, CD133, VE-cadherin,VEGFR1, VEGFR2, and eNOS, and formed tube-like structures on Matrigel[48].Yamamotoet al[49] exposed mouse ESC-derived Flk-1+ cells to laminar shear stress ranging from 1.5 × 10-5to 1 × 10-4N/cm2with a parallel plate-type device. Flk1+cells under the laminar flow significantly increased expression of endothelial markers (Flk1,Flt1, VE-cadherin, and PECAM1) and accumulation of cells in S and G2/M cell cycle phases, formed tubelike structures in collagen gel, and developed an extensive tubular network significantly faster than the static controls did. Biomechanical forces might act to promote hematopoiesis. Also, when murine ESC–derived Flk1+cells were applied with 1 × 10-4N/cm2shear stress, mRNA expression of the arterial EC marker ephrinB2 increased, whereas the levels of the venous EC marker EphB4 decreased in a dosedependent manner[50]. Fluid shear stress (5 × 10-5N/cm2) generated by a dynamic flow system increased expression of CD31 (PECAM1), Runt-related transcription factor 1, Myb, and Krüppel-like factor 2 (KLF2) in mouse ESC-derived CD41+c-Kit+hematopoietic progenitor cells. Moreover, shear stress increased hematopoietic colonyforming potential and expression of hematopoietic markers in the para-aortic splanchnopleura/aorta–gonads–mesonephros of mouse embryos[51].
Huanget al[52] demonstrated that not only steady but also pulsatile flow can promote an EC fate in stem cells[52]. They exposed a cell mixture containing ESCderived Flk1+cells in a compliant microporous polyurethane tube to simulate a pulsatile wall shear stress from 9.8 × 10-6to 2.2 × 10-5N/cm2and a circumferential strain stress 0.46-0.96 N/cm2for 2 d. The inner layer of the cells displayed endothelial-like appearance, and the deeper layer of the cells stained positive for smooth muscle markers.
iPSCs have broad differentiation characteristics similar to ESCs, and avoid the immune rejection and ethical issues of ESCs, so they have quickly become a research hotspot in the field of stem cells. Our group’s previous study showed that exposure of mouse iPSCs to shear stress (5 × 10-5, 1 × 10-4, and 1.5 × 10-4N/cm2) with 50 ng/mL VEGF and 10 ng/mL FGF increased expression of the general EC markers and arterial markers, during which the stress amplitude of 1 × 10-4N/cm2could be regarded as a proper promoter, whereas the venous and lymphatic markers had little or no expression. Shear stress caused cells to align parallel to the direction of the flow,induced cells forming functional tubes, and increased secretion of NO. In addition,Notch1 was significantly upregulated, and the Notch ligand Delta-like 4 was activated in response to shear stress, while inhibition of Notch signaling by dual antiplatelet therapy (DAPT) abolished the shear stress-induced arterial epithelial differentiation[53].
Most of the previous experimental systems involved the use of bioreactors or viscometers, like parallel plate flow and conical flow reactors[3]. To overcomes the limitations of macroperfusion systems in shear application throughput and precision,Toh and Voldman[54] used a multiplex microfluidic array that applied shear stresses varying by > 1000 times (1.6 × 10-7-1.6 × 10-4N/cm2) to mouse ESCs. In the microfluidic device, they used a fluid column or a syringe pump to drive blood or culture medium over cells cultured in capillary tubes or in customized poly(dimethylsiloxane) chips,producing shear stress. Shear stress specifically upregulated the epiblast marker Fgf5.Epiblast-state transition involved heparan sulfate proteoglycans, which have also been shown to transduce shear stress in ECs[3]. Leeet al[55] developed an integrated microfluidic culture device consisting of an air control channel and a fluidic control channel with 4 × 4 microcolumn arrays. They demonstrated that ESCs cultured for 6 d in the integrated microfluidic culture device were more differentiated into PECAM+ECs.
Overall, ESCs and iPSCs are potential sources for cell-based tissue engineering and regenerative medicine applications (Table 3). Increasing evidence suggests that appropriate shear stress, especially 1 × 10-4-1.5 × 10-4N/cm2, may be a useful tool for promoting ESC differentiation into ECs. The adoption of microfluidic technologies will help to circumvent current technical limitations and provide quantitative shear application benchmarks for future scalable stem cell culture systems.
SIGNALING PATHWAYS OF SHEAR-STRESS-INDUCED EC DIFFERENTIATION
Mechanotransduction is the process by which mechanical stimuli are converted into biochemical signals inside the cell, enabling the cell to adapt to its environment,including three stages: Mechanotransmission, mechanosensing and mechanoresponse[56]. Mechanotransmission, as the name suggests, is the propagation of mechanical forces along structures, such as the cytoskeleton. Mechanosensing refers to the process in which a force acts on a mechanically sensitive macromolecule after mechanical transmission, changing its conformation, and thus, affecting its function. Although the biological effects of different mechanical forces in different systems are specific, the underlying physical response is similar: the force promotes a change in the conformation of the applied force. The process that the perceived mechanical signal produces a series of biological effects through the complex cellular signal and transcriptional network is called mechanoresponse, which is not as fast and direct as the first two processes, and in many cases, these reactions alter the mechanically sensitive structures that cause the reactions.
Over the past few decades, it has been widely reported that shear stress plays a critical role in endothelial differentiation of types of stem cells to remodel blood vessels and repair vascular damages[57]. Evidence of several potential signaling pathways involved in shear-stress-induced endothelial specification of different stem cells has been identified; however, the connections and interactions of these mechanosensitive molecules are still unclear. In detail, the mechanism by which shear stress promotes the mobilization of circulating phenotype PSCs from the peripheral blood or bone marrow to the injured site, the differentiation of the tissue adhered cells into mature ECs, and the restoration of vascular structure and functions is worth investigating. In addition, EPCs are divided into two types according to differences in protein expression and function. The cells involved in the early endothelial differentiation, called early EPCs, lack the capacity to differentiate into functional ECs, but activate resident ECs through paracrine factors[58], while the cells associated with the latter process, named late EPCs, are able to incorporate with vascular endothelium and generate new capillariesin vivo[59]. This review shows how the mechanosensors and the downstream signals convert mechanical stimuli into biochemical signals inside the stem cells during the early (Figure 1) and late (Figure 2) stages of endothelial differentiation mediated by shear stress.
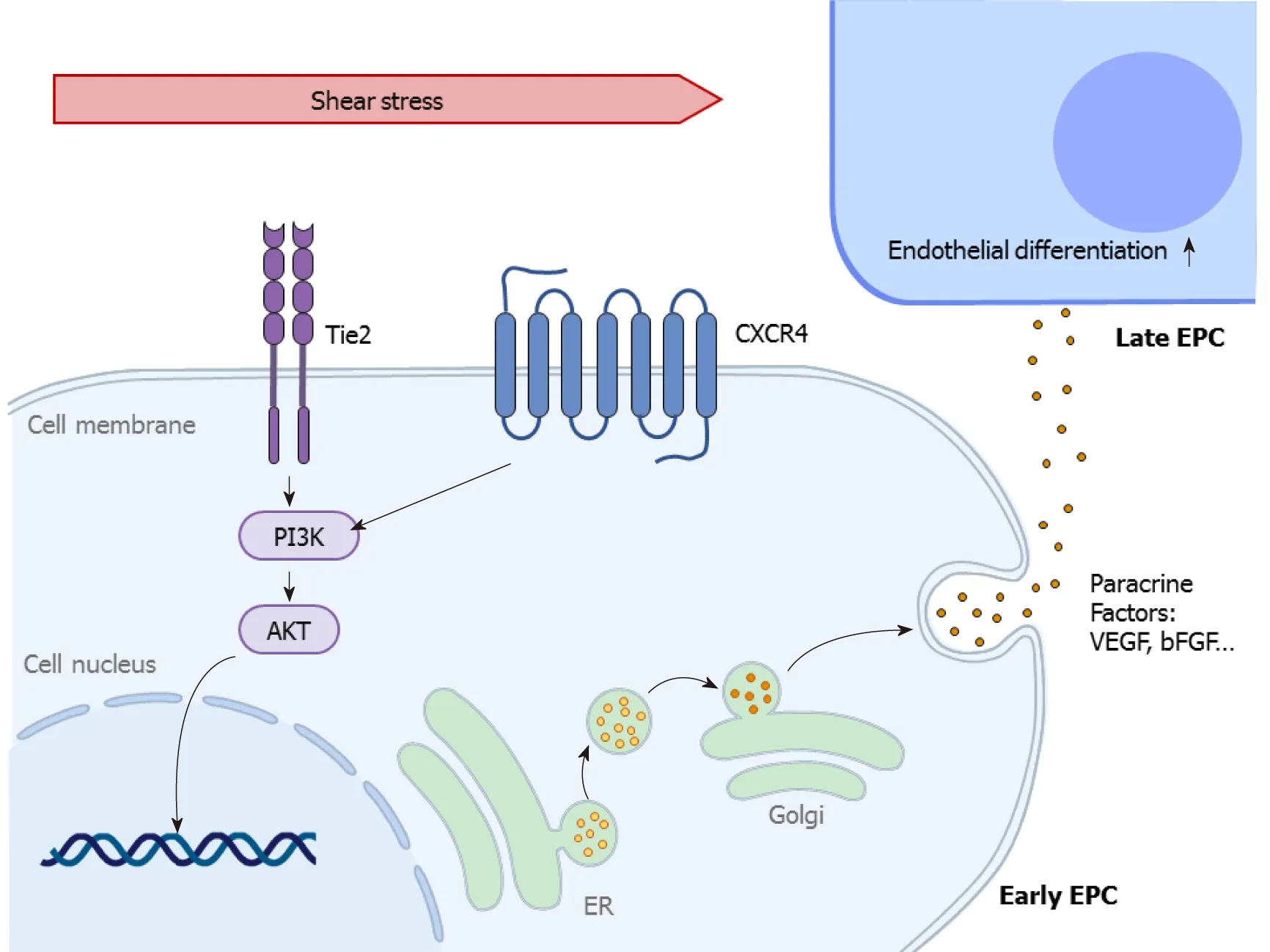
Figure 1 Shear-stress-induced signaling pathways in early endothelial differentiation. This figure describes how the mechanosensitive molecules tyrosine kinase with immunoglobulin and epidermal growth factor homology domain-2and C-X-C chemokine receptor type 4 sense the shear stress, induce phosphoinositide 3-kinase-protein kinase B signaling activation, and further influence the gene expression of various paracrine factors ultimately regulating vessel maintenance and reformation. Akt: Protein kinase B; bFGF: Basic fibroblast growth factor; CXCR4: C-X-C chemokine receptor type 4; EPC: Endothelial progenitor cells PI3K: Phosphoinositide 3-kinase; Tie2: Tyrosine kinase with immunoglobulin and epidermal growth factor homology domain-2;VEGF: Vascular endothelial growth factor.
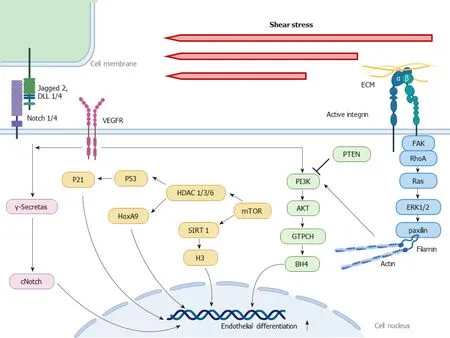
Figure 2 Shear stress-induced signaling pathways in late endothelial differentiation. This figure describes how the mechanosensitive molecules such as Notch 1/4, vascular endothelial growth factor receptor, integrins respond to the shear stress, accompanied by cytoskeleton reorganization via RhoA, Ras,ERK 1/2 and paxillin, and activate phosphoinositide 3-kinase-protein kinase B signaling, regulating various pathways including guanosine triphosphate cyclohydrolase-tetrahydrobiopterin, histone deacetylase (HDAC) 1/3/6-P53-P21, HDAC 1/3/6-HoxA9 and sirtuin 1-H3. These signaling cascades influence the gene expression of stem cells, ultimately regulating vessel maintenance and reformation. Akt: Protein kinase B; GTPCH: Guanosine triphosphate cyclohydrolase; BH4:Tetrahydrobiopterin; ECM: Extracellular matrix; HDAC: Histone deacetylase; KLF2: Krüppel-like factor 2; PI3K: Phosphoinositide 3-kinase; PTEN: Tumor suppressor phosphatase and tensin homolog; VEGFR: Vascular endothelial growth factor receptor.
Notch signaling pathway
The Notch signaling pathway, which contains four Notch receptors and five Notch ligands (Jagged 1 and 2, and Delta-like 1, 3 and 4) in mice and humans[60], plays a fundamental role in promoting arterial-venous differentiation throughout embryonic vascular development[61-63]. It has been universally demonstrated that Notch affects embryonic vascular development[64-66] and regulation of arterial-venous differen-tiation[67]. Specifically, Notch 1, Notch 4, Jagged 2, and Delta-like 1 and 4 have an important role in arterial specification[68-74]. Notch 1 has been identified as a mechanosensor that is responsible for mediating flow-induced arterial homeostasis[75,76]. However, Kimet al[77]found thatin vitroendothelial colony-forming cells(ECFCs), one kind of EPCs, derived from human cord blood, with preconditioning of Notch activation with an immobilized chimeric Notch ligand (Delta-like1ext-IgG), failed to promote vasculogenesisin vivo. Coimplantation of ECFCs and stromal cells expressing the Notch ligand has a positive effect on vessel density and areain vivo[77].Moreover, using a c-secretase inhibitor (DAPT) to block the Notch pathway resulted in progenitor quiescence and reduction of ECFCs colony-formation potential, reflecting loss of progenitor capacity[78]. Therefore, the function of the Notch signaling pathway in shear-stress-induced EC differentiation remains to be studied.
Recently, it has been shown that VEGFR-Notch-EphrinB2 signaling is involved in shear stress regulation of PSC differentiation into arterial ECs[79-83]. VEGF binding to its receptors, VEGFR1 (also known as Flt-1) and VEGFR2 (also known as KDR and Flk-1), increases the endothelial differentiation of stem cells[84-87], including MSCs[88,89], EPCs[90], ESCs[91] and iPSCs[92]. VEGFR has been found to be sensitive to sheer stress in a ligand-independent manner[93], which activates the Notch pathway and determines EC fate[94]. EphrinB2 exists mainly on arterial ECs, and EphB4-preferring venous ECs are known to play a key role in embryonic vascular development[95-97],which could be regulated by VEGFR-Notch signaling[98]. According to previous studies, Notch signaling has been suggested to downregulate expression of EphB4 in ESCs, human umbilical vein ECs, and adult ECs, which is reported to be HERPdependent[99]. To be more specific, activation of the Delta-like 4/Notch pathway by VEGF selectively increases expression of EphrinB2, and thus, promotes angiogenesis[100]. Shear stress induces EC differentiationviaVEGFR-Notch-EphrinB2 signaling.For instance, murine ESC–derived VEGFR2+cells in a shear-stress-loading device with a shear stress of 1 × 10-4N/cm2for 24 h upregulated EphrinB2 expression, which was found to be blocked by DAPT (2.5 μmol/L) and L685 458 (0.1 μmol/L), inhibitors of γsecretase, which was required in the sequential proteolytic events in Notch signaling pathway activation. The VEGFR kinase inhibitor SU1498 (10 μmol/L) suppresses shear-stress-induced cleaved Notch, which is essential in transactivation of various gene promoters during embryonic vascular development. Our previous study demonstrated that mouse iPSCs cultured with 50 ng/mL VEGF and 10 ng/mL FGF induced expression of Notch 1 and Delta-like 4 in response to shear stress (5 × 10-5, 1 ×10-4, and 1.5 × 10-4N/cm2) for 4 h. In turn, this caused upregulation of arterial markers EphrinB2 and neuropilin-1, which was blocked by DAPT (50 μmol/L)[53]. However,shear stress cannot continuously maintain high mRNA expression of EphrinB2 and EphB4 in murine tumor models, which might be related to a negative-feedback loop between VEGF-Notch signaling[50]. In addition, the mRNA expression of VEGF and VEGFR2 was upregulated in SHED cultured with or without VEGF (50 ng/mL) for 12 h after shear stress values of 4 × 10-5and 1.6 × 10-4N/cm2for 2 h. There were no significant changes in the expression of EphrinB2 and EphB4[20]. These results suggest that the EC differentiation potential of post-natal MSCs mediated by VEGFR-Notch-Ephrin B2 signaling is limited.
As mentioned above, Notch is the junctional adhesion receptor activated by binding to ligands expressed on adjacent cells. This adhesion appears in shear-stress-induced late EC differentiation of PSCs with high plasticity. Therefore, the Notch signaling pathway regulates cells to home to ischemic lesions and generate mature endothelial progeny under shear stress. More importantly, Ephrin B2, a downstream molecule of Notch, plays a key role in arteriovenous differentiation, and is not persistently expressed in post-natal MSCs under shear stress conditions. More studies are required to clarify the mechanisms.
Phosphoinositide 3-kinase-Akt signaling
Shear stress promotes early endothelial differentiation of stem cells mobilized from bone marrow, such as MSCs and early EPCs,viaphosphatidylinositol 3 kinase (PI3K)-Akt signaling. Tie2 and its ligand, angiopoietin 2 (Ang2), contribute to the increase in proliferation, migration, and survival of CD34+stem cells derived from human umbilical cord blood, and enhance neovascularization to restore injured vasculature[101-104]. Shear stress (2 × 10-4N/cm2for 5 min) activated PI3K and Akt signalsviaTie/Ang2 signaling in human ECs[105], which might also occur in EC differentiation of stem cells. Thus, Yanget al[37] focused on the Tie2/PI3k/Akt signaling pathway in early EPCs exposed to laminar shear stress and demonstrated that shear stress activated Tie2 and phosphorylated Akt (pAkt) in a dose-dependent manner, thus increasingin vitroendothelial differentiation andin vivore-endothelialization capacity of human EPCs in nude mouse model, which was markedly inhibited after Tie2 knockdown or PI3K inhibition[37]. More importantly, they found that eNOS, one of the recognized vascular repair molecules of circulating EPCs[106-108], could be directly controlled by shear stress through the Tie2/PI3K/Akt pathway on account that shRNA knockdown of theTie2gene or pharmacological inhibition of PI3K could significantly inhibit shear-stress-induced phosphorylation of Akt and eNOS in EPCs.Recently, it was demonstrated that an increase of C-X-C chemokine receptor type 4(CXCR4) and pAkt protein expression in MSCs and eEPCs, which could be promoted by shear stress (1.2 × 10-4N/cm2) for 24 h even under hypoxic conditions, facilitated these cells toward mature ECs, reflected by an increase in endothelial markers PECAM-1 and VEGFA, indicating that the CXCR4-PI3K-Akt pathway was important to regulate the early endothelial differentiation in a hypoxic microenvironment[21]. All these findings demonstrate that Tie2-PI3K-Akt signaling or CXCR4-PI3K-Akt signaling was, at least in part, concerned with the shear-stress-mediated function of MSCs and eEPCs bothin vitroand invivo.
PI3K-Akt signaling was also involved in the late stage of endothelial differentiation induced by shear stress through increasing adhesion, migration, proliferation, and finally, tube formation. The tumor suppressor phosphatase and tensin homolog(PTEN) is activated during angiogenesis of ECs and EPCs by reducing expression of the PI3K-Akt pathway, and in turn downregulates angiogenesis and vasculogenesis[109-111]. Recently, Wuet al[112] demonstrated that shear stress (5 × 10-5, 1.5 × 10-4, and 2.5 × 10-4N/cm2for 5, 10, and 15 h) enhanced the functions of EPCsin vitroandin vivo, which was associated with downregulation of PTEN expression, phosphorylation of Akt and activation of the guanosine triphosphate cyclohydrolase (GTPCH)-tetrahydrobiopterin (BH4) pathway, which is critical to the synthesis of NO. When Akt phosphorylation specific inhibitor, LY was added, shear-stress-induced activation of the GTPCH-BH4 pathway and tube formation of late EPCs were suppressed,suggesting that the PTEN-Akt-GTPC-BH4 pathway contributed to shear-stressenhanced functions of late EPCs during angiogenesis[112]. Moreover, VEGFR2 was another accepted upstream signaling molecule of PI3K-Akt signaling, which could further stably activate histone deacetylases (HDACs). In late EPCs derived from human umbilical cord blood, culture with shear stress (2.5 × 10-6, 5 × 10-6, 1 × 10-5, 2.5 ×10-5N/cm2) for 24 and 48 h promoted differentiation of the cells into mature ECs, with expression of the endothelial marker VEGF-R2 increasing in a ligand-independent manner. After treatment with PI3K inhibitor and mTOR inhibitor in EPCs exposed to a shear stress of 2.5 × 10-5N/cm2for 48 h, expression of endothelial marker proteins VEGFR1, VEGFR2, VE-cadherin, and Tie2 was markedly decreased, indicating that PI3K-Akt-mTOR signaling is the most potent transduction pathway of endothelial differentiation in response to shear stress[113]. Rössiget al[36] found that inhibition of HDACs prevented endothelial differentiation from adult progenitor cells.Furthermore, overexpression of HoxA9, a homeobox transcription factor, partially rescued the negative influence of HDAC inhibitors and mediated EC maturation induced by shear stress (1.5 × 10-4N/cm2for 24 h). These results indicated that inhibition of HDACs decreased expression of HoxA9 and then inhibited the endothelial lineage commitment of different progenitor cell sources induced by shear stress[36]. Furthermore, 1.2 × 10-4N/cm2laminar shear stress for 12 and 24 h enhanced the differentiation of ESC-derived progenitor cells into ECs by stabilizing and activating HDAC3 through the Flk-1-PI3K-Akt pathway, which in turn deacetylated p53, leading to p21 activation, thus promoting EC differentiationin vitroandin vivo[48]. As for the sirtuin (SIRT) family, class III HDACs, shear stress of 1.5 × 10-4N/cm2for 2, 6, 12, and 24 h upregulated SIRT1 by activation of the PI3k-Akt pathway and resulted in deacetylating histone H3, which induced EPC differentiation toward ECsin vitro[114]. HDAC6 is important for restoring primary cilia after 24 h of 2 × 10-4N/cm2shear stress applied to iPSC-derived ECs[115]. However, the HDAC inhibitor trichostatin A increases expression of EC markers, such as VE-cadherin, vWF and Flk1 in bone marrow progenitor cells and stimulates vascular network formationin vivo,indicating that the HDACs have an opposite effect and suppress endothelial differentiation[116]. Therefore, the role of HDACs in differentiation into ECs and the effect of the PI3K-Akt pathway on shear-stress-induced HDACs regulation require further research.
PI3K-Akt binding to different upstream and downstream molecules plays an important role at any stage of shear-stress-induced endothelial differentiation. Early endothelial differentiation is regulated by the Tie2-PI3K-Akt-eNOS signaling pathway in early EPCs and CXCR4-PI3K-Akt pathway in MSCs, while the inhibitory effect of PTEN on PI3K-Akt signaling and VEGFR2-PI3K-Akt-mTOR pathway occurs in late endothelial differentiation of several stem cells, including ESCs, EPCs and iPSCs.HDACs are known for their effects in regulating vascular health[117-119]. However,the mechanisms by which HDACs regulate shear-stress-induced EC differentiationviathe PI3K-Akt pathway have not been elucidated.
Integrin-cytoskeleton system
Integrin[120] and cytoskeletal filaments[121] have been verified to contribute to mechanotransduction,i.e.mediating the shear-stress-induced endothelial commitment of stem cells. Integrin can be activated by shear stress and mediate cell-extracellular matrix (ECM) and cell-cell interactions, which trigger downstream signals, including RhoA, Rac (belonging to the Ras super-family of proteins), and Cdc42 activation[122,123]. Cuiet al[38] have shown that shear stress at 1.2 × 10-4N/cm2for 2, 6, 12 and 20 h upregulated the expression of integrin β1 and β3 and increased expression of EC differentiation markers in late EPCs isolated from rat bone marrow, in a timedependent manner. This process could be inhibited by anti-β1 integrin and anti-β3 integrin antibodies that blocked the binding of integrins to the ECM. Therefore,integrins β1 and β3 play a key role in regulating the shear-stress-induced late EPC differentiation[38]. During integrin β1-related signals, Ras, one of the small G proteins,was the earliest link between mechanical perception and the downstream signal transduction cascades[124]. Moreover, integrins associated with RhoA participate in the process of cytoskeletal rearrangement, and further EPC differentiation after applying shear stress[125]. Chenget al[126] have demonstrated that Ras, ERK1/2 and paxillin activated by integrin β1 are important mechanosensors involved in cytoskeletal remodeling, which subsequently upregulate expression of endothelial markers vWF and CD3 in late EPCs from rat bone marrow exposed to shear stress at 1.2 × 10-4N/cm2for 5, 30 or 60 min and promotes re-endothelialization in rats with arterial injury[126]. Additionally, to determine the order in which these signaling molecules act, pretreatment of EPCs with the anti-integrin β1 antibody (50 mg/mL),Ras-negative mutant (RasN17), ERK1/2 specific inhibitor PD98059, and the mediated silencing of the paxillin under shear stress at 1.2 × 10-4N/cm2for 1 h was applied. This suggested that cytoskeletal remodeling is associated with shear-stress-induced endothelial differentiation by activating integrins, especially integrin β1 and β3, Ras,ERK1/2 and paxillin in sequence.
Cytoskeletal rearrangement associated with integrins might mediate the translocation of numerous signaling molecules, which in turn facilitate activation of the downstream signal transduction cascades that regulate endothelial differentiation on encountering shear stress. For instance, VEGFR-2/Flk-1 might be of importance to transduce signals through the integrin-cytoskeleton system and contribute to shear stress-induced endothelial differentiation because of the upregulation of integrin expression by VEGF during angiogenesis in mature ECs[127,128]. Also, cytoskeletal rearrangement contributes to the shear-stress-induced PI3K and Akt activation mediated by integrins, which in turn facilitates endothelial differentiation. More recently, the Notch signaling-regulating role of cytoskeletal protein vimentin has been addressed during arterial remodeling accelerated by shear stress[129]. In this research,1 Pa shear stress for 24 h enhanced Jagged 1 levels and further increased Notch signal activation. When the Notch reporter cells cocultured with vimentin knock-out cells were exposed to shear stress, the regulation of Jagged 1-Notch signaling was inhibited,indicating the importance of vimentin in Jagged 1-Notch transactivation during shear stress. Chuet al[130] also pointed out that KLF2, a member of the zinc finger transcription factor family, was essential for the regulation of the integrin-cytoskeleton system in endothelial differentiation of rat late EPCs under shear stress (1.2 × 10-4N/cm2)[130]. Downregulation of KLF2 expression by siRNA resulted in inhibition of endothelial differentiation, with decreased protein levels of EC markers, CD31, and vWF. Moreover, blocking integrin β1/β3 with antiintegrin antibodies, or disrupting cytoskeletal protein F-actin with cytochalasin D, interfered with activation of KLF2.Upregulation of KLF2 expression is involved in shear-stress-induced differentiation of EPCs toward mature ECs, which may be associated with the integrin-cytoskeleton system.
The association of integrins and the cytoskeleton mediated by RhoA, Ras, ERK1/2,paxillin and focal adhesion kinase in turn happens at focal adhesions[131], indicating that the integrin-cytoskeleton system reacts to shear-stress-induced endothelial differentiation of attached tissue type EPCs. In addition, the integrin-cytoskeleton system, as a critical mechanosensor, responds to shear stress and activates signaling pathways that regulate EC differentiation, such as VEGFR-2/Flk-1, PI3K-Akt pathway, Notch signaling and KLF2.
CONCLUSION
The present review summarizes recent results on how shear stress influences stem cell differentiation into ECs. The underlying mechanisms involved in this process include integrin-mediated signaling, cytoskeletal reorganization, activation of intracellular signaling cascades, such as Notch signaling, PI3K-Akt pathway, VEGFR-2/Flk-1,KLF2, and nuclear translocation, leading to expressions of a variety of genes, which finally promotes an endothelium-oriented phenotype in stem cells.
However, many questions concerning the relationship between shear stress and the responses of stem cells remain unanswered. For example, stem cells can differentiate into ECs, muscle cells, osteocytes, adipocytes, and neural cellsin vitro, and also are stimulated by shear stretch and compressive stressin vivo. How do the stem cells sense these physical forces to go in a specific direction? Further investigation on this issue would enhance our control or manipulation of stem cell differentiation. Individual shear stress discussed here does not seem to drive stem cells toward arterial or venous ECs. It is unlikely that a single factor will be able to determine the clear demarcation between arterial and venous phenotypes. Hence, it is necessary to combine a variety of biophysical and biochemical cues to support more effective phenotypic specification.In addition, it is unknown if the blood flow under physiological or pathological conditions influences the EC-oriented differentiation of stem cells. Finally, with the advent of nanotechnology and microfluidic technologies, elucidating microfluidic dynamics would help our understanding of how shear stress affects stem cell differentiation. Taken together, the deeper understanding of how stem cells respond to mechanical forces would be able to produce a mechanical-force-mediated tissueengineered vessel and enhance the clinical translation of stem-cell-based strategy.
杂志排行

World Journal of Stem Cells的其它文章
- Epigenetic modulators for brain cancer stem cells: Implications for anticancer treatment
- Mechanisms involved in selecting and maintaining neuroblastoma cancer stem cell populations, and perspectives for therapeutic targeting
- Roles of mitochondrial unfolded protein response in mammalian stem cells
- Stem cell therapies in tendon-bone healing
- Exosomal microRNAs from mesenchymal stem/stromal cells:Biology and applications in neuroprotection
- Immunotherapy against programmed death-1/programmed death ligand 1 in hepatocellular carcinoma: Importance of molecular variations, cellular heterogeneity, and cancer stem cells